Phenotypic and Genotypic Profile of Enterobacteriaceae Isolated at a Teaching Hospital in Ghana
Bismark Donkor, Richael Odarkor Mills, Philimon Mwintige, Alberta Bedford Moses, Abigail Asmah Brown, Faustina Halm‐Lai, Oheneba Charles Kofi Hagan

TL;DR
This study analyzed antibiotic-resistant bacteria from a Ghanaian hospital, finding high resistance and diverse genetic factors that could worsen healthcare outcomes.
Contribution
The study provides a detailed genomic and phenotypic profile of Enterobacteriaceae isolates in a Ghanaian hospital setting.
Findings
Isolates showed high resistance to β-lactams and harbored multiple antibiotic resistance genes.
Common resistance genes included CTX-M-15 and NDM-1 in some Klebsiella pneumoniae isolates.
Virulence factors and plasmid replicons were detected, emphasizing the need for genomic surveillance.
Abstract
Antibiotic resistance in Enterobacteriaceae continues to rise, and its implications for healthcare delivery have intensified. We investigated the genetic basis of antimicrobial resistance (AMR), virulence genes, and associated plasmids in Enterobacteriaceae isolates from a teaching hospital in Ghana. Antimicrobial susceptibility testing was performed using archived isolates. Whole genome sequencing was performed on a subset of the isolates, that were either multidrug resistant or extended‐spectrum β‐lactamase (ESBL)‐producing. Bioinformatics analyses were performed for speciation, identification of AMR and virulence genes, and associated plasmids. The 100 Enterobacteriaceae isolates included in this study showed high phenotypic resistance to ß‐lactams and susceptibility to aminoglycosides. Nineteen of the 20 WGS isolates were genotypically identified using the housekeeping genes as…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
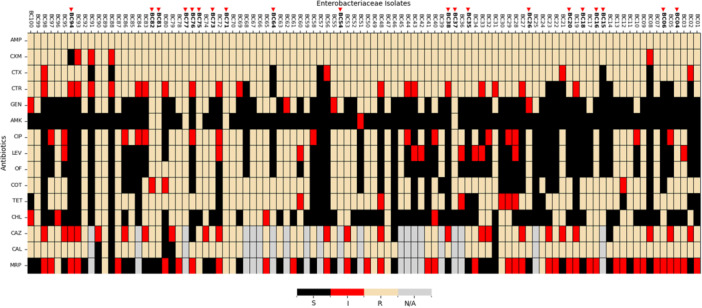 Figure 1
Figure 1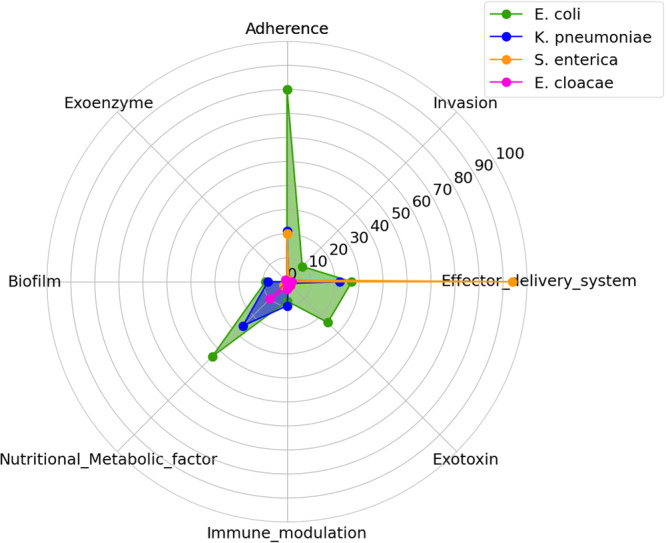 Figure 2
Figure 2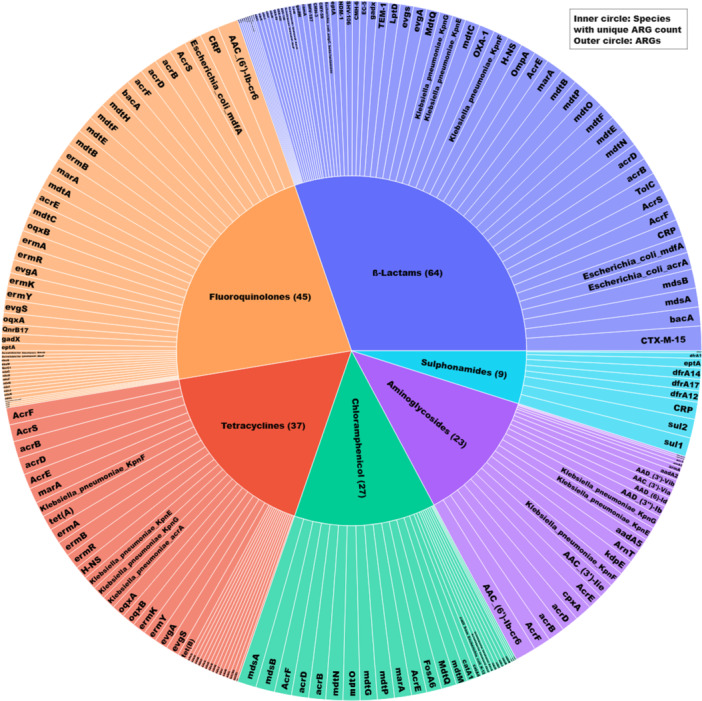 Figure 3
Figure 3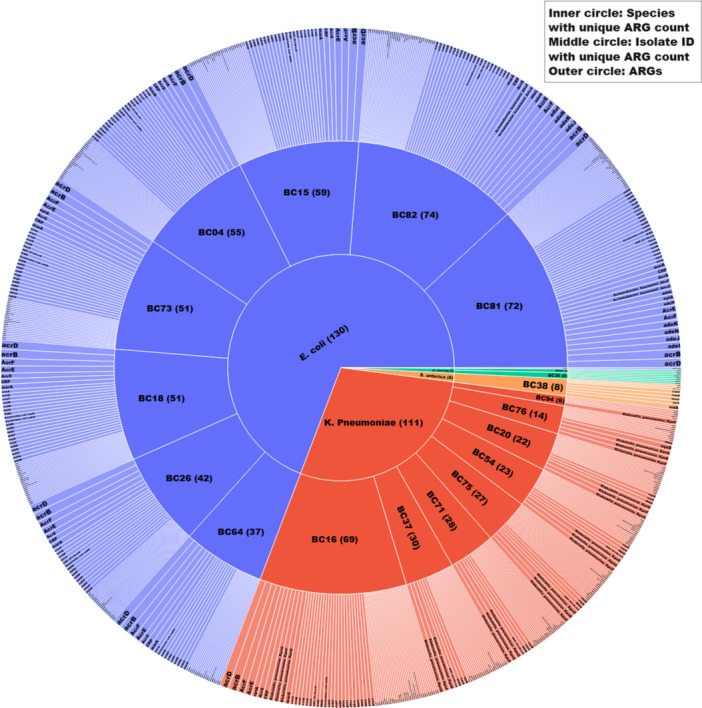 Figure 4
Figure 4| Category | Subcategory | Percentage (%) |
|---|---|---|
| Age in years | ≤ 12 years | 12.0 |
| 13–19 | 2.0 | |
| 20–39 | 34.0 | |
| 40–69 | 37.0 | |
| ≥ 70 | 15.0 | |
| Sex | Female | 70.0 |
| Male | 30.0 | |
| Department | OPD | 51.0 |
| Other Departments | 13.0 | |
| A/E | 10.0 | |
| FMW | 8.0 | |
| MMW | 7.0 | |
| ETAT | 6.0 | |
| PMW | 5.0 | |
| Specimen source | Urine | 44.0 |
| Sputum | 14.0 | |
| HSV | 12.0 | |
| Wound swab | 12.0 | |
| Other sources | 10.0 | |
| Blood | 8.0 |
| Virulence factor genes | |||
|---|---|---|---|
| Organism | Category | Subcategory | No. unique VFs |
|
| Adherence | ||
| Fimbrial adhesin | 77 | ||
| Non‐fimbrial adhesin | 2 | ||
| Invasion | 9 | ||
| Effector delivery system | |||
| Type II secretion system (T2SS) | 11 | ||
| Type V secretion system (T5SS) | 2 | ||
| Type VI secretion system (T6SS) | 14 | ||
| Exotoxin | |||
| Membrane‐acting toxin | 5 | ||
| Intracellularly active toxin | 19 | ||
| Immune modulation | |||
| Antiphagocytosis | 1 | ||
| Complement evasion/Serum resistance | 1 | ||
| Inflammatory signaling pathway | 6 | ||
| Nutritional/Metabolic factor | |||
| Metal uptake | 44 | ||
| Biofilm | |||
| Biofilm formation | 7 | ||
| Quorum sensing | 2 | ||
|
| Adherence | ||
| Fimbrial adhesin | 21 | ||
| Invasion | 1 | ||
| Effector delivery system | |||
| Type II secretion system (T2SS) | 1 | ||
| Type VII secretion system (T7SS) | 21 | ||
| Exotoxin | |||
| Intracellularly active toxin | 1 | ||
| Immune modulation | |||
| Complement evasion/Serum resistance | 5 | ||
| Inflammatory signaling pathway | 5 | ||
| Nutritional/Metabolic factor | |||
| Metal uptake | 25 | ||
| Metabolic adaptation | 1 | ||
| Biofilm | |||
| Biofilm formation | 5 | ||
| Quorum sensing | 3 | ||
|
| Adherence | ||
| Fimbrial adhesin | 19 | ||
| Non‐fimbrial adhesin | 1 | ||
| Invasion | 1 | ||
| Effector delivery system | |||
| Type III secretion system (T3SS) | 85 | ||
| Type VI secretion system (T6SS) | 9 | ||
| Nutritional/Metabolic factor | |||
| Metal uptake | 2 | ||
|
| Effector delivery system | ||
| Type VII secretion system (T7SS) | 2 | ||
| Exotoxin | |||
| Membrane‐acting toxin | 2 | ||
| Immune modulation | |||
| Antiphagocytosis | 3 | ||
| Nutritional/Metabolic factor | |||
| Metal uptake | 10 | ||
| Exoenzyme | |||
| Protease | 1 | ||
| PGs |
|
|
| Number of isolates | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BC04 | BC15 | BC18 | BC26 | BC64 | BC73 | BC81 | BC82 | BC16 | BC20 | BC37 | BC54 | BC71 | BC75 | BC76 | BC94 | BC06 | BC35 | |||
| Incompatibility (I) complex Plasmid |
|
| ||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
| Colicin plasmid |
|
| ||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
|
|
| |||||||||||||||||||
| Total PGs in isolate |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
- —The authors received no specific funding for this study.
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Antibiotic Use and Resistance · Pharmaceutical and Antibiotic Environmental Impacts
Introduction
1
The family Enterobacteriaceae is widely distributed in nature, particularly in the environment and gastrointestinal tracts of animals. They are Gram‐negative, non‐spore‐forming, facultative anaerobes, reduce nitrate to nitrite, and ferment glucose (Octavia and Lan 2014; Janda and Abbott 2021; Jenkins et al. 2017). Although the majority of these are commensals, some genera have an impact on human and animal health. These include Escherichia coli, Klebsiella spp, Citrobacter spp, Salmonella spp, and *Enterobacter spp (*Janda and Abbott 2021; Tacconelli et al. 2018; Renata Silva‐Lacerda et al. 2015).
The emergence and persistence of antimicrobial resistance (AMR) in the Enterobateriaceae to antimicrobials of various classes, especially extended‐spectrum ß‐lactams, carbapenems, and fluoroquinolones, in clinical settings have hampered global healthcare delivery, especially in lower‐ and middle‐income countries (LMICs) 2020 (Walsh et al. 2023; Lynch et al. 2021). A systematic review and meta‐analysis by Bezabih et al. found a 21.1% in the global antibiotic resistance Enterobacteriaceae (ARE) rate, particularly in isolates that are ESBL carriage from 2003 to 2018 in the healthcare settings (Bezabih et al. 2022a). Other studies have highlighted the prevalence of the CTM‐X‐15 gene, a type of ESBL gene that is dominant among pathogenic Enterobacteriaceae (Irenge et al. 2019; Asare Yeboah et al. 2024). With regard to carbapenem resistance, the pooled prevalence globally is higher than the data available in Africa. They reported NDM, IMP, and VIM types as the main drivers of carbapenem‐resistant Enterobacteriaceae (CRE), especially NDM‐1. However, data present in Ghana reveal low harborage of NDM‐1 (Bachelle et al. 2024; Ayibieke et al. 2018). Nonetheless, CRE in Ghana continues to rise. Mills et al. and Ayibieke et al. reported that carbapenem‐hydrolyzing class D β‐lactamases, mainly OXA‐type genes, were observed among Enterobacteriaceae in Ghana (Ayibieke et al. 2018; Mills et al. 2024).
Aside from antibiotic‐resistant genes (ARGs), these bacteria harbor genes of virulence factors (VFs). VFs are important for Enterobacteriaceae to successfully colonize and survive in their hosts (Bujňáková et al. 2022; Dekker et al. 2021), eventually causing infections (Lynch et al. 2021; Irfan et al. 2022; Tilahun et al. 2021). VFs encompass diverse mechanisms, such as host cell adhesion (e.g., pili and adhesins), tissue invasion (e.g., invasins and hyaluronidase), toxin production (e.g., exotoxins and endotoxins), immune evasion (e.g., capsules and antigenic variation), enzymes facilitating spread (e.g., coagulase and proteases), iron acquisition (e.g., siderophores), secretion systems (e.g., Type III/VI), intracellular survival strategies, and antiphagocytic (Bujňáková et al. 2022; Leitão 2020; Liu et al. 2022a). Surveillance studies on VFs among Enterobacteriaceae revealed an abundance of VFs in ARE (Bujňáková et al. 2022; Leitão 2020; Payne and Neilands 1988; Pakbin et al. 2021).
Both ARGs and VFs are harbored within bacterial chromosomes and mobile elements such as plasmids, transposons, and integrons (Lynch et al. 2021; Aleshina et al. 2024; Partridge 2015; Toombs‐Ruane et al. 2017). Present data reveal that they are usually borne on plasmids (Hawkey et al. 2022; Carattoli et al. 2021), the dominant mobile genetic elements in antibiotic‐resistant bacteria (Harris et al. 2023; Wang and Dagan 2024). Incompatibility complex (I‐complex) plasmids, mostly consisting of resistance and fertility groups, are prevalent in ARE (Hawkey et al. 2022; Rozwandowicz et al. 2018; Atala 2020). IncF, IncC, and IncX have been reported to be the most I‐complex plasmids involved in the transmission of AMR among *Enterobacteriaceae (*Carattoli et al. 2021; Ma et al. 2023; Yu et al. 2022 Nov 16). In addition to I‐complex plasmids, Enterobacteriaceae also harbor colicin plasmids. These plasmids produce bacteriocins, which have been reported to inhibit other bacteria and are implicated in ARGs and VFs transmission (Ghanim et al. 2023). They are usually present in Enterobacteriaceae, especially E. coli and Klebsiella spp (Atala 2020; Ghanim et al. 2023; Marković et al. 2022a).
Previous studies have uncovered a high record of Enterobacteriaceae infections in the Cape Coast Teaching Hospital (CCTH) (Dakorah et al. 2022; Anning et al. 2022); however, data on the genetic basis of ARE, especially those that are ESBL‐producing, are revealed at the hospital due to the poor antimicrobial resistance surveillance in Ghana. This study combined phenotypic and genomic approaches to comprehensively characterize the AMR profiles of Enterobacteriaceae isolated from patients at CCTH, providing robust genetic evidence to support existing surveillance data. Through phenotypic identification and antimicrobial susceptibility testing (AST), we determined the prevalence of multidrug‐resistant (MDR) strains, extended‐spectrum β‐lactamase (ESBL) producers, and CRE. Additionally, whole genome sequencing (WGS) of 20 ESBL‐producing isolates was performed to characterize their taxonomy, antimicrobial resistance genes (ARGs), VFs, and harbored plasmid replicons.
Materials and Methods
2
Study Design and Site
2.1
This was a retrospective study of archived Enterobacteriaceae isolates from patients attending CCTH, a tertiary‐level health facility located in the Cape Coast Metropolitan Area of the Central Region of Ghana. The hospital is a 400‐bed capacity facility that serves as a referral center for the central, western, western east, and eastern regions of Ghana. The microbiology laboratory at the hospital routinely conducts culture and sensitivity testing for microorganisms isolated from various specimen sources, including blood, urine, abscess, stool, wound swabs, and high vaginal and endocervical swabs. Specimens received from the various departments within the hospital, including Enterobacteriaceae isolates archived from 2020 to 2023, were included in this study. The departments included were outpatient department (OPD), pediatric ward, emergency wards, intensive care unit (ICU), and obstetrics and gynecology (O/G).
Bacterial Isolates
2.2
Bacterial isolates of the Enterobacteriaceae family were retrieved from storage and phenotypically identified using standard biochemical tests, including citrate, oxidase, indole, urease, and triple sugar iron (TSI).
Antimicrobial Susceptibility Testing (AST)
2.3
AST was performed for the isolates using the following antimicrobial agents: ß‐lactams: ampicillin [AMP 10 mcg], ceftriaxone [CTR, 30 mcg], cefotaxime [CTX 30 mcg], cefuroxime [CXM 30 mcg], and meropenem [MRP, 10 mcg]; fluoroquinolones: ciprofloxacin [CIP 5 mcg], levofloxacin [LEV 5 mcg], and ofloxacin [OF 5 mcg]; aminoglycosides: amikacin [AMK 30 mcg] and gentamicin [GEN 10 mcg]; sulfonamide: cotrimoxazole [COT 25 mcg]; tetracycline [TET 30 mcg]; and chloramphenicol [CHL 30 mcg]. AST was performed using the Kirby–Bauer disk diffusion method, and the breakpoints were interpreted according to the Clinical and Laboratory Standard Institute (CLSI) 2020 guidelines (CLSI 2020). Briefly, pure colonies of the cultured isolates were picked from agar culture plates and inoculated into peptone broth to achieve turbidity equivalent to 0.5 McFarland standards. Using a sterile cotton bud, a swab of the bacterial suspension was streaked onto the entire surface of a freshly prepared Mueller–Hinton agar (MHA) plate. Antibiotic discs were placed on the MHA in the plates within 15 min of bacterial inoculation. The plates were then incubated at 37°C for 24 h. Subsequently, the zone of inhibition was measured in millimeters using a meter rule and interpreted as sensitive, intermediate, or resistant according to the CLSI 2020 guideline (CLSI 2020). Based on the AST findings, we classified the isolates resistant to 3 or more classes of antibiotics as MDR (Magiorakos et al. 2012) and ESBL‐producing according to the CSLI 2020 guideline.
ESBL Producers Screening
2.4
Isolates exhibiting an inhibitory zone diameter ≤ 27 mm upon exposure to cefotaxime (30 mcg) and ≤ 25 mm when tested with ceftriaxone (CTR: 30 mcg) according to the (CLSI 2020) guide were identified as possible ESBL producers. ESBL confirmatory tests were performed (n = 79) using ceftazidime (CAZ:10 mcg) and ceftazidime‐clavulanic acid (CAL: 40 mcg). Isolates that exhibited an inhibitory zone diameter of ≤ 17 mm upon exposure to ceftazidime (30 mcg) and difference around CAL to CAZ ≥ 5 mm when compared were confirmed as ESBL producers.
Whole Genome Sequencing (WGS)
2.5
Twenty ESBL‐producing isolates were selected for WGS. Genomic DNA (gDNA) was extracted from the bacteria using the Quick‐DNA Mini Prep Plus Kit™ (Zymo Research, Irvine®, United States) according to the manufacturer's instructions. WGS was performed at the WACCBIP NGS Laboratory (Accra, Ghana) using an Illumina MiSeq® (Illumina, Inc.®, San Diego, USA). DNA quality and quantity were determined using a Qubit™ 4.0 fluorometer (Thermo Fisher Scientific®, Waltham, USA). Short read sequencing was performed using (2 × 250 bp) paired end (PE) sequencing with the MiSeq® Reagent Kit v3 (600 cycles). For each sample, 100 ng total DNA was used for library preparation. Sequencing libraries were prepared from the enriched DNA using the Illumina® DNA Prep (Illumina) library preparation kit from the enriched DNA, following the manufacturer's instructions. Using the Nextera XT Index Kit v2™ (Illumina), distinct indices and Illumina sequencing adapters were attached to each library according to the manufacturer's instructions. Subsequently, each library was purified using Agencourt AMPure XP beads (Beckman Coulter®). Agilent 4200 TapeStation™ (Agilent®) was then used to check the expected size distribution and quality of the library. The library concentrations were measured using a Qubit™ 4.0 fluorometer (Life Technologies™). Based on the TapeStation and Qubit results, the barcoded libraries were normalized and pooled at an equimolar concentration. The combined library was diluted to 18 pM, spiked with 5% Phix™ (v3), and then sequenced at a depth of 50X and a coverage of 30X.
Bioinformatics Analyses
2.6
Adapters and low‐quality reads of (n = 20) PE reads were trimmed using Trimmomatic (v0.39) (https://github.com/usadellab/Trimmomatic), setting Phred score at 33, ILLUMINACLIP: Nextera‐PE. fa: 2:30:10, LEADING:3, TRAILING:3 and MINLEN:22. The PE trimmed reads (n = 20) were assembled using SPAdes Genome Assembler (v4.0.0) (https://github.com/ablab/spades), and quality assessment was performed using QUAST (v5.2.0) (https://github.com/ablab/quast). Multilocus sequence typing (MLST) (https://github.com/tseemann/mlst), which scans contigs against traditional PubMLST typing schemes in addition to ribosomal MLST (https://pubmlst.org/species-id), was performed to validate the identity of the isolates. Concurrently, each draft genome was screened for the presence of AMR genes using Abricate (https://github.com/tseemann/abricate), which hosts (CARD (v3.2.6), PlasmidFinder, and VFDBS) (accessed June 2024). The resistance and virulence genes were determined using CARD (Alcock et al. 2020; Jia et al. 2017) and VFDBS (Liu et al. 2022a), respectively. Assembled contigs were further assessed to identify plasmid genes using PlasmidFinder (Carattoli et al. 2014) to understand the transmission of AMR genes within the Enterobacteriaceae population at the hospital. Genes were selected based on coverage and identity equal to or greater than a threshold of 95%.
Data Analysis
2.7
Data visualization was conducted using Python (v3.11) in Jupyter (v3.6.3) via the Anaconda Navigator (v2.4.6), employing Matplotlib, pandas, plotly express, and seaborn libraries. Heatmaps illustrating antibiotic susceptibility patterns, sunburst presenting ARGs found in isolates and against antibiotics, and radar plot depicting virulence factor distributions among Enterobacteriaceae isolates were generated using the same libraries. Statistical analyses were performed using Jamovi (v2.3.28). Descriptive statistics summarized patient demographics and clinical characteristics, with median and interquartile ranges. Cohen's Kappa was used to evaluate interrater reliability between phenotypic, MLST, and rMLST identification methods, implemented via the ClinicoPath module in Jamovi.
Results
3
Socio‐Demographics and Clinical Data
3.1
The majority of bacterial isolates (70%) were from females, with 34% and 36% being within the age categories of 20–29 and 40–69, respectively. The ages ranged from 8 days to 85 years, with a median age of 40 years and an interquartile range of 35 years. Of the 100 samples, 51% and 10% were specimens referred from the OPD and accident and emergency department (A&E), respectively. Other departments, including the intensive care unit (ICU), female surgical ward (FSW), male surgical ward (MSW), and obstetrics and gynecology (O/G), contributed to 13% of the isolates. The majority of the isolates (44%) were obtained from urine samples as presented in Table 1.
Antimicrobial Susceptibility Patterns
3.2
Of the hundred (100) isolates, 6% were MDR. ESBL testing was successful for 79 isolates, out of which 74(93.7%) were ESBL‐producing. Approximately 93%, 69%, and 63% of the 100 isolates were susceptible to amikacin, chloramphenicol, and gentamicin, respectively. However, all isolates were resistant to ampicillin (Figure 1). Meropenem sensitivity testing was performed for the 79 isolates, of which 40.51% (32/79), 48.10% (38/79), and 11.39% (9/79) were susceptible, intermediate, or resistant, respectively. This suggests that the isolates are mostly susceptible to meropenem in the hospital.
Antibiotic susceptibility profiles (phenotype) of Enterobacteriaceae bacteria Isolates. The heatmap displays antibiotic susceptibility patterns across various Enterobacteriaceae isolates for different antibiotics. Each row represents a specific antibiotic, while each column corresponds to an isolate. The color‐coding indicates the susceptibility categories: black: susceptible (S), red: intermediate resistance (I), beige: resistant (R), gray: data not available (N/A). The antibiotics tested were (ß‐lactams: Ampicillin [Amp], Ceftriaxone [CTR], Cefotaxime [CTX], Ceftazidime [CAZ], Meropenem [MRP], and Cefuroxime [CXM]; Fluoroquinolones: Ciprofloxacin [CIP], Levofloxacin [LEV], and Ofloxacin [OF]; Aminoglycosides: Amikacin [AMK] and Gentamicin [GEN]; Sulfonamides: Cotrimoxazole [COT]; Tetracycline [TET]; Chloramphenicol [CHL]; and Ceftazidime‐Clavulanic acid [CAL]).
Whole Genome Assembly
3.3
Computation of the total read count and quality metrics of the assemblies (Table A5) was consistent and of high quality, with mean number of contigs, N50, and assembly size as 647, 212,022, and 7,025,968, respectively. Tseeman/mlst analysis predicted E. coli (8/20, 40%), K. pneumoniae (8/20, 40%), E. cloacae (2/20, 10%), and S. enterica (1/20, 5%). Additionally, (1/20, 5%) isolate initially identified as E. coli by biochemical tests was identified as Pseudomonas aeruginosa, which belongs to the family Pseudomonadaceae (Table A1). Agreement between the phenotypic and genotypic identification of the Enterobacteriaceae isolates was moderate (0.60), with a kappa value of 0.47. The statistical difference between the phenotypic and genotypic identification of the Enterobacteriaceae isolates was significant (p‐value < 0.001). This suggests that the phenotypic and genotypic methods, including MLST and rMLST, do not consistently agree with their classification of bacterial isolates.
Resistome
3.4
CARD revealed 139 unique ARGs that would confer resistance to the following classes of antibiotics: ß‐lactams (64/139), fluoroquinolones (45/139), aminoglycosides (23/139), phenicol (27/139), tetracycline (37/139), and sulfonamide (9/139) (Figure 3; Table A3). High diversity of ARGs was observed in E. coli (130/139), followed by K. pneumoniae (111/139), E. cloacae (9/139), and S. enterica (8/139) (Figure 4, Table A3). Isolate IDs BC82 (E. coli) harbored the most ARGs (74/139), whereas the lowest was detected in BC94 (K. pneumoniae) (6/139). CTX‐M‐15, an ESBL gene, was the most commonly observed ARG, detected in (13/19) isolates (Table A2). Nine ARGs, including *APH (3')‐Ia, Erm (*Ares‐Arroyo et al. 2021 ), EC‐15, Escherichia_coli_emrE, golS, mdsC, MdtK, QnrD1, and SHV‐80, were recorded as the least common occurring ARGs among the isolates. Each was observed once in a single isolate. Several variants of MDR genes (91/139) were observed in the isolates, especially in E. coli (65/91), followed by K. pneumoniae (44/91), S. enterica (5/91), and E. cloacae (1/91). They were observed to have conferred resistance, especially to 4 classes of antibiotics, including ß‐lactams, fluoroquinolones, tetracycline, and chloramphenicol.
Virulome
3.5
Using VFDBS, several VFs previously described in the Enterobacteriaceae family (E. coli, K. pneumoniae, E. cloacae, and S. enterica) were detected. In total, 414 unique virulence genes were detected (Table A4) and categorized into 8 major groups and 17 subcategories, as detailed in Table 2 and Figure 2. The predominant VF categories included adhesins, invasins, effector delivery systems, exotoxins, immune evasins, nutrient and metabolic factors, biofilms, and exoenzymes. Most of these VFs were observed in E. coli isolates, followed by K. pneumoniae, E. cloacae, and S. enterica. Adhesin‐associated VFs, which are essential for bacterial attachment to host cell mucosae (Abraham et al. 1988), were predominantly identified in E. coli and K. pneumoniae. Invasion‐related VFs, which facilitate host cell penetration, were detected in all four genera. Effector delivery systems, including T2SS and T3SS, are known for injecting bacterial proteins into host cells to manipulate immune signaling, cell death, and nutrient acquisition pathways (Nans et al. 2015; Pinaud et al. 2018) and were also observed in all the genera, principally in the S. enterica isolate, as shown in Figure 2. Exotoxins, particularly membrane‐acting, intracellular active toxins, were observed in E. coli, K. pneumoniae, and E. cloacae isolates; however, none were observed in the S. enterica isolate. Similarly, immune evasion VFs, including antiphagocytosis and complement evasion mechanisms, were present in all Enterobacteriaceae isolates except E. cloacae. Nutritional and metabolic VFs were mostly present in E. coli, followed by K. pneumoniae and E. cloacae. These factors, particularly those involved in metal uptake, metabolic adaptation, and iron sequestration, are crucial for bacterial survival and proliferation in host environments (Payne and Neilands 1988; Shealy et al. 2021). Biofilm‐associated VFs, which contribute to antibiotic resistance, are confined to E. coli and K. pneumoniae. Meanwhile, proteases that degrade exogenous proteins, enhancing bacterial growth and metabolism (Liu et al. 2022b), were uniquely observed in E. cloacae. Bacterial survival in host environments plays an important role in persistent infections (Grant and Hung 2013). Enterobacteriaceae strains harboring these VFs can subvert the host immune defenses, leading to recurrent infections (Bujňáková et al. 2022; Folgori et al. 2021). The diversity of VFs identified in the Enterobacteriaceae isolates accentuates their adaptability within host environments, enhancing their ability to evade antimicrobial defenses and establish infections.
Virulence factor distribution among Enterobacteriaceae bacterial species. This radar chart illustrates the distribution of eight virulence factors (Adherence, Invasion, Effector delivery system, Exotoxin, Immune modulation, Nutritional/Metabolic factor, Biofilm, and Exoenzyme) across four Enterobacteriaceae bacterial species: E. coli (green), K. pneumoniae (blue), S. enterica (orange), and E. cloacae (pink). Each axis represents a virulence factor, with the distance from the center showing its strength or pathogenic and AMR contribution in each species. E. coli stands out in Adherence and Nutritional/Metabolic factors, while S. enterica is strong in the Effector delivery system. The other species show varying levels of these factors, reflecting their different pathogenic profiles.
Distribution of ARGs conferring resistance to major antibiotic classes. The inner ring represents each class, and the outer ring represents individual ARGs. ARGs conferring resistance against β‐lactams were the most represented (Sawa et al. 2020), followed by fluoroquinolones (Shealy et al. 2021) and tetracyclines (CLSI 2020) (see Table A3).
Distribution of ARGs among bacterial isolates, with the inner circle representing species, the middle circle showing individual isolates, and the outer circle depicting detected ARGs. The visualization highlights E. coli and K. pneumoniae as the predominant species, each harboring diverse ARGs (see Table A3).
Plasmidome
3.6
The results from Plasmid Finder revealed a diverse array of plasmid types classified under Incompatibility (Inc) complex plasmids and colicin (Col) plasmids (Table 3). The number of plasmid replicons per isolate significantly varied. K. pneumoniae exhibited the highest diversity of plasmids (18/26), followed by E. coli (9/26) and E. cloacae (5/26). The highest plasmid diversity was observed in isolate BC37 from pneumoniae (8/26), whereas BC82 (E. coli) and BC26 (K. pneumoniae) exhibited the lowest (1/26 each). Only S. enterica in this collection harbored no plasmid replicons. Among the Inc plasmids that are mostly observed in Enterobacteriaceae as vehicles for AMR transmission (Rozwandowicz et al. 2018), the IncF subgroups were the most frequently detected. IncFIB (AP001918) _1 was the most prevalent, identified in eight isolates, followed by IncFIB(K)_1 and IncFIA_1, which were each found in 7 isolates. IncFII_1, IncHI1B(pNDM‐MAR) _1, and IncR_1 were detected in 3 isolates each. Plasmids such as *IncC_1, IncFII (*Rozwandowicz et al. 2018 ) _1, IncFII(pCoo)_1, IncX4_2, and pKPC‐CAV1321_1 were found in only a single isolate each. Colicin plasmids encoding colicins (bacteriocins), proteins that kill other bacteria (Atala 2020), were also distributed across the isolates. They have been reported to contribute to the spread of antibiotic resistance and other VFs among uropathogenic E. coli strains and Klebsiella spp by co‐integration with other plasmids such as IncC, IncF, and IncN (Ghanim et al. 2023; Ares‐Arroyo et al. 2021). Col156_1 was the most frequently detected gene (Tacconelli et al. 2018). ColRNAI_1 was present in 3 isolates, whereas Col (BS512) _1 and Col440I_1 were each observed in 2 isolates. The least common Colicin plasmids, found in single isolates, included Col (KPHS6) _1, Col3M_1, and ColpVC_1.
Discussion
4
Phenotypic surveillance of bacterial infection in clinical settings complemented with genomic studies enables elucidation of the basis of disease cause, accurate identification, transmission dynamics, and AMR (Ahmed et al. 2024; Govender 2022). Our study highlights the need to adapt genetic surveillance to complement routine phenotypic investigations, especially for the accurate identification of bacteria and their resistance to antibiotics in resource‐limited settings. We noticed a discordance between phenotypic and genotypic identification of the bacterial isolates. For example, some E. coli and K. Pneumoniae were phenotypically misidentified as Citrobacter spp and an E. cloaca as S. enterica. Rosenthal et al. have posited that bacterial misidentification can arise from laboratory procedural errors, such as culture media contamination or the use of suboptimal reagents (e.g., dried agar and expired substrates), which skew biochemical test results (Rosenthal et al. 1978). Additionally, evolutionary mutations in bacteria, driven by selective pressures such as antibiotics or nutrient scarcity, can alter phenotypic changes via genetic mutations or horizontal gene transfer, thereby confounding precise taxonomic identification (Blount et al. 2020). Misidentification of bacteria in the clinical setting could result in deleterious clinical outcomes, a phenomenon that can be forestalled by complementary genomic profiling (Heyman et al. 2025; Liang 2020). Additionally, genomic characterization can reveal the genetic basis of AMRs even for drugs for which AST was not performed. Also, VFs and plasmid identities from genomic profiling can be used to predict the pathogenicity potential and transmissibility of AMRs, respectively (Heyman et al. 2025; Arber 2000).
ESBL‐producing Enterobacteriaceae have been implicated in the outbreak of several bacteria‐borne diseases, including food‐borne diseases and nosocomial and care‐home outbreaks (Ibrahim et al. 2023; Do Tran et al. 2022; van Bilsen et al. 2021). In our study, 93% of the previously suspected ESBL producers were confirmed to be ESBL producers. Some hospital‐based studies in Cape Coast, Ghana (Sampah et al. 2023), and other African countries (Asare Yeboah et al. 2024; Bezabih et al. 2022b; Fadare and Okoh 2021) have reported similar rates of ESBL‐producing Enterobacteriaceae, especially in E. coli and K. pneumoniae. However, a lower rate of 14.6% was observed in a hospital‐based study conducted in Ontario, Canada (Hasan et al. 2023). The ESBL‐producing isolates sequenced harbored 17 individual ß‐lactamase ARGs of Ambler groups A, B, C, and D. ß‐lactamases associated with ESBLs observed were mainly SHV, TEM, and CTX‐M type genes found in the Ambler group A. These variants are mostly penicillinases and cephalosporinases and tend to be resistant to clavulanate, sulbactam, and tazobactam (Sawa et al. 2020; Bush and Jacoby 2010). This could explain why the addition of clavulanate to ceftazidime did not improve the sensitivity profile during AST. CTX‐M‐15, a member of the Ambler class A, was the predominant ß‐lactamase ARG among the isolates, accounting for 68.4% (13/19). Previous studies in Ghana have reported CTX‐M‐15 prevalence of over 80% in Enterobacteriaceae (Obeng‐Nkrumah et al. 2023; Eibach et al. 2016), whereas Asare Yeboah et al (Asare Yeboah et al. 2024) reported a lower proportion of 52%. Globally, CTX*‐M‐15* is the most prevalent ß‐lactamase ARG in Enterobacteriaceae, with a pooled prevalence of 16%, which is lower than 74% (14/19) observed in our study due to widespread dissemination of IncF plasmids that often carry this gene. We also observed the presence of oxacillinases, including OXA‐1, OXA‐58, and OXA‐402, which also confer ESBL activities (Sawa et al. 2020). Generally, most isolates were sensitive to carbapenems, with just above 11% of the isolates (9/79) phenotypically resistant to meropenem. However, only 2 of these resistant isolates, both K. pneumoniae (BC71 and BC75), were part of the whole genome sequenced isolates. These K. pneumoniae isolates harbored NDM‐1 carbapenemase. NDM‐1, an Ambler class B ß‐lactamase, has previously been described in E. coli clinical isolates and hospital environment surveillance studies (Ayibieke et al. 2018; Acolatse et al. 2022).
The majority of VFs identified in the WGS isolates were in adherence, effector delivery systems, and nutritional or metabolic factor families, mostly in E. coli and K. pneumoniae. However, only S. enterica harbored most effector delivery systems. Studies conducted in both Ghana and Slovakia (Bujňáková et al. 2022; Dekker et al. 2021) observed that the prevalence of these 3 VFs among Enterobacteriaceae, especially E. coli, K. pneumoniae, E. cloacae, and S. enterica, was high. Other studies in Africa and some European countries corroborate our findings (Leitão 2020; Dougnon et al. 2020; Hetsa et al. 2024; Amaretti et al. 2020). In our study, the whole genome sequenced isolates included 11 isolates, that is, 3 K. pneumoniae and 8 E. coli, which were isolated from urine infections. Generally, the ability of Enterobacteriaceae to colonize and cause urinary tract infections depends on their ability to form biofilms and produce several VFs, especially adherence factors (P fimbriae, S fimbriae, type I fimbriae), iron acquisition and sequestration (aerobactin and enterobactins), hemolysin, and cytotoxic necrotizing factors (cnf1) (Assouma et al. 2023; Govindarajan and Kandaswamy 2022).
Fimbriae are an absolute prerequisite for urinary tract colonization and disease causation, because Enterobacteriaceae uses that to adhere to host cells and form protective biofilms in the urinary tract, contributing to the pathogenesis and persistence of urinary tract infections (Connell et al. 1996; Hornick et al. 1991). Incidentally, the E. coli and K. pneumoniae sequenced possessed numerous fimbrial genes, including type 1, type 2 (P fimbriae), type 3, and S fimbriae. Specifically, E. coli harbored type I fimbriae (fimA, B, I, D, E G, H, K, F, T, U, V), P fimbriae (papB, C, F, G, H, D, F, J, K, X), and S fimbriae (sfaD, E, F, C, G, H, S, X, Y). Studies in uropathogenic Enterobacteriaceae conducted in Benin, Romania, Mongolia, and Egypt have also reported these fimbrial genes (Assouma et al. 2023). Uropathogenic K. pneumoniae uses two main types of fimbriae for adhesion to the uroepithelium, type 1 and type III (Stahlhut et al. 2012). However, we observed type II fimbriae (papF, papK) in the 3 uropathogenic K. pneumoniae isolates in addition to type I fimbriae (fimA, B, D, E, F, G, H, I, K) and type III fimbriae (mrkC, I, F, A, D, B, H, D).
In addition to these adherence factors, E. coli isolates also harbored iron acquisition and sequestration genes, including aerobactins (iucA, iucB, iutA) and enterobactins (entA, entC, entE, entF, entS, entB). The 3 K. pneumoniae also possessed an aerobactin (iutA.) and enterobactins (entF, entA, entE, entC, entS). Siderophores, such as enterobactins and aerobactins, enable uropathogenic Enterobacteriaceae to thrive in the host by scavenging iron from the host epithelial cells (Qi and Han 2018). According to Moxley et al. (Payne and Neilands 1988; Moxley 2022), Enterobacteriaceae requires iron for growth and virulence. Hemolysin (hlyC, hlyD) and cnf1, which E. coli uses to damage cell nutrients and trigger siderophores to sequester iron from the host for growth (Sung et al. 2024), were also observed among the E. coli isolates. Nhu et al. reported that hemolysins (hlyC, D) and cnf1 were found abundantly in UPEC strains, emphasizing their significance capacity to cause UTI (Nhu et al. 2019).
According to Nordmann and Poirel (2005); Algarni et al. (2024), most ARGs and VFs are borne on mobile genetic elements, especially plasmids, and have become important for the dissemination of these factors within the Enterobacteriaceae family through horizontal or vertical gene transfer (Rozwandowicz et al. 2018; Atala 2020; Ma et al. 2023; Algarni et al. 2024). It has been reported that most plasmid replicons found in Enterobacteriaceae are I‐complex (Incompatibility complex) plasmids and are mostly resistance and fertility plasmids in addition to colicinogenic plasmid (Carattoli et al. 2021; Rozwandowicz et al. 2018; Yu et al. 2022 Nov 16; Benz et al. 2021). In our study, the I‐complex we observed most was the IncF (13/19 total Inc plasmids) (Rozwandowicz et al. 2018; Naseer and Sundsfjord 2011; Stein et al. 2024). These plasmids have been reported worldwide for the dissemination of AMR genes, especially ESBLs, and particularly CTX‐M‐15 (Rozwandowicz et al. 2018; Naseer and Sundsfjord 2011; Stein et al. 2024). Unsurprisingly, CTX‐M‐15 was the most common ARG identified in our study. In our study, these complex groups of plasmid replicons were observed in E. coli and K. pneumoniae, similar to previous reports (Rozwandowicz et al. 2018; Stein et al. 2024). Other Inc plasmid replicons observed in our study included IncC, IncH, IncI, IncR, and IncX.
Consistent with previous investigations, our study also adds to the expanding body of evidence indicating that colicins co‐harbor with ARGs to facilitate AMR co‐selection and dissemination (Marković et al. 2022b; Daniels A. O et al. 2023; Chérier et al. 2021).
This study characterizes the resistome and virulome of Enterobacteriaceae isolates at CCTH, highlighting the pressing threat posed by ESBL‐producing Enterobacteriaceae to healthcare delivery. Furthermore, it highlights plasmid replicons associated with AMR gene transmission, emphasizing the role of genomic data in informing accurate diagnosis and rational antibiotic use.
Conclusions
5
Our study revealed alarming levels of MDR determinants, particularly against β‐lactam antibiotics, in the isolates. This represents one of the first in‐depth genomic assessments of ESBL‐producing Enterobacteriaceae in Ghana. Nearly all isolates were confirmed to be ESBL producers. Genomic analyses confirmed that the selected isolates were ESBL gene carriers. It also revealed 139 distinct resistance determinants across diverse classes of drugs, especially β‐lactams. CTX‐M‐15 β‐lactamase was the predominant ESBL gene, and NDM‐1 carbapenemase was detected in K. pneumoniae isolates, signaling the emergence of carbapenem resistance at CCTH. These pathogens harbored numerous VFs (fimbrial adhesins, siderophores, hemolysins, and secretion system components) and diverse plasmid replicons (especially, IncF and colicin plasmids), facilitating resistance dissemination. In aggregate, these findings provide robust genomic evidence of pervasive ESBL‐mediated resistance and emergent carbapenemase activity in Ghanaian Enterobacteriaceae, highlighting the urgent need for enhanced antimicrobial stewardship, routine genomic surveillance, and robust infection control measures to mitigate this growing public health threat in the Cape Coast Metropolis and other parts of Ghana.
Limitations
6
The phenotypic resistance assessment was limited to disc diffusion methods, mainly constrained by logistical limitations. Our findings potentially do not fully capture the genetic assessment of the plasmids of the individual isolates due to the usage of Illumina short reads, which could lead to incomplete reconstructions of plasmids or misassembles. It should be noted that our study circumscribed only those Enterobacteriaceae isolates accessible from the bacteriology laboratory at CCTH and with the availability of socio‐demographic and clinical data. Thus, our phenotypic AMR assessment does not fully capture the available Enterobacteriaceae isolates present at the time of this study at the hospital. Also due to funding constraints only 20 out of the 100 isolates were whole genome sequenced; our findings potentially do not fully capture the AMR genotypic assessment of all the retrieved isolates.
Author Contributions
Bismark Donkor: conceptualization; methodology; data curation; investigation; formal analysis; visualization; writing – review and editing; writing – original draft; software; validation. Richael Odarkor Mills: methodology; investigation; writing – review and editing; validation; formal analysis. Philimon Mwintige: investigation; methodology. Alberta Bedford Moses: investigation; methodology. Abigail Asmah Brown: investigation; methodology. Faustina Halm‐Lai: conceptualization; writing – review and editing; supervision; investigation; project administration; resources. Oheneba Charles Kofi Hagan: conceptualization; investigation; funding acquisition; writing – original draft; writing – review and editing; visualization; validation; methodology; formal analysis; project administration; supervision; data curation; resources; software.
Ethics Statement
Ethical approval for this study was obtained from the Cape Coast Teaching Hospital Ethical Clearance Review Committee (CCTHERC), Cape Coast, Ghana, under the approval number CCTHERC/EC/2023/102. To ensure privacy and confidentiality, data were retrieved deidentified.
Conflicts of Interest
The authors declare that there are no personal relationships that could have influenced the work reported in this study.
Supporting information
Table A1.
Table A2.
Table A3.
Table A4.
Table A5.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abraham, S. N. , D. Sun , J. B. Dale , and E. H. Beachey . 1988. “Conservation of the D‐Mannose‐Adhesion Protein Among Type 1 Fimbriated Members of the Family Enterobacteriaceae .” Nature 336: 682–684.2904657 10.1038/336682 a 0 · doi ↗ · pubmed ↗
- 2Acolatse, J. E. E. , E. A. R. Portal , I. Boostrom , et al. 2022. “Environmental Surveillance of ESBL and Carbapenemase‐Producing Gram‐Negative Bacteria in a Ghanaian Tertiary Hospital.” Antimicrobial Resistance & Infection Control 11, no. 1: 49.35296353 10.1186/s 13756-022-01090-2PMC 8925048 · doi ↗ · pubmed ↗
- 3Ahmed, S. K. , S. Hussein , K. Qurbani , et al. 2024. “Antimicrobial Resistance: Impacts, Challenges, and Future Prospects.” Journal of Medicine, Surgery, and Public Health 2: 100081.
- 4Alcock, B. P. , A. R. Raphenya , T. T. Y. Lau , et al. 2020. “CARD 2020: Antibiotic Resistome Surveillance With the Comprehensive Antibiotic Resistance Database.” Nucleic Acids Research 48, no. D 1: 517.31665441 10.1093/nar/gkz 935PMC 7145624 · doi ↗ · pubmed ↗
- 5Aleshina, Y. , A. Yeleussizova , A. Mendybayeva , P. Shevchenko , and R. Rychshanova . 2024. “Prevalence and Antimicrobial Resistance of Enterobacteriaceae in the North of Kazakhstan.” Open Veterinary Journal 14, no. 2: 604–616.38549569 10.5455/OVJ.2024.v 14.i 2.1PMC 10970123 · doi ↗ · pubmed ↗
- 6Algarni, S. , D. D. Gudeta , J. Han , R. Nayak , and S. L. Foley . 2024. “Genotypic Analyses of Inc HI 2 Plasmids From Enteric Bacteria.” Scientific Reports 14, no. 1: 9802.38684834 10.1038/s 41598-024-59870-2PMC 11058233 · doi ↗ · pubmed ↗
- 7Amaretti, A. , L. Righini , F. Candeliere , et al. 2020. “Antibiotic Resistance, Virulence Factors, Phenotyping, and Genotyping of Non‐Escherichia coli Enterobacterales From the Gut Microbiota of Healthy Subjects.” International Journal of Molecular Sciences 21, no. 5: 1847.32156029 10.3390/ijms 21051847 PMC 7084377 · doi ↗ · pubmed ↗
- 8Anning, A. S. , E. Baah , S. D. Buabeng , et al. 2022. “Prevalence and Antimicrobial Resistance Patterns of Microbes Isolated From Individuals Attending Private Diagnostic Centre in Cape Coast Metropolis of Ghana.” Scientific Reports 12, no. 1: 14282.35995932 10.1038/s 41598-022-18595-w PMC 9395395 · doi ↗ · pubmed ↗