Rare skin disease helps uncover mechanisms of acyl ceramide biosynthesis
Susanne Tollinger, Daniela Ortner, Thomas Trafoier, Verena Moosbrugger-Martinz, Robert Gruber, Matthias Schmuth

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
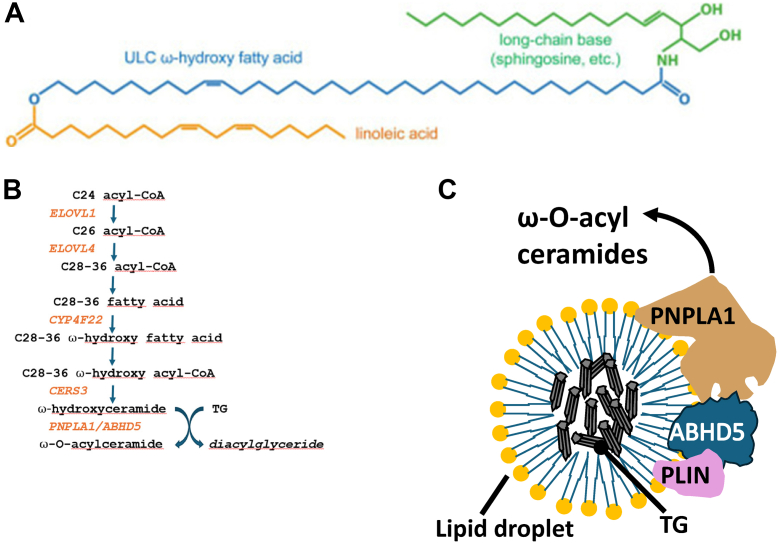 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSphingolipid Metabolism and Signaling · Polyamine Metabolism and Applications · Lipid metabolism and biosynthesis
Omega-O-acyl ceramides in the epidermis are composed of three different carbon chains. A long chain base, most often sphingosine, is combined with an omega-hydroxylated ultra-long-chain fatty acid (28–36 carbon atoms) and with linoleic acid (Fig. 1A). Thus, as opposed to most ceramides, omega-O-acyl ceramides have three rather than two hydrophobic chains and contain ultra-long fatty acids, unique to the outer layers of the epidermis. The work of Schratter et al. (1) published in this issue of the Journal of Lipid Research illustrates how we can learn from genetic ABHD5 (1-acylglycerol-3-phosphate O-acyltransferase) variants about the molecular and spatial requirements for omega-O-acyl ceramide synthesis mediating epidermal permeability barrier function.Fig. 1. Omega-O-acyl ceramide structure, biosynthesis, and colocalization of enzymes and cofactors in proximity to lipid droplets. A: Structure of omega-O-acyl ceramide. Modified from the study by Kien et al. (2). B: Current knowledge of the biosynthesis pathway. C: Putative scheme of the proximity between PNPLA1, ABHD5, lipid droplets, and perilipin (PLIN). Modified from the study by Schratter et al. (1). ABHD5, 1-acylglycerol-3-phosphate O-acyltransferase, also known as comparative gene identification-58 (CGI-58); C, carbon number; CERS3, ceramide synthase 3; CYP4F22, cytochrome P450, family 4, subfamily F, polypeptide 22; ELOV, elongation of very-long-chain fatty acid protein; PNPLA1, patatin-like phospholipase domain-containing 1; TG, triglyceride; ω, omega.
The biosynthesis of omega-O-acyl ceramides from fatty acid to acyl ceramide requires multiple steps (Fig. 1B). Several of these steps are associated with genetic variants in humans and cause disease when mutated. Whereas ELOVL1 and ELOVL4 variants, which are responsible for elongation to ultra-long fatty acids, cause both skin (scaling) and neurologic abnormalities (paraplegia, ataxia, brain atrophy), CYP4F22, CERS3, and PNPLA1 (patatin-like phospholipase domain-containing 1) variants result in skin disease only, that is, excessive scaling of the skin. Interestingly, abnormalities in the most distal step of the pathway (Fig. 1B) can either be caused by PNPLA1 variant or ABHD5 variant and are associated with a similar scaly skin phenotype. All these entities are characterized by deficiency in omega-O-acyl ceramides and impaired permeability barrier function (2, 3).
Abnormal barrier function is a feature of genodermatoses, termed epidermal differentiation disorders (EDD) (4). This group of diseases exhibits thickened, scaly skin, often accompanied by inflammation, due to erroneous keratinocyte differentiation. Physiologically, epidermal differentiation results in the coordinated formation of a stratum corneum, the outermost skin layer. Essential components of this barrier are i) scaffolding structures; that is, the cornified envelope, surrounded by ii) highly organized lipid lamellae consisting of cholesterol, sphingomyelin/ceramide, and free fatty acids (5), including a critical complement of tissue-specific, very long-chain N-acyl fatty acids (6). An optimal composition of skin lipids prevents the ingress of harmful external substances and at the same time blocks uncontrolled fluid loss to the outside. If omega-O-acyl ceramide is not generated in sufficient quantities and positioned in the correct location, its ability to form a protein-bound ultra-long-chain-ceramide coating on the external surface of the cornified envelope leads to abnormal stratum corneum lipid composition and structure and impaired permeability barrier function (7, 8).
Subjects with ABHD5-syndromic (s)EDD or Chanarin-Dorfman syndrome (9), in contrast to those with PNPLA1 variants, additionally exhibit pathologies in other organs, due to a lack of breakdown of triacylglycerol to free fatty acids, resulting in the accumulation of lipid droplets in specific organs. The organ involvement defines the disease as syndromic (hearing abnormalities and increased liver and muscle function parameters) (4), correlating with the role of ABHD5 in lipid metabolism.
Schratter et al. (1) tested the effects of various naturally occurring missense mutations in ABHD5 by combining coimmunoprecipitation with solid-phase interaction assays and immunofluorescence live-cell imaging. The authors demonstrate that the proximity of ABHD5 with lipid droplets, specifically perilipins, and simultaneous enzymatic activation of PNPLA1 is critical (Fig. 1C). This work demonstrates that ABHD5 has multiple functions. It modulates the spatial proximity to lipid droplets, and it serves as a cofactor for PNPLA1.
Table 1 by Schratter et al. (1) illustrates the variability of the disease phenotype with different ABHD5 variants. Genetic alterations affecting different parts of the 3-dimensional molecular structure of ABHD5 have differential effects, which explain phenotype variability between subjects with ABHD5-sEDD. Some individuals have more severe liver involvement, whereas others have predominant skin involvement. A recent report even described a variant that causes skin disease only, with no other organ involvement (10). Schratter et al. (1) suggested that this is likely due to different molecular surfaces being involved in mediating the local positioning of ABHD5 in different ways. Alternatively, multiple PNPLA enzymes, for example, PNPLA3, or other, hitherto unrecognized additional components of lipid droplet complexes may interact differently with ABHD5 when it is genetically altered.Table 1. Naturally occurring variants in genes relevant for omega-O-acyl ceramide synthesisLipid enzymeFunctionNonsyndromicSyndromicDisease phenotypeELOVL fatty acid elongase 1Synthesis of C26 fatty acids and sphingolipidsNoYespEDD, migratory erythematous scaling of the skin, spastic paraplegia, and dysmorphic faciesELOVL fatty acid elongase 4Synthesis of very-long-chain polyunsaturated fatty acidsNoYesErythrokeratodermatous plaques, spastic paraplegia, developmental delay, and sometimes brain atrophyCYP4F22Catalyzes ULCFA ω-hydroxylationYesNoSkin thickening and generalized fine scaling, can spare the trunk, sometimes self-improvingCERS3Catalyzes the formation of ceramides from sphingoid base and acyl-coA substratesYesNoErythema, generalized fine scalingPNPLA1Transacylase transferring linoleic acid from triglycerides to omega-hydroxyceramidesYesNoErythema, fine scales on flexor and extensor surfaces and facial skin, formerly known as erythrokertodermia variabilis et progressiveABHD5Interactions with members of the perilipin (PLIN) and patatin-like phospholipase domain-containing protein (PNPLA) protein families enhancing linoleic acid transesterificationYesYesErythema, generalized fine scaling, ataxia, myopathy, hearing loss, and alopeciaABHD5, 1-acylglycerol-3-phosphate O-acyltransferase, also known as comparative gene identification-58 (CGI-58); C26, carbon number 26; CERS3, ceramide synthase 3; CYP4F22, cytochrome P450, family 4, subfamily F, polypeptide 22; LOV, elongation of very-long-chain fatty acid protein; pEDD, palmoplantar epidermal differentiation disorder; PNPLA1, patatin-like phospholipase domain-containing 1; ULCFA, ultra-long-chain fatty acid.
By studying a rare disease, this work makes a valuable contribution to the understanding of EDD pathogenesis and the biochemistry of omega-O-acyl ceramide synthesis. The results reported by Schratter et al. should motivate the study of additional naturally occurring genetic variants relevant for the various steps in the pathway to further elucidate the biochemistry of ceramide synthesis. A continued search for mechanisms of disease pathogenesis in these disorders will provide us with a better understanding of the pathways and potential new therapeutic approaches.
While one logical approach to treating ABHD5 deficiency is to apply the missing lipids exogenously, that is, acyl ceramide, the present work suggests that pharmaceutical compounds modulating the spatial positioning of ABHD5 in relation to PNPLA1 in the vicinity of lipid droplets may be another therapeutic option. Finally, gene transfer or gene editing would be promising, but in syndromic disease, the intervention needs to take place at early developmental stages. In summary, a better understanding of lipid droplet formation and composition in EDD may improve our ability to treat these disorders (11).
Conflict of interest
The authors declare that they have no conflicts of interest with the contents of this article.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Schratter M.Holubek D.Koeffler L.Züllig T.Eichmann T.O.Wolinski H.Defective targeting of PNPLA 1 to lipid droplets causes ichthyosis in ABHD 5-syndromic epidermal differentiation disorder J. Lipid. Res.66202510087510.1016/j.jlr.2025.100875 PMC 1246503740818613 · doi ↗ · pubmed ↗
- 2Kien B.Grond S.Haemmerle G.Lass A.Eichmann T.O.Radner F.P.W.ABHD 5 stimulates PNPLA 1-mediated ω-O-acylceramide biosynthesis essential for a functional skin permeability barrier J. Lipid. Res.592018236023673036141010.1194/jlr.M 089771 PMC 6277169 · doi ↗ · pubmed ↗
- 3Meyer J.M.Boeglin W.E.Brash A.R.Recombinant PNPLA 1 catalyzes the synthesis of acylceramides and acyl acids with selective incorporation of linoleic acid J. Lipid. Res.64202310037910.1016/j.jlr.2023.100379 PMC 1020901837087101 · doi ↗ · pubmed ↗
- 4Paller A.S.Teng J.Mazereeuw-Hautier J.Hernández-MartínÁ.Tournier C.G.Hovnanian A.Syndromic epidermal differentiation disorders: new classification towards pathogenesis-based therapy Br. J. Dermatol.19320255926184018449610.1093/bjd/ljaf 123 · doi ↗ · pubmed ↗
- 5Feingold K.R.Elias P.M.The role of ceramides in the disruption of the cutaneous permeability barrier, a common manifestation of skin disorders J. Lipid. Res.65202410059310.1016/j.jlr.2024.100593 PMC 1134843139002948 · doi ↗ · pubmed ↗
- 6Yamaji M.Ohno Y.Shimada M.Kihara A.Alteration of epidermal lipid composition as a result of deficiency in the magnesium transporter Nipal 4J. Lipid. Res.65202410055010.1016/j.jlr.2024.100550 PMC 1115324238692573 · doi ↗ · pubmed ↗
- 7Uchida Y.Cho Y.Moradian S.Kim J.Nakajima K.Crumrine D.Neutral lipid storage leads to acylceramide deficiency, likely contributing to the pathogenesis of Dorfman-Chanarin syndrome J. Invest. Dermatol.1302010249724992052062910.1038/jid.2010.145 · doi ↗ · pubmed ↗
- 8Demerjian M.Crumrine D.A.Milstone L.M.Williams M.L.Elias P.M.Barrier dysfunction and pathogenesis of neutral lipid storage disease with ichthyosis (Chanarin-Dorfman syndrome)J. Invest. Dermatol.1262006203220381674151610.1038/sj.jid.5700332 · doi ↗ · pubmed ↗