Complex lipids shape cell growth and communication in moss
Blanca Jazmín Reyes-Hernández

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
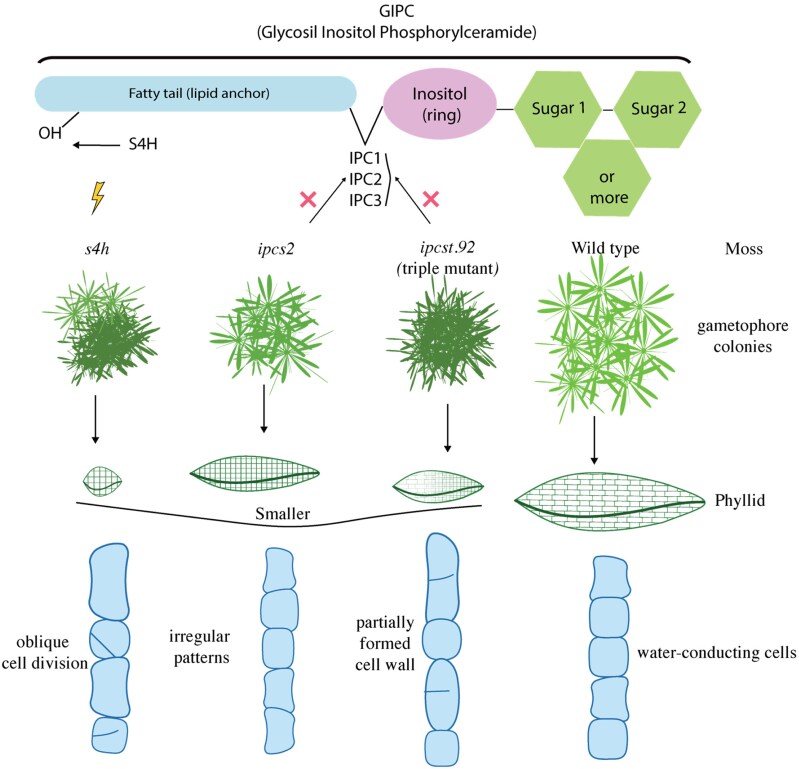 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBryophyte Studies and Records · Lichen and fungal ecology · Botany and Plant Ecology Studies
Biological membranes are mainly built from 3 classes of lipids: glycerophospholipids, sterols, and sphingolipids. Together, they provide structure, regulate fluidity, and create specialized domains for signaling and interactions. In plants, one of the most important group of sphingolipids is the glycosyl inositol phosphorylceramides (GIPCs). They are highly abundant in the plasma membrane and play unique structural and functional roles (Gronnier et al. 2016). Deficiency in GIPCs generally leads to visible growth defects, while plants completely missing GIPCs cannot be recovered (Rennie et al. 2014), showing that GIPCs are important for plant physiology and development.
GIPCs are a special type of plant lipid made of 3 main parts: a fatty tail, a small ring-shaped molecule called inositol, and one or more sugars attached (Fig.). The fatty part anchors the molecule into the membrane, while the inositol and sugars extend outward, where they can interact with the environment. Plants can decorate GIPCs with different sugars, sometimes 1 or 2, sometimes many, creating a remarkable variety of structures. This sugar diversity may allow GIPCs to perform specialized roles in different tissues. Yet, understanding their precise functions remains difficult because their biosynthesis is tightly linked to other sphingolipids, making it hard to separate the effects of GIPCs depletion from broader imbalances in lipid metabolism.
To address this challenge, Wagner and collaborators (2025) recently reported in Plant Physiology the use of the moss Physcomitrium patens (P. patens) as a model to study GIPCs. The moss offers unique advantages such as making precise genetic edits without passing through a reproductive cycle, which allows the maintenance of mutants with severe developmental defects (Schaefer and Zrÿd 2001). The moss has a simple body, with filamentous protonema and single-cell-thick leaf-like phyllids, making it ideal for tracking cell division, differentiation, and growth. Lipid profiling has revealed a rich diversity of GIPCs in P. patens, including variants with different sugar types such as hexoses (Hex), N-acetyl hexoses (HexNAc), and pentoses (Pent) (Resemann et al. 2019). Together, these features make P. patens a powerful system for investigating the physiological functions of GIPCs.
Last year, Haslam et al. (2024) described several P. patens mutants affecting GIPCs biosynthesis. One was an ipcs2 single mutant with reduced Hex-GIPCs, and another was a more severe triple mutant (ipcst.92, affected in IPCS1, IPCS2, and IPCS3 genes) showing stronger losses of Hex-GIPCs than ipcs2 and Hex-Hex-GIPCs. These mutants are affected in the enzyme Inositol Phosphorylceramide Synthase (IPCS), which produces the direct precursor of GIPCs, Inositol Phosphorylceramide (IPC). Interestingly, they also reported a third mutant named s4h (sphinganine C4-hydroxylase), altered in an enzyme that modifies the sphingoid base backbone (Fig.). This mutant accumulated 5 to 6 times more complex GIPCs (Gömann et al. 2021).
In their current work, the authors examined the growth of gametophore colonies (the leafy shoots) of those P. patens mutants and studied cell division in their leaf-like phyllids on their stem-like structure. They found strong defects in cell division in the triple ipcst.92 and s4h mutants, as well as malformed plasmodesmata, the microscopic channels that connect the cytoplasm of neighboring cells (Zanini and Burch-Smith 2024). Because plasmodesmata control communication between cells, defects in their structure or maturation often disrupt developmental coordination and stress responses. These defects were further explored with assays of protein and molecule movement between cells, showing how sphingolipids are closely linked to fundamental aspects of cell biology.
In the wild-type moss, plasmodesmata normally mature from pore to branched form, and the 3 mutants analyzed by the authors failed to mature. Functional assays confirmed that these structural changes impacted transport. For example, in the triple mutant ipcst.92 there was uncontrolled diffusion of molecules through incomplete walls, while s4h mutant restricted protein movement despite retaining plasmodesmata pore form. Together, these results highlight how GIPCs are critical for plasmodesmata maturation and cell-to-cell communication.
Lipid profiling performed on ipcst2 and ipcst.92 confirmed that as more disrupted IPCS isoforms are present in P. patens, the level of GIPCs decreases progressively; this correlates with the severity of phenotype. The same analysis of the s4h mutant showed that S4H enzyme is important to keep the balance of complex GIPCs (determined by number of sugars attached), which also leads to severe developmental abnormalities. These findings demonstrate that both the quantity and balance of GIPCs are essential for plant development.
Morphological analysis supported these findings. Using side-by-side growth assays of colonies, the authors showed that the ipcs2 mutant had smaller colonies than the wild type, and the reduction in colony size was even more reduced in ipcst.92 and s4h mutants (by 90%). Although all these mutants had dwarf, dense, and rounder shapes than wild type, interestingly, they presented more dry mass. Together, these findings reflected that altered sphingolipid metabolism does not simply regulate plant size but also determines colony shape, compactness, and structural integrity.
Further examination of phyllids revealed that the 3 mutants analyzed had fewer cells than wild type, while ipcst.92 and s4h showed more severe defects. These mutants had less and shorter margin cells and had poorly developed conducting tissue, showing that GIPCs are needed for coordinated differentiation and organ formation. Transmission electron microscopy further highlighted defects like the incomplete walls with unanchored cell plates in the triple mutant ipcst.92 and irregular callose-like deposits in s4h. Moreover, all mutants displayed plasma membrane detachment from the cell wall, with distinct patterns depending on the mutation.
In summary, this study shows that GIPCs are not only building blocks of membranes but also key players in regulating cell division, wall formation, and intercellular communication. By using P. patens as a complementary model to vascular plants, the work uncovers roles for sphingolipids that extend beyond what is known from studies of Arabidopsis. A particularly intriguing observation is that GIPCs-deficient moss mutants display defects like Arabidopsis mutants lacking PI4P (Lin et al. 2019), even though these lipids have been proposed to act in opposition (Ito et al. 2021). This proposed opposition raises important questions about how the balance between GIPCs and PI4P is maintained and how shifts in this equilibrium might influence other membrane lipids. Could differences in lineage-specific cell division mechanisms explain why moss and Arabidopsis respond differently to sphingolipid disruption? And do environmental cues reshape GIPC–PI4P relationships to adjust plant growth and signaling? Or even how exactly do GIPCs stabilize plasmodesmata during maturation? Addressing these questions will be essential to integrate lipid biochemistry with developmental outcomes and to understand how plants fine-tune connectivity across evolutionary scales.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gömann J, Herrfurth C, Zienkiewicz A, Ischebeck T, Haslam TM, Hornung E, Feussner I. Sphingolipid long-chain base hydroxylation influences plant growth and callose deposition in physcomitrium patens. New Phytol. 2021:231(1):297–314. 10.1111/nph.1734533720428 · doi ↗ · pubmed ↗
- 2Gronnier J, Germain V, Gouguet P, Cacas J-L, Mongrand S. GIPC: glycosyl inositol phospho ceramides, the major sphingolipids on earth. Plant Signal Behav. 2016:11(4):e 1152438. 10.1080/15592324.2016.1152438 PMC 488392127074617 · doi ↗ · pubmed ↗
- 3Haslam TM, Herrfurth C, Feussner I. Diverse INOSITOL PHOSPHORYLCERAMIDE SYNTHASE mutant alleles of physcomitrium patens offer new insight into complex sphingolipid metabolism. New Phytol. 2024:242(3):1189–1205. 10.1111/nph.1966738523559 · doi ↗ · pubmed ↗
- 4Ito Y, Esnay N, Platre MP, Wattelet-Boyer V, Noack LC, Fougère L, Menzel W, Claverol S, Fouillen L, Moreau P, et al Sphingolipids mediate polar sorting of PIN 2 through phosphoinositide consumption at the trans-Golgi network. Nat Commun. 2021:12(1):4267. 10.1038/s 41467-021-24548-034257291 PMC 8277843 · doi ↗ · pubmed ↗
- 5Lin F, Krishnamoorthy P, Schubert V, Hause G, Heilmann M, Heilmann I. A dual role for cell plate-associated PI 4Kβ in endocytosis and phragmoplast dynamics during plant somatic cytokinesis. EMBO J. 2019:38(4):e 100303. 10.15252/embj.201810030330617084 PMC 6376452 · doi ↗ · pubmed ↗
- 6Rennie EA, Ebert B, Miles GP, Cahoon RE, Christiansen KM, Stonebloom S, Khatab H, Twell D, Petzold CJ, Adams PD, et al Identification of a sphingolipid α-glucuronosyltransferase that is essential for pollen function in Arabidopsis. Plant Cell. 2014:26(8):3314–3325. 10.1105/tpc.114.129171 PMC 437183125122154 · doi ↗ · pubmed ↗
- 7Resemann HC, Lewandowska M, Gï Mann J, Feussner I. Membrane lipids, waxes and oxylipins in the moss model organism physcomitrella patens. Plant Cell Physiol. 2019:60(6):1166–1175. 10.1093/pcp/pcz 00630698763 PMC 6553664 · doi ↗ · pubmed ↗
- 8Schaefer D, Zrÿd J. The moss physcomitrella patens, now and then. Plant Physiol. 2001:127(4):1430–1438. 10.1104/PP.01078611743086 PMC 1540175 · doi ↗ · pubmed ↗