TTC21B variants disrupt the left-right asymmetry and pronephric development in zebrafish
Linxia Deng, Yuan Yang, Xiaoling Yin, Jing Yang, Yijie Duan, Kang Wang, Weicheng Duan, Yu Zhang, Bo Xiong, Jianhua Zhou

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
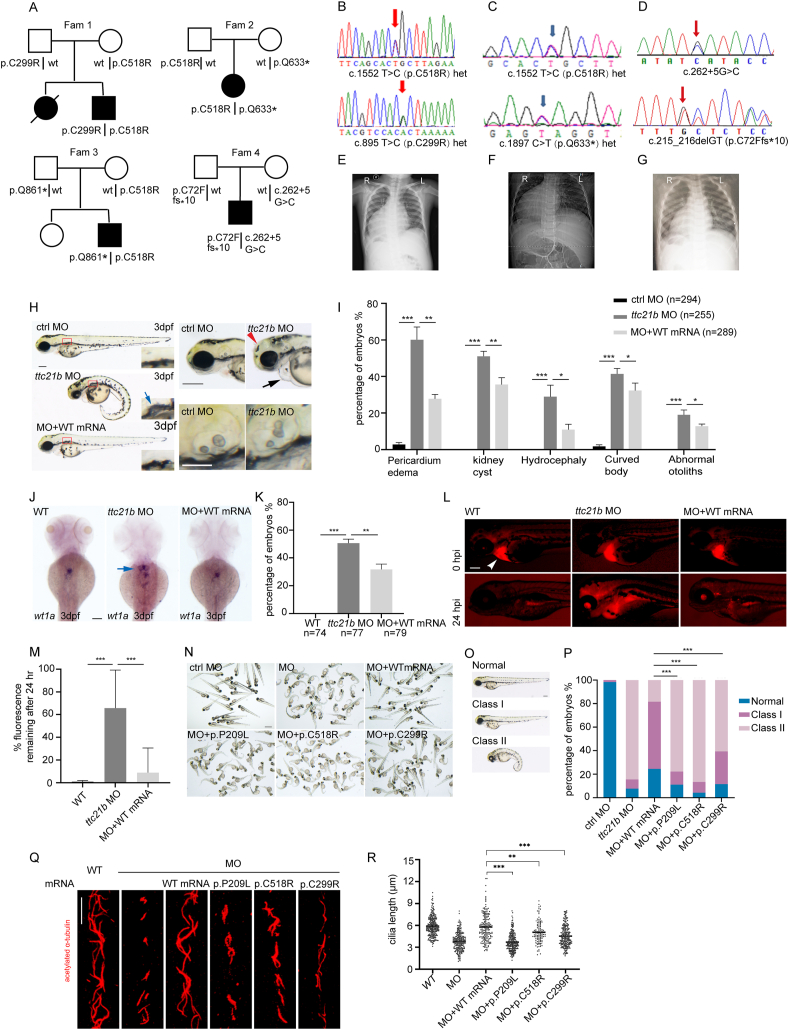 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCongenital heart defects research · Craniofacial Disorders and Treatments · Developmental Biology and Gene Regulation
Nephronophthisis (NPHP) is an autosomal recessive kidney disease and is the most prevalent monogenic cause of end-stage renal disease in childhood. The tetratricopeptide repeat domain 21B (TTC21B) gene encodes the ciliary protein intraflagellar transport protein 139 (IFT139) and has been recently implicated in heterogeneous diseases, including nephronophthisis type 12 (NPHP12), short-rib thoracic dysplasia 4 (SRTD4), and Joubert syndrome (JBTS).1^,^2 In Europe and North Africa, the prevalent TTC21B variant c.626C > T (p.P209L) has been associated with focal segmental glomerulosclerosis in adults.2^,^3 To date, only a limited number of TTC21B gene variants have been reported in Chinese individuals, predominantly presenting with infantile NPHP, which differs from the manifestations observed in Caucasian patients.4^,^5 In this study, we identified novel TTC21B gene variants in Chinese children with NPHP and investigated their role in left-right asymmetry and pronephric development.
Firstly, we analyzed 5 cases of NPHP12 from 4 non-consanguineous Chinese families, with detailed clinical manifestations summarized in supplementary results and Table S1. Utilizing exome sequencing, we identified 4 novel variants in the TTC21B gene: c.1897C > T (p.Q633∗), c.2581C > T (p.Q861∗), c.215_216delGT (p.C72Ffs∗10), and c.262+5G > C (Fig. 1A–D). Surprisingly, 3 out of 4 patients shared the missense variant c.1552T > C (p.C518R), which was also found in 14 other Chinese patients, suggesting that this variant may be a potential hotspot mutation in the Chinese population (Table S1 and Fig. 1A). A comprehensive review of 18 patients carrying the p.C518R variant revealed that the majority presented infantile NPHP and progressed to end-stage renal disease at an average age of 3.5 years (range: 12 days to 16 years). Clinical features included mild to severe proteinuria, hypertension, and extra-renal manifestations such as situs inversus (8/18), congenital heart defects (5/18), cirrhosis or hepatomegaly (4/18), splenomegaly (2/18), and brachydactyly or polydactyly (2/18). These observations suggest that the C518R variant likely substantially impairs the function of the IFT139 protein.Figure 1TTC21B variants cause nephronophthisis in children and disrupt the cilia and pronephric development in zebrafish. (A) Pedigrees of affected families with TTC21B variants. (B–D) Sequence chromatograms in family 1, 2, and 4. (E–G) The chest X-ray showed situs inversus, where the heart was on the right side. (E) The chest X-ray of the proband's sister of family 1. (F) The chest X-ray of the patient in family 2. (G) The chest X-ray of the patient in family 3. (H) Representative images of ttc21b morphants at 3 days post-fertilization (dpf) with curved body axis, pronephric cysts (blue arrow; scale bar, 200 μm), hydrocephalus (red arrowhead) and pericardial edema (black arrow; scale bar, 200 μm), and abnormal otoliths in otic vesicles (scale bar, 100 μm). (I) Statistical analysis of gross morphology of embryos injected with the indicated morpholinos (MOs) or mRNAs. (J) Pronephric cysts validated by whole-mount in situ hybridization with the podocyte differentiation marker wt1a. Scale bars, 100 μm. (K) Quantification of embryos with pronephric cysts. (L) The top and bottom panels indicate the embryos injected into the pericardium with rhodamine dextran at 0 h post-injection (hpi) and 24 hpi, respectively, in wild-type (WT) or ttc21b morphant or rescued embryos. The white arrowheads indicate the heart (scale bars, 100 μm). (M) Percentage fluorescent intensity remaining after 24 hpi in ttc21b morphants group versus WT group and WT mRNA rescued group. (N) A group image of 3 dpf morphology of WT embryos or embryos injected with ctrl MO, MO, or co-injected with MO and human WT TTC21B or mutant TTC21B mRNAs. Scale bars, 1000 μm. (O) Embryos were categorized by severity of the phenotype: the normal was categorized as no obvious change; class I indicated mild ciliopathy defects, including slight pericardial edema and/or pronephric cysts; class II referred to embryos that presented with severe ciliopathy defects, including severe pericardial edema or body curvature or hydrocephalus. Scale bars, 200 μm. (P) Stacked bar charts show the percentages of the normal, class I, and class II in different experimental groups. Co-injection of WT TTC21B mRNAs with MO resulted in significant rescue, while mutant TTC21B mRNAs exhibited attenuated rescue ability compared with WT mRNAs. (Q) Representative confocal images show whole-mount immunostaining of anti-acetylated tubulin antibody (red) labeled cilia in pronephric ducts at 27 hpf. Scale bars, 10 μm. (R) Histogram analysis of cilia length (measured by ImageJ software). Cilia's length was decreased in ttc21b morphant compared with the WT embryo. Whereas cilia defects were well rescued by co-injected with human WT TTC21B mRNAs, the variant TTC21B mRNAs failed to effectively rescue the defects. Data were represented as mean ± standard deviation. ∗P < 0.05, ∗∗P < 0.01, and ∗∗∗P < 0.001.Figure 1
Subsequently, we generated a zebrafish model using morpholino technology. Knockdown of the ttc21b gene resulted in typical manifestations of ciliopathy at 3 days post-fertilization (dpf), including a curved body axis, hydrocephalus, pericardial edema, pronephric cysts, and abnormal otoliths in otic vesicles (Fig. 1H). Whole-mount in situ hybridization analyses of Wilms tumor gene wt1a, a podocyte marker, confirmed the presence of pronephric cysts (Fig. 1J). To evaluate renal function in ttc21b morphants, rhodamine-labeled 10-kDa dextran was injected into the pericardium of 3-dpf embryos, and dextran excretions were calculated at 24 h post-injection (hpi). In wild-type (WT) embryos, nearly all the dextran was excreted by 24 hpi, while approximately 72.1% of dextran remained in the pericardial cavity of ttc21b morphants, indicating impaired clearance of low molecular weight components (Fig. 1L, M). Importantly, all ttc21b morpholino-induced defects were significantly rescued by overexpression of human WT TTC21B mRNA (Fig. 1I, K, M).
Next, we performed in vivo complementation tests to determine the pathogenicity of the TTC21B missense variants. We co-injected the ttc21b morpholinos with each human variant mRNA (p.P209L, p.C518R, and p.C299R) into zebrafish embryos and compared the rescue efficiency with WT TTC21B mRNA. Phenotypic alterations were classified into three categories: appear to be normal; mild ciliopathy defects (class I), such as slight pericardial edema and/or pronephric cysts; and severe ciliopathy phenotypes (class II), including pronounced pericardial edema, body curvature, or hydrocephalus (Fig. 1O). Notably, 84.4% of ttc21b morphants exhibited severe ciliopathy phenotypes, which was decreased to 18.4% by co-injection with WT TTC21B mRNA (Fig. 1N–P). Interestingly, the rescue efficacy of all variant mRNAs was significantly attenuated (Fig. 1N–P). Furthermore, we analyzed the role of ttc21b in ciliary formation. The results showed that the anterior pronephric cilia length was dramatically shortened in ttc21b morphants at 27 h post-fertilization (hpf) compared with WT embryos. Similarly, co-injection with WT TTC21B mRNA resulted in a complete restoration of cilia length, while the mutant TTC21B mRNAs only partially restored cilia length (Fig. 1Q, R). These results underscore the pathogenicity of the p.C518R and p.C299R variants in vivo.
Then, we performed whole-mount in situ hybridization using the cardiac-specific marker cardiac myosin light chain 2 (cmlc2) and the liver marker succinate-CoA ligase GDP-forming subunit beta (suclg2) to assess the impact of the ttc21b gene variants on the development of left-right asymmetry. Our findings revealed that 98.6% of control morpholino-injected embryos exhibited a leftward heart tube jog compared with 34.6% of ttc21b morphants (Fig. S1A, B). Consistent with the cmlc2 findings, a significant proportion of ttc21b morphants (42.4%) displayed a reversed suclg2 expression pattern (Fig. S1C, D). Moreover, we collected the 18-hpf and 19.5-hpf embryos to detect the expression patterns of the Nodal-related gene spaw and paired-like homeodomain transcription factor 2c (pitx2c), respectively, both of which are critical for situs-specific organogenesis. Our analysis showed that only 30.3% of ttc21b morphant embryos had left-side lateral plate mesoderm spaw expression, compared with 75.0% in controls (Fig. S1E, F). The expression pattern of pitx2c alteration after ttc21b ablation resembled that of the spaw (Fig. S1G, H). Moreover, these aberrant expression patterns observed in ttc21b morphants were significantly ameliorated by co-injection with WT TTC21B mRNA (Fig. S1F, H). Collectively, these findings demonstrate that ttc21b is required for the left-right patterning of internal organs during zebrafish embryogenesis.
Further, to evaluate the formation of Kupffer's vesicle (KV) cilia, which is crucial for the development of left-right asymmetric organs, ttc21b-morpholino was injected into one- or two-cell stage Tg(sox17:gfp) embryos, followed by immunostaining with anti-GFP and anti-acetylated tubulin antibodies at the 10-somite stage. Although the number of KV cilia remained unchanged, the average cilia length was significantly shorter in ttc21b morphants compared with WT embryos (Fig. S2A, D, E). Subsequently, examination of KV formation in live embryos at the 10-somite stage revealed a significant impact on KV morphogenesis due to ttc21b knockdown (Fig. S2B, F). Charon (DAN domain BMP antagonist family member 5/dand5) is the initial gene exhibiting asymmetric expression, influenced by the intensity and direction of KV flow during early embryonic development. In WT embryos, asymmetric charon expression was detected on the right side of the KV. Conversely, ttc21b morphants exhibited a disturbed expression pattern with a higher proportion on the left side and no directional bias (Fig. S2C, G). These findings highlight the critical role of ttc21b in zebrafish KV formation and fluid flow regulation.
To further substantiate the findings observed in ttc21b morphants, we generated ttc21b mutants with a 3-bp deletion and a 2-bp insertion in exon 3 using CRISPR-Cas9 (Fig. S3A). At 72 hpf, all homozygous mutants exhibited kidney cysts, with some embryos also exhibiting mild pericardial edema. These phenotypes were ameliorated by co-expressing WT TTC21B mRNA (Fig. S3B–F). Notably, body curvature and growth retardation were observed at 30 dpf (Fig. S3H). Additionally, the homozygous ttc21b mutants exhibited shorter cilia in the pronephric duct compared with their WT siblings (Fig. S3I, J). However, only 7% (4/57) of the ttc21b mutant embryos showed a no-loop heart, and 4.3% (2/45) presented with a right-sided liver at 72 hpf (Fig. S4A–D). Regrettably, most of the homozygous ttc21b mutants succumbed within 30 days (Fig. S3G), and those that survived to adulthood were sterile. Consequently, homozygous embryos could only be obtained by crossing heterozygous individuals. Therefore, this discrepancy may be attributed to the presence of the maternally derived WT ttc21b mRNAs. Collectively, these results indicate the crucial role of ttc21b in ciliogenesis and the development of left-right asymmetry.
In conclusion, our study has identified 4 novel TTC21B variants and indicated a hotspot variant (c.1552T > C, p.C518R) that appears to be unique in Chinese patients with NPHP12. The in vivo studies conducted in zebrafish have confirmed the pathogenicity of TTC21B missense variants and demonstrated a novel association of TTC21B in regulating the development of left-right asymmetry.
CRediT authorship contribution statement
Linxia Deng: Writing – original draft, Project administration, Methodology, Formal analysis, Data curation. Yuan Yang: Writing – original draft, Project administration, Data curation. Xiaoling Yin: Writing – original draft, Project administration, Formal analysis. Jing Yang: Writing – original draft, Project administration, Formal analysis. Yijie Duan: Formal analysis, Data curation. Kang Wang: Formal analysis, Data curation. Weicheng Duan: Data curation. Yu Zhang: Supervision, Conceptualization. Bo Xiong: Writing – review & editing, Supervision, Data curation, Conceptualization. Jianhua Zhou: Writing – review & editing, Supervision, Funding acquisition, Conceptualization.
Ethics declaration
This study was approved by the Human Ethics Committees of the Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology (approval number: No.TJ-IRB20220950). Written informed consent was obtained from participants or their parents.
Data availability
The data are available upon request from the authors.
Funding
This work was supported by the 10.13039/501100001809National Natural Science Foundation of China (No. 81873596), the Key Research and Development Program of Hubei Province, China (No. 2022BCA047), and the 10.13039/501100012166National Key Research and Development Program of China (No. 2022YFC2705102 and No. 2022YFC2705103).
Conflict of interests
The authors declared no competing interests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Tran P.V.Haycraft C.J.Besschetnova T.Y.THM 1 negatively modulates mouse sonic hedgehog signal transduction and affects retrograde intraflagellar transport in Cilia Nat Genet 40420084034101832725810.1038/ng.105PMC 4817720 · doi ↗ · pubmed ↗
- 2Davis E.E.Zhang Q.Liu Q.TTC 21B contributes both causal and modifying alleles across the ciliopathy spectrum Nat Genet 43320111891962125834110.1038/ng.756PMC 3071301 · doi ↗ · pubmed ↗
- 3Bullich G.Vargas I.Trujillano D.Contribution of the TTC 21B gene to glomerular and cystic kidney diseases Nephrol Dial Transplant 32120171511562694012510.1093/ndt/gfv 453 · doi ↗ · pubmed ↗
- 4Li Y.Dai L.Xu H.Clinical report and genetic analysis of rare premature infant nephronophthisis caused by biallelic TTC 21B variants Mol Genet Genomic Med 1232024 e 2399
- 5Zhang J.Sun L.Kuang X.Y.Clinical phenotype analysis of 6 cases of TTC 21B gene related nephronophthisis Zhonghua Er Ke Za Zhi 60820228208243592219510.3760/cma.j.cn 112140-20211223-01076 · doi ↗ · pubmed ↗