The ER-localized susceptibility factor RTP1 negatively regulates plant immunity
Josephine H R Maidment

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
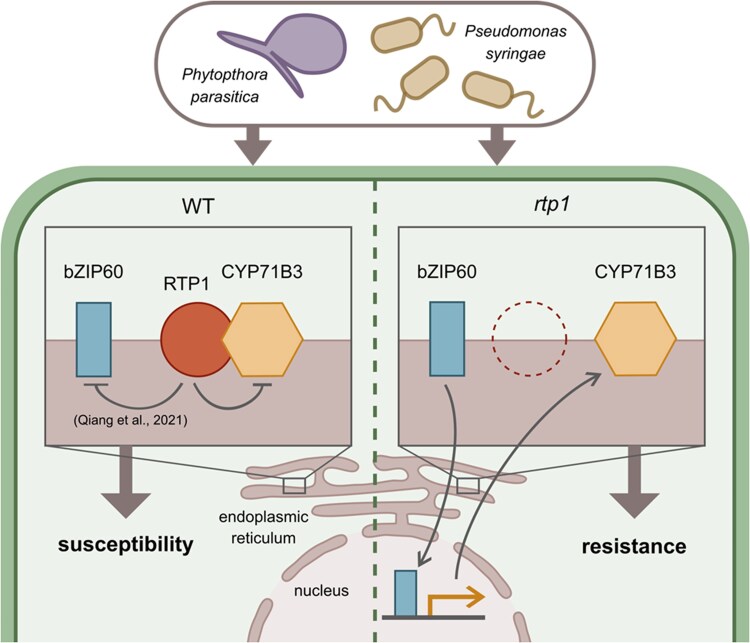 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-Microbe Interactions and Immunity · Plant pathogens and resistance mechanisms · Plant Parasitism and Resistance
Plant pests and pathogens reduce crop yields and quality (Savary et al. 2019). Resistant crop varieties typically contain 1 or more dominant resistance genes encoding receptors that detect molecular signatures of infection. Some, however, carry mutations in susceptibility (S) genes. S genes can be involved in enabling pathogen entry/establishment, negatively regulating plant immune responses, providing the pathogen with necessary nutrition, or facilitating movement through the host plant (Van Schie and Takken 2014).
Arabidopsis thaliana Resistance to Phytophthora parasitica 1 (RTP1) mediates susceptibility to biotrophic pathogens (Pan et al. 2016; Qiang et al. 2021). Belonging to the A. thaliana MtN21 gene family, RTP1 is similar to nodulin-related genes from Medicago truncatula. RTP1 contains 8 to 10 predicted transmembrane domains and localizes to endoplasmic reticulum (ER) membranes (Pan et al. 2016). Compared with wild-type (WT) A. thaliana plants, rtp1 mutants are more resistant to the oomycete P. parasitica, the fungus Golovinomyces cichoracearum, and the bacterial pathogen P. syringae pv tomato (Pst) DC3000 (Pan et al. 2016). Furthermore, rtp1 mutants show increased reactive oxygen species (ROS) production and accelerated defense gene expression (Pan et al. 2016), which are hallmarks of an immune response and indicate that RTP1 may be involved in negative regulation of plant defences.
Previously, it was reported that RTP1 acts on regulators of the unfolded protein response (UPR) (Qiang et al. 2021). Under stress conditions, such as pathogen infection, unfolded or misfolded proteins accumulate in the endoplasmic reticulum (ER). To resolve this situation and prevent cell death, plant cells activate signaling cascades collectively referred to as the UPR. The ER membrane–associated transcription factors bZIP28 and bZIP60 act as stress sensors in parallel signaling pathways to upregulate expression of UPR genes (Kwan Ko and Brandizzi 2024). Production of active bZIP60 is achieved by unconventional splicing of the bZIP60 mRNA; increased splicing of bZIP60 was observed in rtp1 mutants, suggesting that RTP1 is a negative regulator of bZIP60 activation. In addition, RTP1 appears to interact with and stabilize bZIP28, possibly leading to increased retention in the ER and reduced translocation to the nucleus (Qiang et al. 2021).
In a recent study published in Plant Physiology, Wei et al. (Wei et al. 2025) probed the function of RTP1 in greater detail. Comparative analysis of the transcriptomes of P. parasitica-infected roots of A. thaliana rtp1 mutants and WT plants revealed upregulation of several genes from the Cytochrome P450 family 71 (CYP71) in rtp1 mutants compared with WT plants. Cytochrome P450 (CYP) proteins are a diverse family of heme-containing enzymes that catalyze a wide range of reactions (Xu et al. 2015). CYP71 genes have been reported to have a function in pathogen resistance, with some known to be required for biosynthesis of the antimicrobial molecule camalexin. The authors focused on an uncharacterized member of the CYP71 family, CYP71B3, for further study. A. thaliana cyp71b3 mutants were more susceptible than WT Col-0 plants to both the oomycete P. parasitica and the bacterial pathogen Pst DC3000. In contrast, CYP71B3 overexpressing lines were more resistant to both pathogens, suggesting that CYP71B3 is a positive regulator of plant resistance to biotrophic plant pathogens (Wei et al. 2025).
After demonstrating that, like RTP1, GFP-tagged CYP71B3 is localized in the ER, the authors used split-luciferase and coimmunoprecipitation assays to investigate whether RTP1 binds to CYP71B3. The results suggested that the two proteins interact. By coexpressing CYP71B3 in N. benthamiana with either RTP1 or an empty vector (EV) control, the authors observed reduced accumulation of CYP71B3 in the presence of RTP1 compared with EV and concluded that RTP1 destabilizes CYP71B3 (Wei et al. 2025).
The authors subsequently aimed to identify regions of CYP71B3 involved in interaction with RTP1. Comparison of CYP71B3 with other A. thaliana CYP71 family members identified I38 as a residue exclusively present in RTP1-interacting CYP71 proteins. Wei et al. hypothesized that I38 was therefore important for RTP1 binding. Introduction of an I38A mutation into CYP71B3 resulted in a loss of the luciferase complementation signal observed when WT CYP71B3 was coexpressed with RTP1. Furthermore, unlike WT CYP71B3, CYP71B3^I38A^ could not genetically complement cyp71b3 mutants (Wei et al. 2025). However, it is yet to be determined whether CYP71B3^I38A^ lies at an RTP1 binding interface or whether this mutation affects the ER localization of the protein or alters its structure to render it nonfunctional.
Finally, the authors note that CYP71B3 expression is regulated by the transcription factor bZIP60 (Wei et al. 2025). The results presented here complement the previous study by Qiang et al. (2021) and collectively support a function for RTP1 as a regulator of UPR during biotic stress. As the precise enzymatic activity of CYP71B3 remains unknown, the metabolic consequences of upregulation of CYP71B3—and how this leads to enhanced resistance—remain to be explored.
Studies of susceptibility genes can give insights into how pathogens successfully cause disease. In addition, understanding at a molecular level how S genes condition host susceptibility opens new avenues for development of resistant crop varieties. While some S genes have been successfully deployed in the field, in other cases, knocking out S genes leads to pleiotropic phenotypes that prevent their use in an agricultural context (Garcia-Ruiz et al. 2021). Insights into the precise function of S gene products could enable targeted modification to alter their activity, expression, or interactions with other proteins (Garcia-Ruiz et al. 2021).
In summary, the work presented here extends our understanding of the role of RTP1 in modulating host plant susceptibility to biotrophic pathogens and implicates the previously uncharacterized CYP71B3 protein in regulating plant biotic stress responses (Figure). Functional characterisation of S genes provides insight into pathogen infection requirements and opportunities to develop novel genetic resistance to crop pathogens.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Garcia-Ruiz H, Szurek B, Van den Ackerveken G. Stop helping pathogens: engineering plant susceptibility genes for durable resistance. Curr Opin Biotechnol. 2021:70:187–195. 10.1016/j.copbio.2021.05.005PMC 887809434153774 · doi ↗ · pubmed ↗
- 2Kwan Ko D, Brandizzi F. Dynamics of ER stress-induced gene regulation in plants. Nat Rev Genet. 2024:25(7):513–525. 10.1038/s 41576-024-00710-438499769 PMC 11186725 · doi ↗ · pubmed ↗
- 3Pan Q, Cui B, Deng F, Quan J, Loake GJ, Shan W. RTP 1 encodes a novel endoplasmic reticulum (ER)-localized protein in Arabidopsis and negatively regulates resistance against biotrophic pathogens. New Phytol. 2016:209(4):1641–1654. 10.1111/nph.1370726484750 · doi ↗ · pubmed ↗
- 4Qiang X, Liu X, Wang X, Zheng Q, Kang L, Gao X, Wei Y, Wu W, Zhao H, Shan W. Susceptibility factor RTP 1 negatively regulates Phytophthora parasitica resistance via modulating UPR regulators b ZIP 60 and b ZIP 28. Plant Physiol. 2021:186(2):1269–1287. 10.1093/plphys/kiab 12633720348 PMC 8608195 · doi ↗ · pubmed ↗
- 5Savary S, Willocquet L, Pethybridge SJ, Esker P, Mc Roberts N, Nelson A. The global burden of pathogens and pests on major food crops. Nat Ecol Evol. 2019:3(3):430–439. 10.1038/s 41559-018-0793-y 30718852 · doi ↗ · pubmed ↗
- 6Van Schie CCN, Takken FLW. Susceptibility genes 101: how to be a good host. Annu Rev Phytopathol. 2014:52(1):551–581. 10.1146/annurev-phyto-102313-04585425001453 · doi ↗ · pubmed ↗
- 7Wei Y, Zong D, Tang Y, Kong L, Gao X, Wang X, Zhang Y, Yang Y, Qiang X, Shan W. RTP 1 negatively regulates plant immunity by interacting and destabilizing cytochrome P 450 protein CYP 71B 3 in Arabidopsis. Plant Physiol. 2025:198(3):kiaf 284. 10.1093/plphys/kiaf 28440592489 · doi ↗ · pubmed ↗
- 8Xu J, Wang XY, Guo WZ. The cytochrome P 450 superfamily: key players in plant development and defense. J Integr Agric. 2015:14(9):1673–1686. 10.1016/S 2095-3119(14)60980-1 · doi ↗