Beyond the Amazon: vector surveillance and emerging Oropouche virus in South America
Ruth Dálety da Silva Brito, Jéssica Pires Farias, Alexander Birbrair, Luís Carlos de Souza Ferreira, Paloma Oliveira Vidal, Jaime Henrique Amorim

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
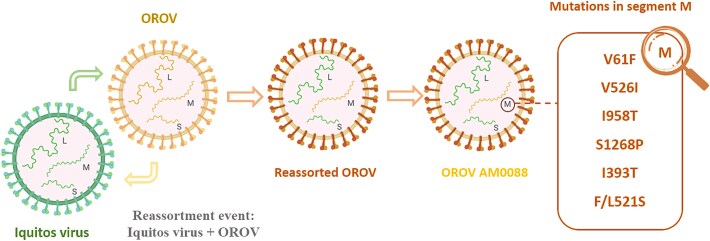 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-Borne Animal Diseases · Mosquito-borne diseases and control · Viral Infections and Vectors
To the Editor,
We commend the recent study by Vieira et al. (Vieira et al. 2024) for the innovative approach of integrating xeno-monitoring and metatranscriptomic sequencing to investigate arboviral diversity. This innovative approach not only revealed diverse co-circulating arboviruses in Australia but also exemplifies how next-generation surveillance can outperform conventional methods in ecologically complex regions. Tropical areas in South and Central America, especially the Amazon basin and surrounding biomes, share similar conditions for arboviral endemicity, vector heterogeneity, and underdiagnosis due to symptom overlap following infection with different co-circulating viruses (Files et al. 2022, Bai et al. 2025). These findings highlight the potential of genomic surveillance tools in arbovirus-endemic regions.
Of particular relevance stand neglected arboviruses such as the Oropouche virus (OROV), which is currently expanding its geographic and epidemiological footprint across the Americas (Gutierrez et al. 2020, Moreira et al. 2024). While historically endemic to the Amazon basin, OROV recently caused outbreaks in previously non-endemic areas of Brazil, including the northeast (e.g. Bahia State), the southeast (e.g. Espírito Santo State), and the south (e.g. Santa Catarina State), as well as in neighbouring countries, such as Peru and Panama (Moreira et al. 2024, Bai et al. 2025). These outbreaks were geographically distinct across multiple Brazilian states and neighbouring countries, with partially overlapping time windows during 2024. Recent genomic surveillance revealed that latest outbreaks are associated with a novel reassortant virus variant, whose small and large segments are more closely related to Iquitos virus, a related Orthobunyavirus, while the medium segment retains similarity to the OROV prototype strain (Gutierrez et al. 2020, Moreira et al. 2024). This reassortment event is accompanied by the emergence of non-synonymous mutations across different genomic segments (Fig. 1). In the M segment, substitutions V61F (in the Gn envelope glycoprotein), V526I, I958T, S1268P, and F/L521S (in the Gc envelope glycoprotein), as well as I393T in the nonstructural protein NSm, were identified. Mutations in the Gc envelope glycoprotein, which functions as the viral antireceptor, may influence host range and susceptibility by altering virus–receptor interactions. In the L segment, which encodes the RNA-dependent RNA polymerase (RdRp), substitutions such as I758V, I1942V, H476Y, E847G, and R346K were identified. These evolutionary and geographic events are summarized in Table 1. These amino acid changes may have phenotypic consequences, potentially affecting viral fitness, replication efficiency, and virulence (Moreira et al. 2024, Silva Júnior et al. 2024, Scachetti et al. 2025). Despite these alarming developments, OROV remains a neglected pathogen, with widespread underreporting due to limited molecular diagnostic capacity and lack of systematic vector surveillance programs (de Souza Luna et al. 2017, Bai et al. 2025). In addition, it is important to emphasize that in most places, vector surveillance is focused on mosquitoes and not midges. This diagnostic gap hinders early detection, impairs outbreak response, and masks the true disease burden, particularly in rural and peri-urban settings.
Effective surveillance of OROV must consider the diversity of vectors involved in the transmission cycle. Species such as Aedes serratus, Culex quinquefasciatus, Coquillettidia venezuelensis, and Mansonia venezuelensis have been found naturally infected in the field, indicating their participation in the virus’s sylvatic cycle. Laboratory studies further confirmed that mosquitoes, including A. serratus, Aedes scapularis, Aedes albopictus, C. quinquefasciatus, and Psorophora ferox, can be experimentally infected and are capable of transmitting OROV, with C. quinquefasciatus exhibiting the highest vector competence described to date. However, it is important to clarify that the actual epidemiological relevance of mosquitoes remains uncertain, and no species has yet demonstrated the consistent field-to-human transmission dynamics established for biting midges. Nevertheless, the biting midge Culicoides paraensis remains the primary vector historically associated with human outbreaks, with its competence firmly established through virus isolation in wild specimens and successful transmission to susceptible hosts under laboratory conditions (Bai et al. 2025). The possibility of expansion of vector range raises urgent questions about urban spillover and the risk of future outbreaks in densely populated regions. In this context, metatranscriptomic surveillance applied directly to field-collected vector pools—as implemented in the Australian study—offers a powerful, unbiased approach to detect emerging OROV variants, including reassortant variants and co-infections with other arboviruses. Such methods can uncover evolutionary trajectories, inter-species transmission events, and genomic signatures of adaptation that are otherwise missed by conventional virological assays (Gutierrez et al. 2020, Vieira et al. 2024). Integrating these techniques into routine vector monitoring would not only enhance early detection of OROV but also support more accurate modelling of its spatial dynamics and evolutionary potential.
The successful implementation of genomic arbovirus surveillance through xeno-monitoring and metatranscriptomics in Australia offers a timely and transferable model for Latin America, where arbovirus emergence is shaped by ecological pressures, human mobility, and climate change (Sah et al. 2024). We advocate for the urgent adaptation of such strategies to South American contexts, particularly in Brazil, where arboviruses like OROV, mayaro virus (MAYV), yellow fever virus (YFV), and others demonstrate the capacity to cause unexpected outbreaks through sylvatic-urban spillover events (Moreira et al. 2024, Bai et al. 2025). Establishing regional genomic surveillance networks—integrating public health laboratories, academic institutions, and field entomology teams—would not only enhance early warning capabilities but also foster data sharing and coordinated responses across national borders. This collaborative infrastructure is essential to detect, characterize, and contain emerging viral threats before they reach epidemic scale. In addition, broader impact modelling of metatranscriptomic implementation and comparative epidemiological insights across related arboviruses represent promising directions for future research. However, the implementation of such strategies in Brazil and neighbouring countries faces practical challenges. These include the high cost of sequencing, limited access to equipped laboratories, the need for specialized training, and barriers to data integration across institutions. Recognizing and addressing these constraints is essential for the success of genomic surveillance in the region. In addition, recent studies have emphasized that the resurgence of OROV poses a diagnostic challenge in the region (Cherem and Barçante 2025), with frequent misclassification as dengue, reinforcing the need for expanded genomic surveillance and improved differential diagnosis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bai F, Denyoh PMD, Urquhart C et al. A comprehensive review of the neglected and emerging Oropouche virus. Viruses 2025;17:439. 10.3390/v 1703043940143366 PMC 11945866 · doi ↗ · pubmed ↗
- 2Cherem J, de Barçante JM. Addressing the escalating burden of dengue in the Americas amid global challenges. J Infect Public Health 2025;18:102879, 1–2. 10.1016/j.jiph.2025.10287940614676 · doi ↗ · pubmed ↗
- 3de Souza Luna LK, Rodrigues AH, Santos RIM et al. Oropouche virus is detected in peripheral blood leukocytes from patients. J Med Virol 2017;89:1108–11. 10.1002/jmv.2472227787907 · doi ↗ · pubmed ↗
- 4Files MA, Hansen CA, Herrera VC et al. Baseline mapping of Oropouche virology, epidemiology, therapeutics, and vaccine research and development. npj Vaccines 2022;7:1–10. 10.1038/s 41541-022-00456-235301331 PMC 8931169 · doi ↗ · pubmed ↗
- 5Gutierrez B, Wise EL, Pullan ST et al. Evolutionary dynamics of Oropouche virus in South America. J Virol 2020;94:e 01127–19. 10.1128/JVI.01127-19PMC 702235331801869 · doi ↗ · pubmed ↗
- 6Moreira FRR, Dutra JVR, de Carvalho AHB et al. Oropouche virus genomic surveillance in Brazil. Lancet Infect Dis 20240;24:e 664–6. 10.1016/S 1473-3099(24)00558-939208828 · doi ↗ · pubmed ↗
- 7Sah R, Srivastava S, Kumar S et al. Oropouche fever outbreak in Brazil: an emerging concern in Latin America. Lancet Microbe 2024;5:100904. 10.1016/S 2666-5247(24)00136-838971172 · doi ↗ · pubmed ↗
- 8Scachetti GC, Forato J, Claro IM et al. Re-emergence of Oropouche virus between 2023 and 2024 in Brazil: an observational epidemiological study. Lancet Infect Dis 2025;25:166–75. 10.1016/S 1473-3099(24)00619-439423838 PMC 11779697 · doi ↗ · pubmed ↗