Effects of a two-week modified ketogenic diet on circulating lipoprotein subclasses, GDF15, and FGF21 in obese adults
Nana Zhang, Na Liu, Guoxia Zhao, Juan Yan, Pinghua Zhang, Xiaomiao Li, Jie Zhou

TL;DR
A two-week modified ketogenic diet reduced weight and body fat in obese adults while changing specific blood markers and HDL subclasses.
Contribution
The study reveals novel metabolic effects of a modified ketogenic diet, including GDF15 elevation and FGF21 reduction, alongside beneficial changes in HDL subclasses.
Findings
A two-week modified ketogenic diet significantly reduced body weight and BMI in obese participants.
FGF21 levels decreased by 51.91%, while GDF15 levels increased by 5.76%.
HDL-c subfractions showed significant changes, with larger HDL-c subfractions increasing and smaller ones decreasing.
Abstract
As key metabolic regulators, the roles of GDF15 and FGF21 in mediating the effects of modified ketogenic diet (MKD) on weight loss and lipoprotein remodeling in obese patients require further investigation. This study enrolled 30 metabolically healthy obese participants (BMI ≥ 28 kg/m²) for a 2-week MKD intervention. Using a self-controlled pre-post design, we performed measurements including body composition analysis, fasting serum GDF15 and FGF21 levels measurement, and serum lipoprotein subclass quantification at both baseline and post-intervention time points. Following a 2-week MKD intervention, participants exhibited statistically significant reductions in body weight (96.14 ± 27.23 kg vs. 91.63 ± 26.47 kg; Δ4.8%, P < 0.001) and BMI (33.99 ± 6.08 kg/m2 vs. 32.41 ± 5.95 kg/m2; Δ4.7%, P < 0.001). Body fat parameters significantly improved, with body fat mass (BFM) and visceral fat…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
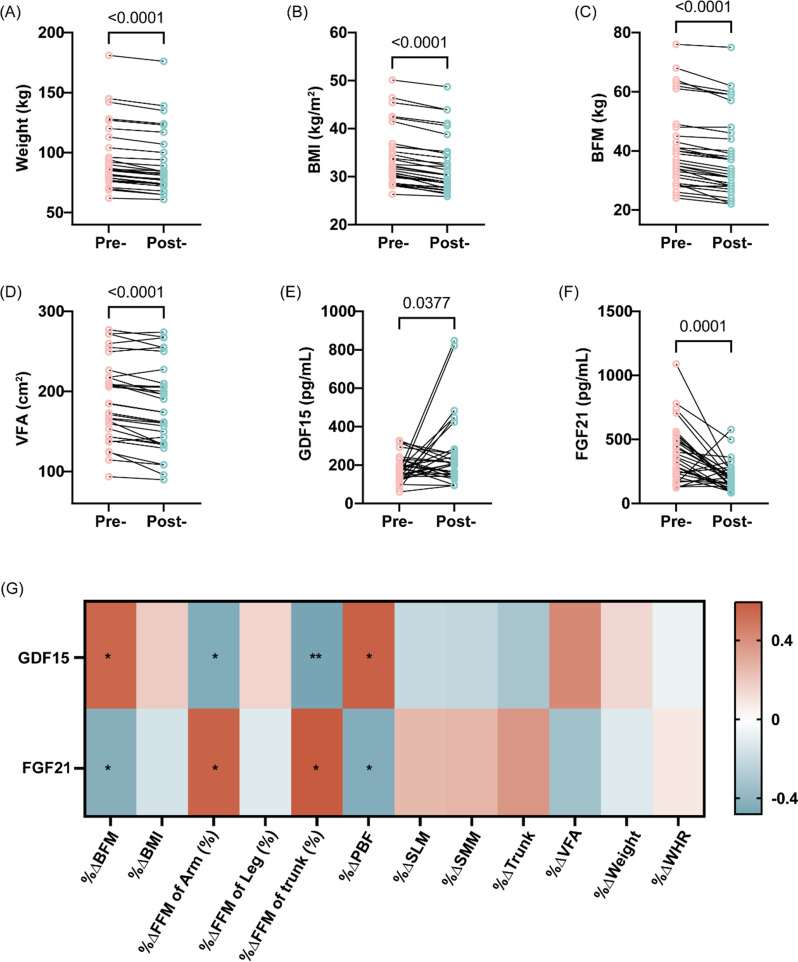 Figure 1
Figure 1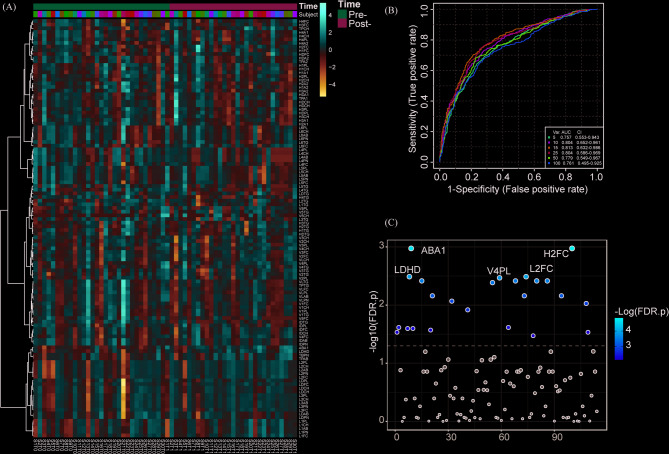 Figure 2
Figure 2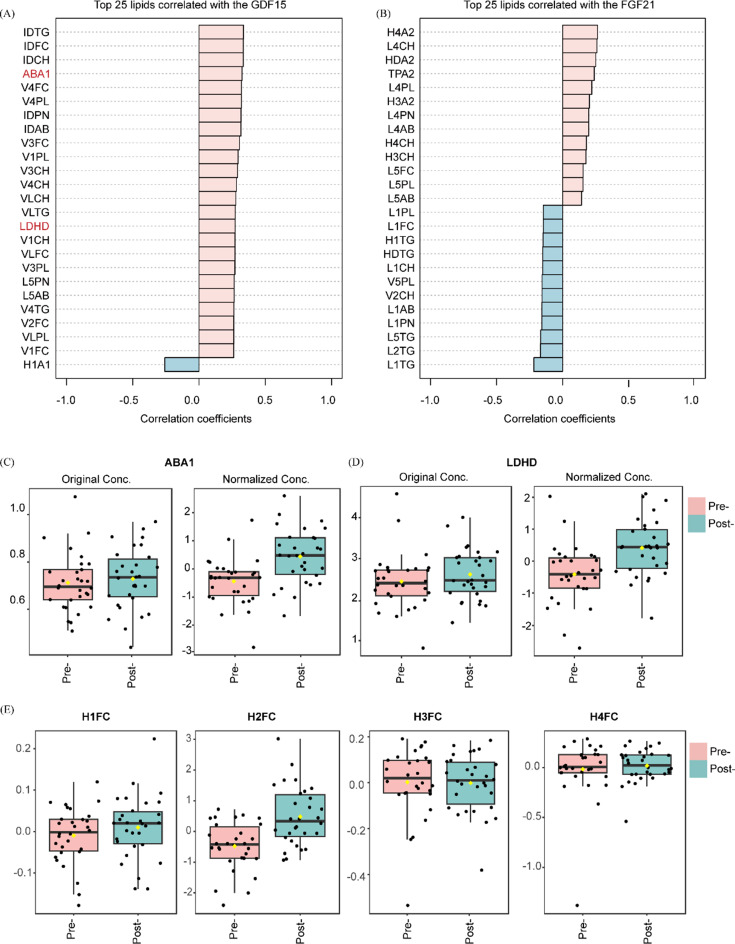 Figure 3
Figure 3- —http://dx.doi.org/10.13039/501100015401Key Research and Development Projects of Shaanxi Province
- —Xijing Medical Personnel Development Booster Initiative
- —http://dx.doi.org/10.13039/501100007390Instituto Nacional de Ciência e Tecnologia Centro de Estudos das Adaptações da Biota Aquática da Amazônia
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDiet and metabolism studies · Fibroblast Growth Factor Research · GDF15 and Related Biomarkers
Introduction
Obesity has become a major global public health challenge. According to the World Obesity Atlas 2025, the number of obese adults worldwide is projected to surge from 524 million in 2010 to 1.13 billion by 2030, an increase of over 115%. In China, the number of overweight/obese adults is expected to reach 515 million [1]. Obesity not only impacts physical appearance but is also strongly associated with diabetes, hypertension, cardiovascular diseases, and other health conditions [2]. Approximately, 1.6 million premature deaths annually are attributed to obesity, with 55% of type 2 diabetes-related premature deaths linked to obesity [3]. Therefore, it is imperative to explore weight management strategies that demonstrate efficacy, feasibility, and optimal patient adherence.
The classic ketogenic diet (KD), as a low-carbohydrate and high-fat dietary regimen, has been demonstrated by numerous studies to be effective for weight reduction and improvement of metabolic parameters [4–6]. Although, KD has demonstrated therapeutic potential for obesity, it has not been endorsed as a first-line treatment in clinical guidelines. The primary limitations include: poor long-term adherence in Chinese populations accustomed to traditional carbohydrate-rich diets (providing over 60% of energy intake), and significant reduction in cardioprotective foods such as whole grains, legumes, and dark green leafy vegetables. To address these critical clinical challenges, our study has innovatively developed a Modified Ketogenic Diet (MKD) better adapted to Chinese dietary patterns. This protocol features an optimized macronutrient distribution: 20% carbohydrates, 40% protein, and 40% fat, ensuring concurrent ketogenic efficacy and cultural compatibility.
Growth differentiation factor 15 (GDF15), a unique member of the transforming growth factor-β (TGF-β) superfamily, exhibits distinctive biological characteristics. As a pleiotropic cytokine, GDF15 plays a crucial role in stress responses following cellular injury. Its most prominent biological function involves regulating energy metabolism by suppressing appetite and reducing food intake, earning it the scientific designation of “anorexigenic hormone” [7, 8]. Recently studies have demonstrated that the signaling pathway formed by GDF15 and its specific receptor GDNF-family receptor α-like (GFRAL) plays a pivotal role in various metabolic disorders and pathological processes [9]. Currently, this signaling system has been established as an important biomarker for multiple diseases including obesity, type 2 diabetes mellitus, malignant tumors, and metabolic-associated fatty liver disease (MAFLD), providing new research directions for the diagnosis and treatment of related conditions [10, 11].
Fibroblast growth factor 21 (FGF21), a key member of the FGF19 subfamily, is predominantly expressed in liver tissue. It exerts its metabolic regulatory functions by targeting multiple organs including adipose tissue, liver, and pancreas to modulate glucose and lipid homeostasis [12, 13]. Research has demonstrated that FGF21 possesses multifaceted metabolic effects, including body weight reduction, improvement of insulin sensitivity, and reversal of hepatic steatosis, highlighting its significant physiological roles [14, 15].
Current evidence indicates that the anti-obesity effects of the classic KD are primarily mediated through two distinct pathways: GDF15-induced appetite suppression (reducing energy intake) and FGF21-enhanced energy expenditure (increasing metabolic activity) [16]. To investigate whether the MKD exerts its weight-loss effects, we conducted a 2-week MKD intervention in obese subjects, aiming to systematically evaluate its efficacy for weight reduction and identify potential circulating biomarkers related to its metabolic effects.
Materials and methods
Study participants
Between September 2023 and March 2024, we prospectively enrolled 35 metabolically healthy obese individuals (BMI ≥ 28 kg/m^2^) from the Endocrinology and Metabolism Clinic at the Xijing Hospital of The Fourth Military Medical University. All eligible participants underwent standardized baseline health assessments prior to study participation. Exclusion Criteria: (a) Secondary obesity (Cushing’s syndrome, hypothyroidism, growth hormone-secreting tumors, etc.); (b)Current use of obesity-inducing medications (e.g., corticosteroids, psychotropic drugs); (c)Conditions including pernicious anemia, malignancies, acute/chronic infections, severe hepatic/renal dysfunction, severe cardiopulmonary diseases, or cerebrovascular disorders; (d) Pregnancy or lactation status; (e) Diagnosis of type 2 diabetes mellitus; (f) Current use of antidiabetic drugs or lipid-lowering medications. During the study period, three participants withdrew and two others had insufficient key parameter data. Consequently, the final analysis included complete datasets from 30 participants.
The study protocol received ethical approval from the Medical Ethics Committee of the Xijing Hospital of Fourth Military Medical University (Approval No. KY20232166) and was prospectively registered with the Chinese Clinical Trial Registry (Registration No. ChiCTR 2300071823). All participants provided written informed consent. The investigation was conducted in full accordance with the ethical principles outlined in the Declaration of Helsinki.
Study protocol
One week prior to the study initiation, all participants were instructed to maintain their habitual dietary intake. After enrollment, participants received individualized weight-loss interventions and were required to collect first-void morning urine samples daily for urinary ketone assessment. Fasting serum samples and anthropometric measurements were obtained at two time points: baseline (pre-intervention) and 2-week follow-up (post-intervention). All serum samples were immediately aliquoted and stored at -80 °C until further analysis.
Diet interventions
This self-controlled, paired study implemented a comprehensive 2-week weight management program incorporating dietary, exercise, and psychological interventions. Participants received personalized KD plans (20% carbs, 40% protein, 40% fat) with daily caloric intake calculated as 70% of actual weight (kg) × (20–25 kcal). Prior to the intervention, each participant received a personalized meal plan from clinical nutritionists. Adherence was supported through daily remote monitoring via WeChat, which included morning urine ketone self-testing (values of +- to + + confirmed ketosis), submission of daily photo logs for meals and hydration, and provision of real-time feedback and psychological support to ensure protocol compliance. To minimize variations in energy expenditure, all participants were instructed to maintain their habitual levels of physical activity for the duration of the study. The primary endpoints included changes in body weight, GDF15, FGF21 levels, and serum metabolomic profiles after the 2-week intervention period.
Data collection
Body composition analysis
Body composition assessment was conducted using the InBody 770 bioelectrical impedance analyzer (Biospace Inc., Seoul, Korea) according to manufacturer’s protocols. Prior to assessment, participants were required to remove outer clothing and metallic items (e.g., watches and necklaces), stand barefoot on the footplate electrodes, and grasp the hand electrodes with both hands. The recorded parameters included height, weight, BMI, waist-to-hip ratio, fat mass, skeletal muscle mass, and visceral fat area, which were collected at both baseline and post-intervention.
Serum biomarker analysis
Serum biomarkers were quantified using ELISA assays: GDF15 was measured with Human GDF15 Quantikine ELISA Kit (R&D Systems, USA; Cat# DY957; range: 31.2–2000 pg/mL; sensitivity: <10 pg/mL), and FGF21 with Human FGF21 Quantikine ELISA Kit (Cat# DF2100; range: 15.6–1000 pg/mL; sensitivity: <5 pg/mL). All samples were analyzed in duplicate (inter-assay CV < 12%) with kit standards and internal controls. Plate readers were calibrated per manufacturer’s protocols, and out-of-range samples were re-tested after appropriate dilution. Paired blood samples were collected at baseline and post-intervention for biomarker analysis.
Serum lipoprotein subclasses analysis
Serum lipoprotein subclasses analysis was conducted using the Bruker AVANCE IVDr NMR metabolomics profiling system for comprehensive ¹H-NMR-based metabolic profiling. All procedures were performed in strict accordance with the IVDr Standard Operating Procedures (SOP) to quantify lipoprotein subclasses. Nuclear magnetic resonance (NMR) lipoprotein subfraction quantification includes subtypes of very low-density lipoprotein cholesterol (VLDL-c), low-density lipoprotein cholesterol (LDL-c), intermediate-density lipoprotein cholesterol (IDL-c) and high-density lipoprotein cholesterol (HDL-c), totaling 114 lipoprotein particles. Paired blood samples were collected at baseline and post-intervention for metabolomics analysis.
Statistical analysis
All statistical analyses were performed using SPSS software (version 26.0; IBM Corporation, Armonk, NY, USA) and MetaboAnalyst platform (version 6.0; McGill University, Montreal, QC, Canada; https://www.metaboanalyst.ca, accessed on 12 May 2025). Paired t-test analysis was performed to assess changes in body composition parameters pre- and post-MKD intervention. Non-normally distributed variables underwent logarithmic transformation. Candidate biomarkers were identified through comprehensive bioinformatics analysis. Receiver operating characteristic (ROC) curves were constructed using orthogonal partial least squares-discriminant analysis (OPLS-DA) methodology for comprehensive evaluation and screening of potential biomarkers. The OPLS-DA score chart was used to find different lipoprotein subclasses through the obtained VIP values. We used OPLS-DA to calculate the VIP value, which was analyzed with VIP > 1 and a t-test (P < 0.05), to screen the differential lipoprotein subclasses between groups. Furthermore, we evaluated the correlation between the concentration of GDF15, FGF21 and body composition, as well as lipid subclasses.
Results
Effects of a 2-week MKD on body composition in obese adults
To assess the impact of a 2-week MKD on body composition, all participants adhered to the diet and achieved ketosis, as quantitatively verified via ¹H-NMR metabolomics (Table 1). As shown in Table 1; Fig. 1, participants exhibited statistically significant reductions in both body weight (96.14 ± 27.23 kg vs. 91.63 ± 26.47 kg; Δ4.8%, P < 0.001) and BMI (33.99 ± 6.08 kg/m^2^ vs. 32.41 ± 5.95 kg/m^2^; Δ4.7%, P < 0.001) following the 2-week MKD intervention. Notably, patients exhibited significant reductions in body fat parameters, with BFM and VFA decreasing > 5% and PBF declining by 2.71%. Conversely,, lean mass indices (SMM, SLM, FFM) remained stable (change < 3%), indicating that the MKD promoted fat loss while effectively preserving lean mass.
Table 1. Effects of a 2-Week MKD on body composition and ketone body in obese adultsPre-Post-Change (%)t P Age (years)31.00 ± 8.56Sex (Female%)22 (71%)Weight (kg)96.14 ± 27.2391.63 ± 26.474.7612.743< 0.001BMI (kg/m^2^)33.99 ± 6.0832.41 ± 5.954.7113.681< 0.001WHR0.97 ± 0.080.97 ± 0.080.01-0.2450.808BFM (kg)41.10 ± 13.8938.28 ± 13.737.329.020< 0.001VFA (cm^2^)188.00 ± 50.45178.97 ± 53.635.455.236< 0.001PBF (%)42.40 ± 5.4241.29 ± 5.702.714.065< 0.001SLM (kg)51.85 ± 14.3850.25 ± 13.692.955.826< 0.001SMM (kg)30.57 ± 9.0429.64 ± 8.712.975.477< 0.001FFM (kg)55.04 ± 15.2153.35 ± 14.492.935.856< 0.001FFM of trunk (%)101.95 ± 5.67101.21 ± 5.580.701.9270.064FFM of Arm (%)17.25 ± 4.6616.66 ± 4.391.696.266< 0.001FFM of Leg (%)201.83 ± 10.87198.93 ± 9.701.383.1670.005Acetoacetate0.11 ± 0.050.12 ± 0.058.520.8250.416β-hydroxybutyrate0.01 ± 0.040.04 ± 0.08393.512.0600.048Acetone0.06 ± 0.110.16 ± 0.20164.502.1420.041Abbreviation: BFM: Body Fat Mass; BMI: Body Mass Index; FFM: Fat Free Mass; PBF: Percent Body Fat; SLM: Soft Lean Mass; SMM: Skeletal Muscle Mass; VFA: Visceral Fat Area; WHR: Waist-Hip Ratio
Fig. 1. Effects of a 2-week MKD on body composition and serum biomarkers in obese adults. (A-F) Changes in Weight (A), BMI (B), BFM (C), VFA (D), GDF15 (E) and FGF21 (F) after 2-week MKD. (G) Correlation matrix of GDF15/FGF21 with body composition (blue/red: positive/negative correlation). Abbreviation: BFM: Body Fat Mass; BMI: Body Mass Index; VFA: Visceral Fat Area. ^^P < 0.05, ^^P < 0.01, ^^P < 0.001
Effects of a 2-week MKD on serum GDF15 and FGF21 concentrations in obese adults
We next examined whether MKD modulates key serum biomarkers in obesity. As shown in Fig. 1, serum biomarker GDF15 levels increased significantly by 5.76% (P = 0.0377), whereas FGF21 levels decreased markedly by 51.91% (P = 0.0001) after the 2-week MKD intervention (Fig. 1E-F). Using body composition change-to-baseline ratios, we further assessed their correlations with baseline levels and intervention-induced changes in GDF15 and FGF21. Notably, baseline circulating GDF15 levels showed significant positive correlations with reductions in body fat indices (%ΔBFM, %ΔPBF; r = 0.43 to 0.45, P < 0.05), while demonstrating inverse associations with lean mass preservation (%ΔArm, %ΔTrunk; r=-0.48 to -0.45, P < 0.05). Conversely, baseline FGF21 concentrations exhibited negative correlations with body fat reduction (%ΔBFM, %ΔPBF; r=-0.44 to -0.43, P < 0.05) but positive correlations with lean mass changes (%ΔArm, %ΔTrunk; r = 0.44 to 0.47, P < 0.05). These distinct correlation patterns suggest that the magnitude of weight loss during MKD intervention is positively associated with baseline GDF15 levels and negatively associated with FGF21 levels (Fig. 1G).
Effects of a 2-week MKD Intervention on lipoprotein subclass profiles
To characterize the diet-induced lipoprotein remodeling, we analyzed subclass abundances pre- and post-intervention (Fig. 2A). ROC analysis (AUC = 0.779, Fig. 2B) indicated significant modifications in plasma lipoprotein profiles following the 2-week intervention. Principal component analysis (PCA) revealed distinct clustering between pre- and post-intervention states (Fig. 2C). Notably, 22 lipoprotein subclasses exhibited significant alterations post-MKD (VIP > 1, P < 0.05), with the top 15 most substantially changed lipoprotein subclasses highlighted in Fig. 2C. These data demonstrate that short-term MKD induces substantial lipoprotein redistribution.
Fig. 2. Changes in lipoprotein subclasses following a 2-week MKD intervention. (A) Heatmap displaying temporal changes in lipoprotein subclasses during the 2-week MKD intervention (pre- and post-intervention). (B) Receiver operating characteristic (ROC) curve analysis (AUC values shown) of plasma lipoprotein subclass profiles comparing pre- and post-MKD intervention states. (C) Orthogonal partial least squares-discriminant analysis (orthogonal partial least squares discriminant analysis, OPLS-DA) of lipoprotein fractions and subfractions reveals distinct clustering between pre- and post-MKD intervention groups
Association between circulating GDF15/FGF21 levels and lipoprotein subclass profiles in obese adults
Finally, we investigated whether GDF15 and FGF21 levels correlate with specific lipoprotein changes. Figure 3A and B demonstrate the associations between GDF15/FGF21 and lipoprotein subclass after adjustment for sex, age, and time. The ApoB/ApoA1 (ABA1) ratio (t = 5.381, P < 0.001) and LDL-c/HDL-c(LDHD) ratio (t = 5.095, P < 0.001) significantly increased during the MKD intervention (Fig. 3C and D). Notably, we observed that larger HDL subfractions (H1FC/H2FC) exhibited an increasing trend following treatment, whereas smaller HDL subfractions (H3FC/H4FC) displayed a decreasing trend. The most pronounced change was the elevation in H2FC levels (t = 6.119, P < 0.001) after MKD intervention (Fig. 3E). These findings imply that MKD induces a distinct shift toward larger HDL particles, which correlates with composition of MKD and may reflect altered lipid metabolism.
Fig. 3. Associations between circulating GDF15/FGF21 levels and lipoprotein subclasses. (A-B) Correlations of GDF15 and FGF21 with lipoprotein subclasses. (C-D) Dynamic changes in lipoprotein ratios: ApoB/ApoA1 (ABA1) and LDL-c/HDL-c (LDHD). (E) Changes in HDL-c subfractions (H1FC, H2FC, H3FC, H4FC) following MKD intervention
Discussion
In the present study, we investigated the effects of short-term MKD intervention on both weight loss and circulating GDF15/FGF21 levels in obese adults. Our results demonstrated that the 2-week MKD intervention induced statistically significant reductions in body weight and BMI, accompanied by an increase in GDF15 and a decrease in FGF21 levels. Further analysis revealed that these changes in biomarkers were closely associated with lipoprotein subclass remodeling, both the ABA1 and LDHD ratios showed significant increases, and HDL-c subclasses exhibited a marked shift toward larger particle sizes (increased H1FC/H2FC and decreased H3FC/H4FC), with the H2FC subclass demonstrating the most prominent elevation.
The KD is a metabolic intervention strategy characterized by low carbohydrate, high fat, and moderate protein intake. Its core mechanism involves shifting the body’s energy metabolism from glucose utilization to fat mobilization and ketone body production. The classical KD employs a strict 4:1 ratio of fat to (carbohydrates + protein), with 90% of calories derived from fat and only 2–3% from carbohydrates [17]. While this extreme ratio has demonstrated significant therapeutic effects in managing metabolic disorders such as obesity, type 2 diabetes, metabolic-associated fatty liver disease, and polycystic ovary syndrome (PCOS), its clinical application faces multiple challenges: the ultra-high fat content results in poor palatability, frequent gastrointestinal discomfort, restriction of cardioprotective foods like whole grains and legumes, and particularly poor long-term adherence among Chinese populations (traditional carbohydrate intake accounts for >60% of energy) [18–21]. To address these limitations, this study innovatively developed a MKD that optimizes macronutrient ratios to 20% carbohydrates, 40% protein, and 40% fat. Clinical research demonstrates that MKD not only preserves the metabolic benefits of traditional KD but also significantly improves its clinical application drawbacks. Just two weeks of MKD intervention achieved a 4.8% weight reduction and 4.7% BMI decrease, with body fat mass and visceral fat area significantly reduced by >5%, while effectively maintaining lean body mass stability (change < 3%). This modified approach not only enhances dietary variety and patient tolerance but also provides a more sustainable metabolic management solution for populations accustomed to high-carbohydrate diets, such as the Chinese population.
The discovery of the GDF15-GFRAL axis in 2017 opened new therapeutic avenues for obesity treatment [22]. This signaling pathway not only suppresses food intake through central mechanisms but also exerts peripheral anti-obesity effects by promoting thermogenesis, delaying gastrointestinal emptying, enhancing lipolysis, and maintaining skeletal muscle energy expenditure [23–26]. A landmark study by Gregory R. Steinberg’s team at McMaster University, published in Nature, demonstrated that the GDF15-GFRAL pathway increases skeletal muscle calcium futile cycling to simultaneously suppress appetite and maintain energy expenditure during caloric restriction, providing a novel solution to overcome the challenge of “weight rebound” in obesity management [27]. Notably, the anti-obesity effects of metformin, a first-line antidiabetic drug, have been shown to be mediated through GDF15. Accumulating evidence indicates that metformin upregulates GDF15 expression in various tissues (including intestine and kidney), which then acts on the central nervous system to suppress appetite and regulate energy metabolism [28–30]. Furthermore, the positive feedback loop between GDF15 and AMPK is essential for complete metformin activation, establishing GDF15’s central role in metformin’s metabolic regulatory network [31]. Complementary studies by Wu JW’s team revealed that KD can activate hepatic PPARγ signaling to upregulate GDF15 transcription, thereby promoting weight loss [17]. Our research further demonstrates that MKD intervention significantly elevates circulating GDF15 levels and produces marked weight reduction. Consistent with previous findings, our study not only confirm the critical role of GDF15 in weight regulation, but also provides a clinically feasible long-term dietary intervention strategy for obesity management.
FGF21, as a key metabolic regulatory hormone, demonstrates a specific response pattern to dietary nutrients in both humans and rodents. Substantial research evidence indicates that the expression and secretion of this hormone are precisely regulated by the carbohydrate-protein balance. Regarding nutrient-sensing mechanisms, carbohydrates have been identified as the most potent stimulators of FGF21.Clinical studies show that both acute glucose loading and chronic high-carbohydrate diets can significantly elevate circulating FGF21 concentrations [32–34]. This phenomenon has been further validated in rodent models, manifesting as synchronized increases in peripheral blood FGF21 levels and hepatic FGF21 mRNA expression [35]. In contrast to carbohydrates, dietary protein exerts bidirectional regulatory effects on FGF21: protein restriction significantly elevates FGF21 levels in both humans and animals [36, 37], while moderate protein supplementation effectively counteracts carbohydrate-induced FGF21 elevation [38]. This regulatory effect is most pronounced under high-carbohydrate/low-protein dietary conditions. In our study, MKD (low-carbohydrate/high-protein) intervention induced a rapid decline in plasma FGF21 levels within a short period, independent of energy intake and body weight changes, strongly confirming FGF21’s specific responsiveness to macronutrient composition. Particularly insightful is the observation that the characteristically elevated fasting FGF21 levels in obese individuals [26] were significantly reduced after MKD intervention. This result not only excludes the interference from metabolic diseases per se but also establishes FGF21’s molecular role as a “nutritional status sensor”. From a mechanistic perspective, FGF21’s rapid response to dietary changes likely stems from its unique regulatory properties: increased protein intake directly suppresses FGF21 expression, while reduced carbohydrate availability removes its primary stimulatory signal. Notably, our results reveal a crucial dissociation from classic KD patterns. While both classic KD and our MKD elevate GDF15, our modified diet produced a distinct reduction in FGF21 - contrasting sharply with the characteristic FGF21 elevation typically observed in classic KD studies. This divergent response suggests that the higher protein content (40%) in our MKD protocol may attenuate the typical ketogenic stimulation of FGF21 through hepatic PPARα activation and low-carbohydrate stress signaling. Our findings systematically elucidate the dynamic regulatory patterns of FGF21 by macronutrient composition, providing novel experimental evidence and theoretical foundations for understanding the intricate interplay between diet, hormones and metabolism.
Characterized by a high fat intake (70–80%), the classic KD reduces body weight while improving triglyceride levels, blood pressure, and blood glucose regulation. Nevertheless, it elevates total cholesterol and low-density lipoprotein cholesterol (LDL-c) levels, which are established independent risk factors for cardiovascular disease [39]. Our findings indicate that the MKD similarly affects lipoprotein metabolism in a complex manner. Consistent with the established effects of ketogenic diets, we observed increases in several atherogenic lipid parameters. More notably, our NMR-based profiling revealed a shift in HDL subclasses toward larger particles (H1FC/H2FC), particularly H2FC-a pattern that contrasts with some reports on classic KD and may represent a distinctive feature of our modified macronutrient composition. This shift stands in contrast to the findings of Montefusco et al., who reported that a carbohydrate-moderate hypocaloric diet reduced total HDL TG content, suggesting a decrease in larger, TG-rich HDL particles [40]. The shift toward larger HDL particles observed with the MKD appears to be attributable to its specific dietary composition rather than energy restriction alone, which is further supported by the rapid induction of ketosis and distinct alterations in lipoprotein profiles. The clinical implications of these simultaneous changes, including both elevated atherogenic lipids and a distinct shift in HDL subclasses, require further exploration. Future studies directly comparing the MKD with isocaloric classic KD will help clarify the diet-specific effects on lipoprotein metabolism and their potential long-term cardiovascular outcomes.
This study demonstrates three key strengths. Firstly, this study offers a comprehensive metabolic evaluation by innovatively assessing both weight-loss efficacy and serum biomarker responses (increased GDF15 and decreased FGF21 levels) during short-term MKD intervention, offering multidimensional insights into its metabolic regulatory mechanisms. Notably, MKD induces potentially beneficial remodeling of lipoprotein subclasses. Secondly, the findings possess significant clinical value. Our data showed that a mere 2-week MKD intervention achieved rapid and significant reductions in body weight/BMI, thereby establishing strong evidence for MKD as an effective short-term intervention for obesity. Of particular interest, the opposing regulatory trends observed in GDF15 (anorexigenic) and FGF21 (metabolic regulator) suggest that MKD may modulate energy homeostasis through distinct signaling pathways. Thirdly, this study adheres to stringent scientific methodology. Standardized biomarker detection methods (particularly NMR spectroscopy-based HDL-c subclass analysis) were employed to ensure reproducibility. Additionally, rigorous adjustment for confounders (age, sex, and time) significantly enhanced the validity of the conclusions.
This study has several limitations. Firstly, it is important to note that our models incorporated adjustments for age and gender. Nevertheless, the generalizability of the findings to broader populations is limited by a relatively small size, a higher proportion of female participants, and a relatively young age range. Secondly, the single-arm design without a control group (e.g., a classic KD group) prevents us from establishing definitive the superiority of the MKD. Our results should be interpreted as exploratory. Thirdly, the short-term MKD intervention precludes assessment of whether the observed metabolic changes, such as weight loss, elevated GDF15, reduced FGF21, and lipoprotein subclass remodeling, represent sustained adaptations. Furthermore, although participants were instructed to maintain their habitual physical activity levels and dietary was remotely monitored, the lack of rigorous control and objective measurement of energy expenditure may have introduced confounding. Finally, while we identified associations between dietary effects and biomarker changes, the lack of experimental validation (e.g., in vitro or animal models) prevents establishment of causal mechanisms. Future studies should incorporate larger cohorts, longer intervention periods, controlled dietary comparisons, and mechanistic investigations through multi-omics approaches or functional experiments to validate these preliminary findings.
Conclusion
This study demonstrates that a short-term MKD intervention significantly improves adiposity metrics in obese individuals and induces distinct alterations in lipoprotein subclass distribution, accompanied by elevated circulating GDF15 and reduced FGF21 levels. Notably, the observed shift in HDL subclasses occurred concurrently with elevations in atherogenic lipid parameters. Therefore, the potential metabolic significance of HDL remodeling must be interpreted with caution. Subsequent studies should focus on long-term outcomes and explore the molecular pathways associated with GDF15 and FGF21 in the regulation of lipoprotein metabolism.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Trusz GJ. Fibroblast growth factor 21. Differentiation. 2024 Sep-Oct;139:100793.10.1016/j.diff.2024.10079338991938 · doi ↗ · pubmed ↗
- 2Tillman EJ, Rolph T. FGF 21: an emerging therapeutic target for non-alcoholic steatohepatitis and related metabolic diseases. Front endocrinol (Lausanne). 2020 Dec 14;11:601290.10.3389/fendo.2020.601290 PMC 776799033381084 · doi ↗ · pubmed ↗
- 3Sigvardsen CM, Richter MM, Engelbeen S, Kleinert M, Richter EA. GDF 15 is still a mystery hormone. Trends Endocrinol Metab. Oct 29 2024:S 1043-2760(24)00254-6.10.1016/j.tem.2024.09.00239472228 · doi ↗ · pubmed ↗
- 4Montefusco L, D’Addio F, Loretelli C, Ben Nasr M, Garziano M, Rossi A, et al. Anti-inflammatory effects of diet and caloric restriction in metabolic syndrome. J Endocrinol Invest. Nov 2021;44(11):2407–15.10.1007/s 40618-021-01547-y PMC 850212133686615 · doi ↗ · pubmed ↗