Investigating the potential role of metabolic resistance genes in conferring cross-resistance to pyrethroids and polycyclic aromatic hydrocarbon pollutants in the major malaria vector Anopheles coluzzii
Abdullahi Muhammad, Sulaiman S. Ibrahim, Hanafy M. Ismail, Helen Irving, Mark J.I. Paine, Charles S. Wondji

TL;DR
This study shows that certain mosquito genes linked to insecticide resistance can also break down environmental pollutants, potentially increasing resistance in the wild.
Contribution
The study identifies specific P450 enzymes in malaria mosquitoes that metabolize PAHs, linking environmental pollutants to insecticide resistance.
Findings
Microsomes from resistant mosquitoes significantly depleted fluorene and fluoranthene.
CYP6P3 and CYP6Z3 showed high fluorene metabolism, while CYP6Z2 and CYP6Z3 metabolized fluoranthene.
No naphthalene metabolism was observed with the tested P450s.
Abstract
Polycyclic aromatic hydrocarbons (PAHs) are a class of ubiquitous and recalcitrant environmental pollutants generated from petroleum activities and/or biological conversion of organic materials. Environmental exposure of mosquitoes to these pollutants can potentially select resistance to insecticides used in public health for vector control. To understand the cross-resistance potentials between PAHs and pyrethroid insecticides, microsomal fractions prepared from Anopheles coluzzii mosquitoes obtained from agricultural sites and a laboratory susceptible strain, Ngousso, were tested with three major PAHs - fluorene, fluoranthene and naphthalene. Recombinant P450s previously associated with pyrethroid resistance in Anopheles gambiae (CYPs 6M2, 6Z2, 6Z3, 9J5, 6P3, 6P4, 6P5, CYP9K1) and Anopheles funestus CYP6P9a were also used to investigate metabolism of the above PAHs alongside the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
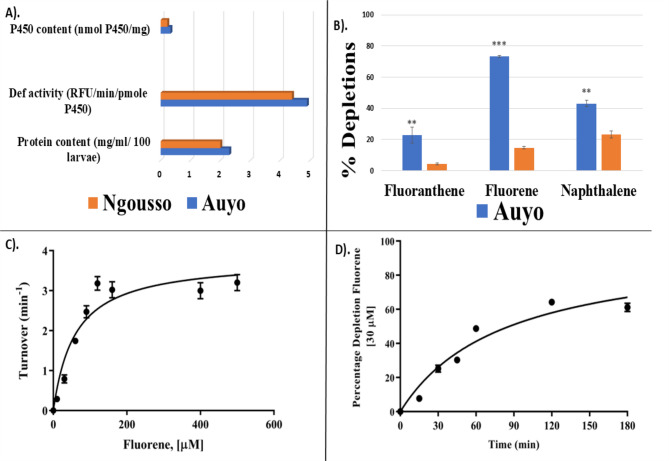 Figure 1
Figure 1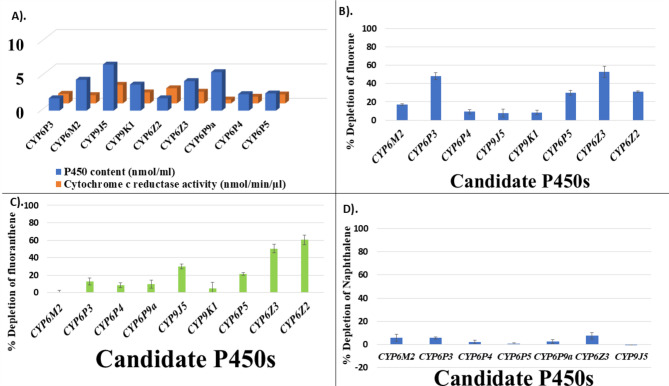 Figure 2
Figure 2- —https://doi.org/10.13039/501100009614Petroleum Technology Development Fund
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Insect Resistance and Genetics · Insect Pest Control Strategies
Background
Insecticide resistance poses a significant threat to the gains made in malaria control through vector control tools [1–3]. Insecticide-based interventions such as the insecticide-treated nets (ITNs) and indoor residual spraying (IRS) are being challenged by the evolution/escalation of resistance even to newer chemistries [4]. Resistance to at least one insecticide has been reported in virtually all the WHO African regions, with strong indications of cross-resistance to other insecticides in most areas [2, 5, 6]. An. coluzzii,* An. gambiae* and An. funestus are the major malaria vectors in Nigeria [7], with An. coluzzii becoming more predominant in all climates of West/Central Africa [8]. Resistance to all classes of insecticides has been reported in various studies with near fixed frequencies of kdr mutations in some cases [9]. An. coluzzii is highly adaptive to breeding in urban areas compared to other vectors. This may be attributed to its increased resistance to osmotic stress from anthropogenic pollutants like the PAHs [10]. This suggests its widespread exposure to urban pollution [11].
When a mosquito population becomes resistant to insecticides, the same mechanism can confer resistance to other insecticides [12]. Cross-resistance is most pronounced through metabolic resistance mechanisms due to the substrate specificity and/or promiscuity [13] of the enzymes involved in the sequestration, metabolism and excretion of xenobiotics, which can lead to the metabolism of a wide range of compounds [14, 15]. The key enzymes involved in insecticide metabolism and resistance are the cytochrome P450 monooxygenases (P450s), glutathione S-transferases (GSTs) and carboxylesterases [16].
On the other hand, prior exposure to environmental pollutants and/or agrichemicals has been shown to increase selection pressure on mosquito vectors, leading to elevated levels of metabolic resistance genes, which are linked to cross-resistance to vector control insecticides. Several studies have demonstrated that survival of mosquitoes in polluted breeding sites has led to an increase in resistance and urban malaria transmission [17–19]. Members of the Anopheles gambiae complex are often implicated in this adaptation as they breed in polluted waters [20–22]. Hence, it is crucial to understand the molecular mechanisms through which environmental pollutants contribute to the selection of insecticide resistance in malaria vectors, thereby unravelling the cross-resistance potentials of individual compounds [23–26]. Although many studies have documented a potential link between exposure to pollutants such as polycyclic aromatic hydrocarbons (PAHs) and insecticide resistance, little is known of the underlying molecular mechanisms driving cross-resistance in Anopheles mosquitoes.
Microsomes are subcellular fractions generated from the ultracentrifugation of homogenised tissues containing the membrane-bound enzymes, including the cytochrome P450 monooxygenases [27]. Microsomal fractions from higher organisms have been proven very useful in the studies of drug and other compounds metabolisms, as well as toxicity studies in drug discovery and related disciplines [28–30]. Thus, they can potentially be explored in the study of insecticide cross-resistance in malaria vectors [29]. Microsomes isolated from field-resistant populations of malaria vectors can be a useful tool in determining the potential cross-resistance of newer chemistries (e.g., synthetic insecticides) and other environmental pollutants such as PAHs [31–33].
Several studies have implicated overexpression and overactivity of key Anopheles cytochrome P450 monooxygenases in insecticide resistance [34]. Some of these P450s have been functionally validated in vitro using heterologous expression in E. coli [14, 35, 36] and in vivo, using transgenic expression in Drosophila flies [37]. For example, in An. funestus, for example, the duplicated CYP6P9a/b genes are the major drivers of pyrethroid resistance and demonstrate the ability to metabolize and confer resistance to non-pyrethroid insecticides [13, 38–40]. In addition, other P450s such as CYP9K1 [41] have been shown to metabolize a type II pyrethroid, deltamethrin, but not a structurally similar type I pyrethroid, permethrin. Others include the highly polymorphic CYP6M7, which was shown to metabolise pyrethroids [42] and CYP6AA1, which was shown to metabolise pyrethroids as well as the carbamate bendiocarb [43].
In An. gambiae, several P450s have also been implicated in the metabolism of pyrethroid insecticides, these include CYP6M2, a strong metaboliser of pyrethroids including permethrin and deltamethrin [44], CYP6P3, found to significantly metabolize types I and II pyrethroids [45], CYP6P5 located on the pyrethroid resistance locus with appreciable copy number variations [46] in the CYP6 cluster [47]. Other important pyrethroid-associated P450s include CYP6Z3 [48] and CYP6Z2, which are found to have a broad range of substrate specificity, suggesting potential roles in survival at the larval stage [49]. CYP9K1 [50] and CYP9J5 [51] are also involved in conferring resistance to pyrethroids.
Sixteen PAHs have been reported as being of environmental concern by the US Environmental Protection Agency due to their persistence and toxicity. The smaller PAHs, with fewer benzene rings and no functional groups substituting the hydrogens in their structures, tend to be more persistent in the environment due to their poor microbial bioremediation [52, 53]. These PAHs also have greater access to the cellular membrane of mosquito larvae in the breeding sites due to their higher hydrophobicity, an important requisite for easy access to the lipid bilayer membrane. The main aim of the current study is to investigate the cross-resistance potentials between this class of ubiquitous pollutants and public health insecticides using recombinant P450s and microsomes isolated from resistant and susceptible strains of An. coluzzii. The study informs on the role of these compounds as drivers of metabolic insecticide resistance in An. coluzzii. To do that, three lower molecular weight PAHs were selected for these experiments due to their characteristics. Naphthalene with two fused benzene rings, fluorene with two benzene rings fused to a five-membered ring and fluoranthene, consisting of a naphthalene moiety and a benzene ring connected by a five-membered ring.
To study the cross-resistance between PAHs and pyrethroids, a P450 panel comprising An. gambiae CYPs 6Z2,* 6Z3*,* 6P5*,* 9K1*,* 6M2*,* 9J5*,* 6P3 and 6P4*, and An. funestus CYP6P9a was investigated for metabolic activities toward the PAHs and pyrethroid insecticides.
Isolated microsomes prepared from the field population as well as from the fully susceptible An. coluzzii lab colony, Ngousso were used in metabolism assays to investigate cross-resistance between pyrethroid insecticides and the three PAHs. In addition, the above P450s were heterologously expressed in E. coli and investigated for their potential to confer cross-resistance to these two unrelated groups of compounds.
Methods
Mosquito collection and rearing
Blood-fed, indoor resting female Anopheles mosquitoes were caught using electric aspirators (John W. Hock, Florida, USA) in Auyo town, Auyo Local Government, Jigawa State, Nigeria in September 2019. This is a highly irrigated site whose details have been described earlier [9]. The F_0_ female mosquitoes were morphologically identified [54] as members of Anopheles gambiae complex and kept in standard insectary conditions of 25 °C and 75% relative humidity and 12:12 h light: dark cycles, and maintained on 10% sucrose solution until they were gravid. Gravid females were individually forced to lay eggs [55] and the eggs were transported to Liverpool School of Tropical Medicine for the downstream analysis. Using the genomic DNA extracted from the F_0_ individuals, species were confirmed using SINE 200 PCR [56]. Eggs were pooled into trays and allowed to hatch. The Auyo population is highly resistant to pyrethroids and therefore used for this study to understand the cross-resistance potentials between pyrethroids and PAHs. The laboratory susceptible Anopheles coluzzii colony, Ngousso [57], which is fully susceptible to all insecticides, was used for comparison to the field resistant populations.
Microsome preparation
The two strains of Anopheles coluzzii Auyo and Ngousso were reared in the insectary under standard insectary conditions to generate a substantial number of fourth instar larvae for the microsome preparations. Microsome preparation was conducted according to the method reported earlier with some modifications [58]. About 600 of 4th instar larvae were homogenised in 20 ml of ice-cold potassium phosphate buffer, pH 7.4, that has been supplemented with 1x protease inhibitor (Roche complete, ultra EDTA free, Sigma Aldrich, MA, USA). The larvae were homogenized in a 40 ml Dounce homogenizer equipped with a loose B pestle (Wheaton Science, Millville, NJ, USA). Filtration was conducted with layers of nylon filters to remove the residual debris. The filtrate was initially centrifuged at 10,000 xg for 10 min at 4 ˚C to separate the cytosolic fractions (supernatant) from other heavier cellular components. The supernatant was then centrifuged at 200,000 xg for 45 min at 4 ˚C to collect the pellet and discard the new supernatant. The pellet was reconstituted in 0.1 M potassium phosphate buffer, pH 7.4, containing 20% glycerol. Total protein content of the microsomes was determined using the Bradford method [59]. Cytochrome P450 content (Cary WinUV Software, Agilent Technologies) and microsomal P450 activity were also determined using spectral activity assay [60, 61].
Heterologous expression of recombinant P450s
The recombinant P450s used in this study were acquired from the enzyme characterization group (ECG) leader, Dr. Mark J.I. Paine of the Vector Biology Department, Liverpool School of Tropical Medicine. All the P450s were expressed from field-resistant populations of An. gambiae except for CYP6P9a, which was expressed from An. funestus. The heterologous expression of the P450s was conducted according to the previously described approaches [35, 44, 49]. Briefly, P450s were expressed by using ompA and pelB signal sequences to direct P450 and An. gambiae CPR (AgCPR), respectively, to the inner membrane of E. coli a functional monooxygenase. The P450 sequences were amplified from cDNA and fused to the ompA signal peptides in simple PCR reactions. This is followed by ligation of the digested fragments into linearized expression vector pCWori+, thereby creating a construct pB13:ompA + 2-P450. Competent JM109 cells were co-transformed with plasmids pB13:ompA + 2-P450 and pACYC-AgCPR for the expression of functional P450 and An. gambiae CPR, respectively. Colonies carrying the two plasmids as confirmed by the colony PCR are used for a 12–14 h starter culture overnight. Typically, 0.2 L cultures were supplemented with 2 mL from the starter cultures and incubated at 37 °C and 200 rpm shaking while the absorbance at 600 nm was being monitored. When the OD reached 0.6–0.8, the cultures were cooled down to 25 °C for 30 min with shaking at 150 rpm. Induction with 1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) and 0.5 mM 5-aminolaevulinic acid (ALA) was conducted. Determinations of the P450 content and CPR activity were conducted using the procedure of [61] and [60], respectively.
Measurement of microsomal P450 activity using model probe substrate
P450 activity was determined in the prepared microsomes from both Auyo and Ngousso strains using diethoxyfluorescein (DEF) as the model fluorogenic substrate using the methods earlier described [15]. Briefly, the enzyme-buffer mix consisted of 0.05 µM of cytochrome P450 (microsomes), 50 mM potassium phosphate buffer, pH 7.4, 0.5 µM cytochrome b5 and 10 µM DEF. To initiate the reactions in a 96-well plates (Thermo Labsystems, Basingstoke, UK), a NADPH (nicotinamide adenine dinucleotide) regeneration mix (consisting of 1mM glucose-6-phosphate, 0.1 M NADP+, 0.25mM magnesium chloride, IU/ml glucose-6-phosphate dehydrogenase and 50 mM potassium phosphate buffer pH 7.4) was added in the positive replicates whereas the same buffer with no NADPH was added in the negative wells. Using the excitation (482 nm) and emission (520 nm) wavelengths of DEF, the absorbance was read for 15 min, and the relative fluorescence unit per min per picomole of the P450 (RFU/min/pmol of P450) was determined.
In vitro metabolism assay of PAHs with heterologously expressed P450s
To compare the metabolic activity of the microsomes and the recombinant cytochrome P450s towards PAH metabolism assays were conducted side by side as described earlier [38, 43]. The NADPH regeneration mixture (consisting of 0.1 M NADP+, 50mM potassium phosphate buffer, pH 7.4, 1 mM glucose-6-phosphate, 0.25 mM magnesium chloride and 1U/ml glucose-6-phosphate dehydrogenase) was used in the positive replicates. For the negatives, the same buffer with no added NADP + was used. Enzyme-buffer mix comprised of 0.05 µM recombinant P450 or microsome, 0.4 µM cytochrome b5, 50 mM potassium phosphate buffer pH 7.4 and 20 µM of the PAH substrates. To initiate the reactions, regeneration mixture was added to the enzyme-buffer mix in a 1:1 ratio (total of 200 µl) following activation for 5 min at 30 °C and 1200 rpm shaking. The reaction was run for 2 h at 30 °C with 200 rpm shaking. To stop the reaction, 200 µl of ice-cold High Performance Liquid Chromatography (HPLC) grade acetonitrile was added, followed by additional shaking for 5 min to precipitate the proteins. The mixture was kept on ice for 10 min before centrifuging at 16,000xg for 20 min. The supernatants were filtered through 0.45 μm PTFE filters (ThermoFisher Scientific, MA, USA) and filtrate transferred to HPLC vials for onward analysis.
Using the Agilent HPLC 1200 infinity series, analysis of the PAHs metabolism was conducted. The HPLC conditions consisted of mobile phases of HPLC grade acetonitrile and water in the ratio of 80/20 and detected at a wavelength of 254 nm. 50 µl of the supernatant was injected on the HPLC and peaks were separated on a 250 × 4.6 mm (5 μm) Supelcosil LC-18-DB column (Supelco, Sigma-Aldrich, Gillingham, U.K.) in a 20 min run. Percentage depletions for each compound were calculated in both the microsomal and recombinant P450 studies. Using pair-wise t-test, comparisons were made between the positives (+ NADPH) (where reactions were expected) and the negative controls (-NADPH) (where no reactions were expected due to the absence of NADPH source in the mix). The effect size between the two means of each comparable group was calculated [62]. Comparisons were also made using the same parameters between depletions in field resistant and laboratory susceptible microsomal metabolism of PAHs.
Fluorene turnover assay using microsomes
Because of the higher percentage of depletion observed in the Auyo microsomal metabolism of fluorene, it was chosen to be analysed further for its turnover and kinetic studies. To understand the turnover rates of the fluorene depletions, reaction run time was varied between 15 and 150 min. Specific points used were 15, 30, 45, 60, 90, 120 and 150 min while the fluorene concentration was maintained at 30 µM in all these time points.
Determination of steady-state kinetics parameters for the microsomal metabolism of fluorene
For the determination of steady-state kinetic parameters, assays were conducted for 30 min with 0.05 µM microsomal P450 while varying the concentrations of fluorene (0–600 µM). Kinetics plot of velocity against the substrate concentrations was made using the least squares non-linear regression in GraphPad Prism 6.03 Software (GraphPad Inc., La Jolla, CA, USA) that fits into the canonical Michaelis-Menten model as previously described for pyrethroid insecticides [43].
Results
P450 content and activity in microsomal fractions
Microsomal preparations from both the field-resistant strain (Auyo) and laboratory susceptible strain (Ngousso) of An. coluzzii produced P450 contents of 0.33 nmol P450/mg and 0.226 nmol P450/mg m, respectively. The microsomes were screened for P450 activities using the model fluorogenic substrate diethoxyfluorescein (DEF). DEF activities of 4.9 RFU/min/pmole P450 and 4.4 RFU/min/pmole P450, respectively (Fig. 1A), were established for the Auyo and Ngousso microsomes, confirming the presence of functional cytochrome P450s. A total protein content of 2.0 mg/ml/100 larvae and 2.3 mg/ml/100 larvae were also determined for the Auyo and Ngousso microsomal preparations, respectively (Fig. 1A).
Metabolism of the PAHs by microsomal fractions
Microsomal P450 demonstrated metabolic activity towards PAHs (fluorene, fluoranthene and naphthalene) with higher percentage depletions observed in the Auyo microsomes. For example, significant depletion of fluorene (73.3 ± 0.44%, P = 0.0001, r = 0.99) was seen with the Auyo microsome compared to the Ngousso microsome (Fig. 1B). A Similar profile was obtained with fluoranthene, with 22.8 ± 5.2% significantly depleted by Auyo microsome (P = 0.001, r = 0.97), compared with the Ngousso microsome. Similar pattern was observed with naphthalene.
Steady-state kinetic parameters were investigated with fluorene, the most significantly depleted PAH by the Auyo microsomes. Turnover of 37.02 min^−1^ ± 3.67 was recorded. The microsome demonstrated moderate affinity towards fluorene (Fig. 1C), with a Km value of 58.69 µΜ ± 20.47. Catalytic rate of (K_cat_) of 4.196 min^−1^ ± 0.436 with a corresponding high catalytic efficiency of 0.0715 ± 0.026 min^−1^ µM^−1^ (Fig. 1D).
Fig. 1. Catalytic activity of the Auyo and Ngousso microsomes towards PAHs and the enzyme kinetics studies of the Auyo microsomes with fluorene as substrate. A Def activity and the protein content of the isolated microsomes. B Percentage depletions of the metabolism of the select PAHs by the microsomes isolated from the pyrethroid-resistant and susceptible strains of Anopheles coluzzii. Results are presented as mean ± SD of three replicates of the positives (+ NADPH) compared to the negatives (-NADPH). ** Significant difference between the Ngousso and Auyo percentage depletions for each substrate. C Michaelis-Menten plot of fluorene metabolism by microsomes isolated from the field-resistant strain (Auyo). D Time course of fluorene turnover, substrate concentration 20 µM
The metabolic activity of recombinant pyrethroid-associated P450s towards PAHs
To understand the potential cross-resistance liabilities between the recalcitrant environmental pollutant PAHs and pyrethroids, cytochrome P450s previously implicated in pyrethroid resistance/metabolism were recombinantly expressed and used for metabolism assays with PAHs. The pyrethroid metabolism associated P450s showed varied levels of metabolism towards the three select PAHs, naphthalene (Fig. 2D), fluorene (Fig. 2B) and fluoranthene (Fig. 2C). For example, none of the P450s used in this study depleted up to 10% of naphthalene, suggesting no cross-resistance towards this PAH and pyrethroid insecticides through the P450 metabolism route (Fig. 2D). On the other hand, fluoranthene was significantly depleted by the recombinant CYP6Z2 and CYP6Z3, with percentage depletions of 60% ± 4.9 and 50.4% ± 5.3, respectively (Fig. 2C). In the case of fluorene, highest depletions were seen with the recombinant CYP6Z3 (52.8 ± 0.8%) and CYP6P3 (47.9 ± 2.3%) (Fig. 2B), with much lower depletions obtained from CYP9K1 (8.5% ± 2.5), CYP6P4 (9.3% ± 4.3), and CYP9J5 (7.7% ± 2.5).
Fig. 2. Metabolism of PAHs by pyrethroid-associated recombinant cytochrome P450 monooxygenases. Details of the P450 membranes used in the study including the P450 content and CPR activity (A). Percentage depletions of fluorene (B), fluoranthene (C) and naphthalene (D) for the various recombinant cytochrome P450s. Values are presented as mean ± S.D. of three technical replicates. Percentage depletions were calculated by comparison of positive (+ NADPH) vs. negative (-NADPH) reactions in triplicate
Discussion
Chemical insecticides used in vector control tools and agriculture are considered the major drivers of insecticide resistance alongside environmental pollutants [51, 63–65]. Microsomal fractions isolated from pyrethroid-resistant and susceptible strains of An. coluzzii were used to study the cross-resistance between pyrethroids and PAHs as a class of ubiquitous environmental pollutants. Microsomes are the cytoplasmic fractions of the cells containing the membrane-bound enzymes, including the cytochrome P450s, and their use in drug discovery and toxicity studies has been well documented [28, 30] compared to vector biology studies. They can thus be explored for the studies of insecticide resistance liabilities without the need for recombinant expression of individual P450s. Successful isolations of microsomes from different insect species have been reported, mainly from Aedes aegypti mosquitoes [32, 33, 66–68]. In the present study, microsomes with substantial P450 activity and content were successfully isolated from the larvae of pyrethroid-resistant and susceptible strains of An. coluzzii. The P450 yield and activity were higher (0.33 nmol P450/mg protein for the resistant and 0.226 nmol P450/mg for the susceptible) than those seen in a recent study [66] but lower than what was obtained from southern armyworm (Spodeptera eridania) microsomal preparations [69].
Generally, metabolism of PAHs by cytochrome P450s in different organisms, including insects, proceeds via C-H activation [70, 71]. For example, the human CYP3A4 has been shown using HPLC coupled with NMR to generate a mono-hydroxylated product, fluorenol and a 9-oxo substituted product, fluorenone, from the fluorene parent compound [70]. The microsomal fractions from pyrethroid-resistant strains metabolised all three PAHs significantly more than the pyrethroid susceptible ones, suggesting there was pyrethroid cross-resistance potential to environmental pollutants. In a related study [33] in Aedes aegypti, the microsomes from pyrethroid-resistant strains metabolised more permethrin than their susceptible counterpart. These findings suggest that prior exposure to pyrethroids can lead to resistance to other pollutants/insecticides and vice-versa.
The highest percentage depletion in both studies with microsomes and recombinant P450s was observed in the Auyo microsomal metabolism of fluorene (73.3 ± 0.4%) and was thus further characterized to understand the kinetics and turnover of this reaction. The Km (58.69 ± 20.47 µM) Kcat (4.20 ± 0.44 min^−1^) were higher than those previously obtained with recombinant P450s metabolism of insecticides. This might be because microsomal fractions contain a mixture of many P450s with varied affinity towards the substrate, thereby compounding the catalytic effects. Examples include the case of metabolism of ecdysone by microsomes of the African migratory locust [72], metabolism of permethrin and deltamethrin by the duplicated An. funestus CYP6P9a/b [42], permethrin and deltamethrin metabolism by An. funestus CYP6AA1 [43] and deltamethrin metabolism by An. minimus CYP6AA3.
Cytochrome P450 monooxygenases, are important Phase I enzymes [73] in the metabolism of xenobiotics and have been implicated as the major players in conferring resistance to insecticides in disease vectors. A large number have been extensively characterized and found to confer resistance to certain insecticides through metabolism, sequestration and excretion of the soluble metabolites. Some of the notable ones associated with pyrethroid resistance in An. gambiae and An. funestus were investigated for their potential in metabolising PAHs, thereby understanding their cross-resistance abilities. CYP6P3 is one of the most extensively characterized P450s in An. gambiae [74, 75] and promiscuous in its ability to metabolise a wide range of compounds, including type I and II pyrethroids [45]. In this study, it metabolised fluorene and fluoranthene with more than 40% depletion, suggesting its potential role in cross-resistance between PAHs and pyrethroids. An. funestus CYP6P9a is an orthologue of the An. gambiae CYP6P3 and equally very promiscuous in metabolising a wide range of pyrethroid and non-pyrethroid insecticides, however, it did not metabolise the PAHs [37, 38, 40]. Furthermore, despite the major role of CYP6M2 in pyrethroid metabolism and resistance [44], there was no activity observed with all three select PAHs, suggesting no direct activity on PAHs despite its role in pyrethroid metabolism.
CYP6Z2 serves as an important marker of pyrethroid resistance although it metabolises the carboxylesterase metabolites of pyrethroid metabolism (3-phenoxybenzoic alcohol and 3-phenoxybenzaldehyde) rather than the parent pyrethroid insecticides [49, 76]. Both CYP6Z2 and CYP6Z3 demonstrated strong metabolic activity towards fluorene and fluoranthene, indicating their potential role in the survival of An. gambiae in polluted breeding sites. Earlier studies have suggested that the CYP6Z family of cytochrome P450s in An. gambiae are most notably expressed at the early life stages of larvae and pupae [76, 77]. This means they inherently help mosquitoes’ timely survival in polluted breeding sites due to their characteristic broad range substrate specificity capable of metabolizing a wide range of compounds, including plants’ secondary metabolites [78]. This similar trend was also reported in Aedes aegypti mosquito’s CYP6Z8, an orthologue of the An. gambiae CYP6Z2 showing catalytic activity on a broad range of substrates, including α-naphthoflavone, resveratrol, and diethylstilbestrol [76]. Naphthalene was not metabolized by any of the P450s used in this study, indicating the lack of cross-resistance potentials between pyrethroids and naphthalene. This may be related to its characteristic chemical structure that gives it an additional stability compared to higher molecular weight PAHs. A similar trend has also been observed in the case of some recombinant P450s metabolising some compounds but not others in the same group. For example, An. funestus CYP9K1 metabolises type II pyrethroids (deltamethrin) but not type I pyrethroids like permethrin [41]. Another example is also the metabolism of all pyrethroids by recombinant An. arabiensis CYP6P4 except deltamethrin (a type II pyrethroid. Here, in both microsome and recombinant P450s studies, fluorene was more highly metabolized than fluoranthene, while naphthalene remained consistently the least metabolized of them all. This differential metabolism may be related to the fact that naphthalene, with lower molecular weight, was less prone to metabolism by Anopheles P450s compared to fluorene and fluoranthene. The findings in this study indicate the potential mechanisms through which environmental pollutants can serve as additional selection pressure in the evolution/escalation of insecticide resistance. This will have an implication on the deployment of insecticide resistance management strategies. However, some of the major limitations include the failure to establish the identity of metabolites generated by P450 depletion of PAHs to rule out a potential bioactivation scenario. In vivo studies with PAHs to ascertain phenotypes would also have added value to the findings. Based on our findings, we recommend using synergists in vector control tools, as the P450s (and other metabolic genes) have evolved to have a wider range of substrate specificities. Another recommendation is paying attention to environmental pollutants in resistance management strategies instead of only focusing on insecticides.
Conclusion
Cytochrome P450s associated with pyrethroids resistance were found to metabolise PAHs in metabolic assays, confirming the potential for cross-resistance between these pollutants and insecticides of public health importance. These findings highlight the importance of the early assessment of the potential cross-resistance liabilities of P450s associated with pyrethroid or other xenobiotic metabolism to new chemistries being introduced for vector control. Future studies that seek to establish the resistance/or susceptibility of malaria vectors to major environmental pollutants will be of great importance to resistance management strategies currently in use. In vivo validation of the role of metabolic P450s in the metabolism/resistance to pollutants is also recommended. Wider studies that include many metabolic enzymes, including the P450s, GSTs, and carboxylesterases, to give a better picture of the cross-resistance relationship between environmental pollutants and public health insecticides.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Corbel V, Guessan RN. Distribution, mechanisms, impact and management of insecticide resistance in Malaria Vectors: a pragmatic review. In: Anopheles mosquitoes - New insights into malaria vectors. Intech open science open mind. 2013. pp. 579–633. Available from: http://www.intechopen.com/books/anopheles-mosquitoes-new- insights-into-malaria-vectors%0A Interested.
- 2Tchouakui M, Ibrahim S, Mangoua K, Thiomela R, Assatse T, Ngongang-Yipmo S, et al. Substrate promiscuity of key resistance P 450s confers clothianidin resistance whilst increasing chlorfenapyr potency in malaria vectors. 114566 ISSN 2024;43(8):2211–47. 10.1016/j.celrep.2024.114566. 10.1016/j.celrep.2024.114566 PMC 1137244139088320 · doi ↗ · pubmed ↗
- 3Yunta C, Hemmings K, Stevenson B, Koekemoer LL, Matambo T, Pignatelli P, et al. Cross-resistance profiles of malaria mosquito P 450s associated with pyrethroid resistance against WHO insecticides. Pestic Biochem Physiol. 2019;161(June):61–7. Available from: 10.1016/j.pestbp.2019.06.007. 10.1016/j.pestbp.2019.06.00731685198 · doi ↗ · pubmed ↗
- 4Azrag RS, Mohammed BH. Anopheles arabiensis in Sudan: a noticeable tolerance to urban polluted larval habitats associated with resistance to Temephos. Malar J. 2018;17(1):1–11. Available from: 10.1186/s 12936-018-2350-1. 10.1186/s 12936-018-2350-1PMC 596019029776357 · doi ↗ · pubmed ↗
- 5Poupardin R, Riaz MA, Jones CM, Chandor-Proust A, Reynaud S, David JP. Do pollutants affect insecticide-driven gene selection in mosquitoes? Experimental evidence from transcriptomics. Aquat Toxicol. 2012;114–115:49–57. Available from: 10.1016/j.aquatox.2012.02.001. 10.1016/j.aquatox.2012.02.00122406618 · doi ↗ · pubmed ↗
- 6Knights KM, Stresser DM, Miners JO, Crespi CL. In vitro drug metabolism using liver microsomes. Curr Protoc Pharmacol. 2016(1). 10.1002/cpph.9.10.1002/cpph.927636111 · doi ↗ · pubmed ↗
- 7Lees RS, Ismail HM, Logan RAE, Malone D, Davies R, Anthousi A, et al. New insecticide screening platforms indicate that mitochondrial complex I inhibitors are susceptible to cross-resistance by mosquito P 450s that metabolise pyrethroids. Sci Rep. 2020;10(1):1–10. Available from: 10.1038/s 41598-020-73267-x. 10.1038/s 41598-020-73267-x PMC 753070233004954 · doi ↗ · pubmed ↗
- 8Mugenzi LMJ, Tekoh A, Ibrahim TS, Muhammad S, Kouamo A, Wondji M, et al. MJ,. The duplicated P 450s CYP 6P 9a/b drive carbamates and pyrethroids cross-resistance in the major African malaria vector Anopheles funestus. P Lo S Genet. 2023;19(3):e 1010678. Available from: 10.1371/journal.pgen.1010678. 10.1371/journal.pgen.1010678 PMC 1008931536972302 · doi ↗ · pubmed ↗