Restoring compromised blood-retina-barrier integrity with Netrin-1 overexpression
Jessica Furtado, Thomas E. Zapadka, Hyojin Park, Kevin Boyé, Jonathan B. Demb, Anne Eichmann

TL;DR
This study shows that increasing Netrin-1 levels can strengthen the blood-retina barrier, potentially preventing vision loss in diseases like diabetic retinopathy and AMD.
Contribution
The study demonstrates that Netrin-1 overexpression selectively stabilizes the blood-retina barrier without significantly affecting retinal angiogenesis.
Findings
Global NTN1 overexpression reduced vascular leak in the P5 angiogenic front and pathological models like OIR and CNV.
Netrin-1/Unc5b signaling was identified as a key pathway for BRB integrity, with distinct effects on vascular leak and angiogenesis.
Unc5b deletion replicated vascular leak seen in Ntn1-deficient retinas, highlighting the pathway's role in BRB stability.
Abstract
The blood-retina barrier (BRB) protects retinal neuronal function and enables vision. A compromised, leaky BRB is a hallmark of vision-threatening retinal diseases such as diabetic retinopathy (DR) and wet age-related macular degeneration (AMD) that affect millions of persons worldwide. Strategies to enhance BRB integrity hold promise as therapeutic interventions to prevent vision loss. Previous studies identified Netrin-1 (NTN1) as a key regulator of BRB stability and revealed reduced Netrin-1 signaling in DR patients, suggesting that Netrin-1 supplementation could help preserve BRB function and prevent disease progression. Herein, we used inducible genetic NTN1 overexpression to investigate effects on BRB development and maintenance. We show that global NTN1 overexpression converted leaky vessels at the P5 angiogenic front into a non-leaky state. In pathological settings, NTN1…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
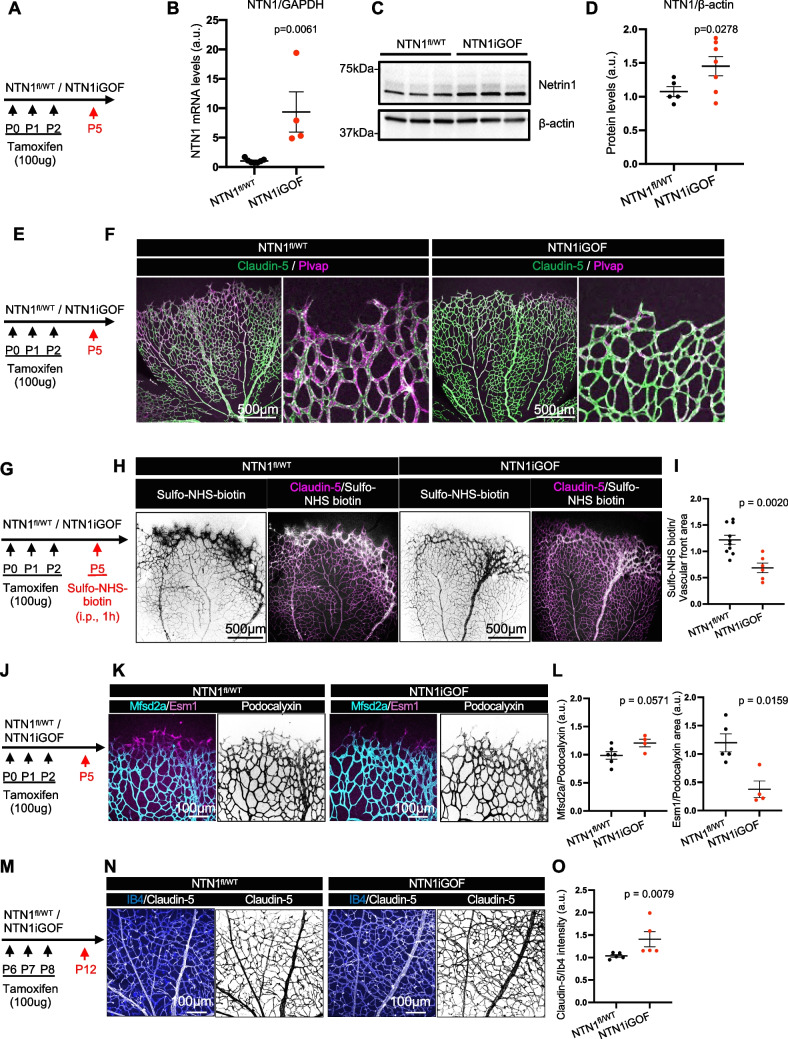 Figure 1
Figure 1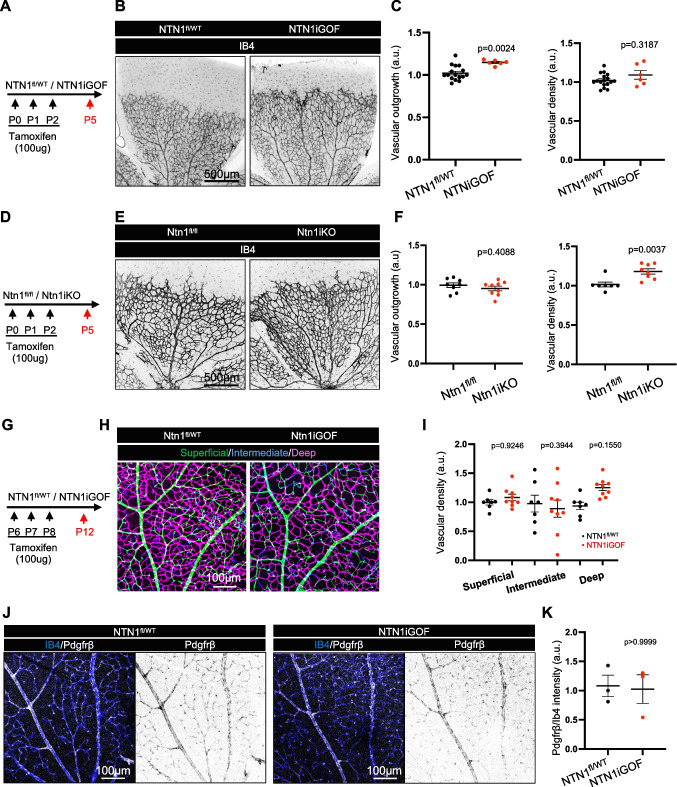 Figure 2
Figure 2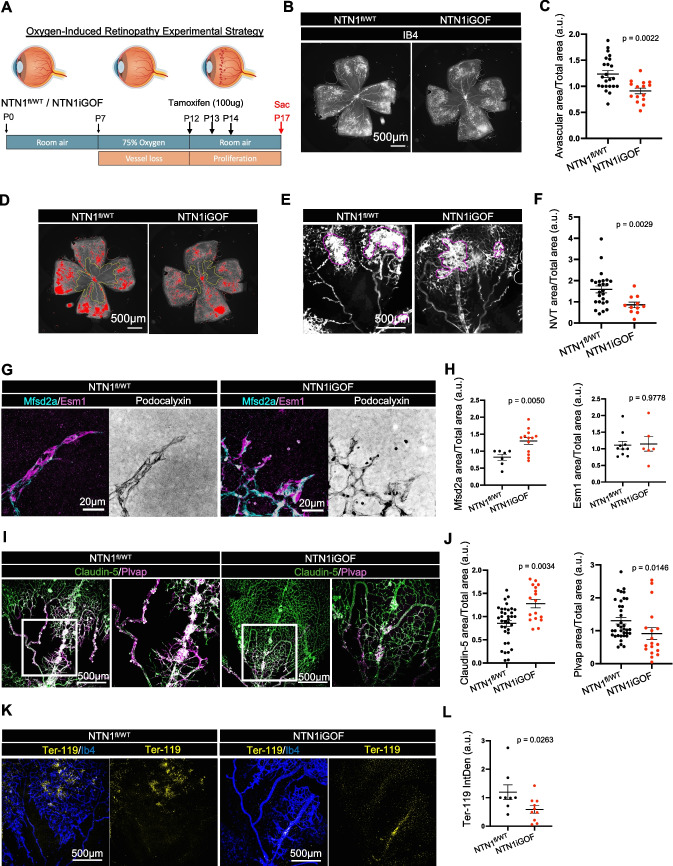 Figure 3
Figure 3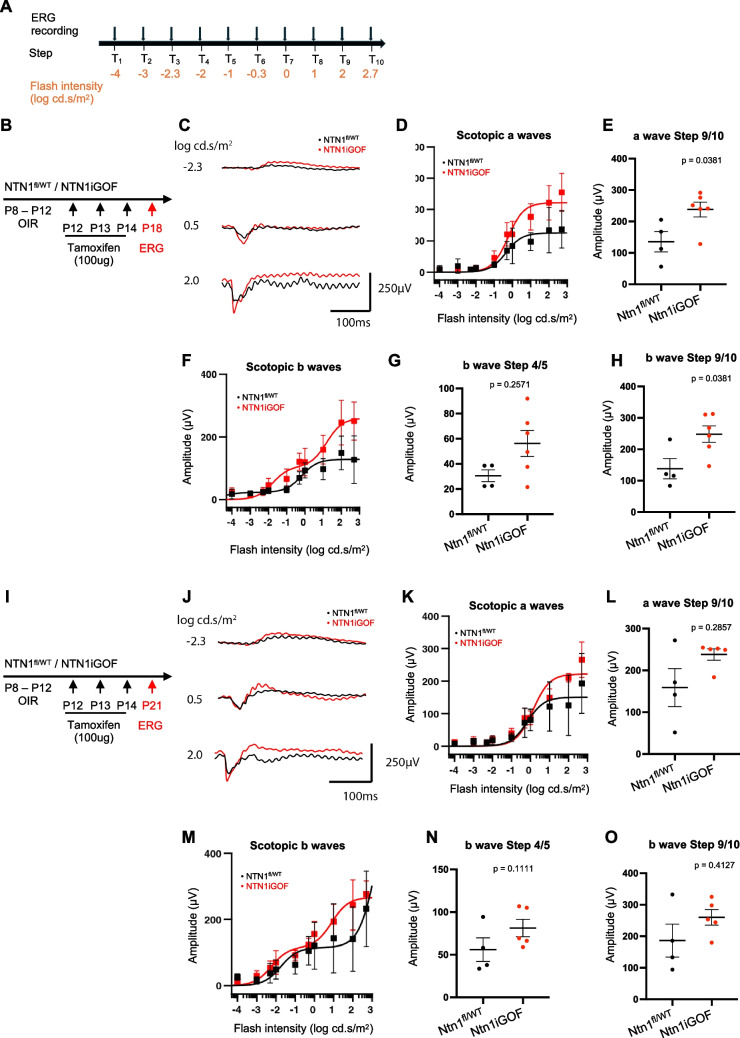 Figure 4
Figure 4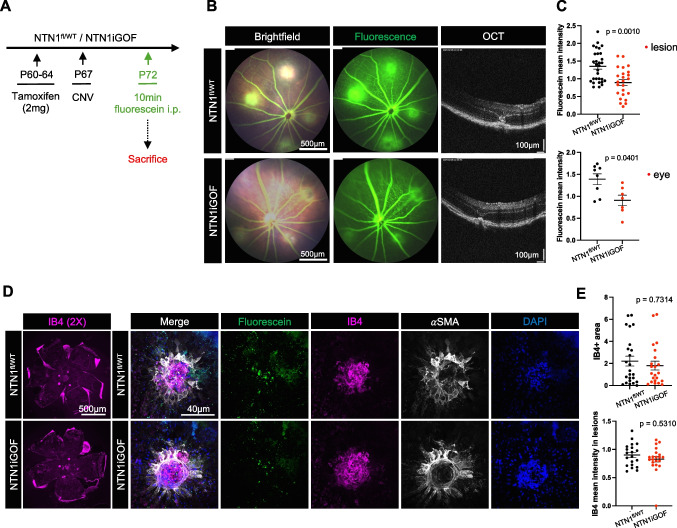 Figure 5
Figure 5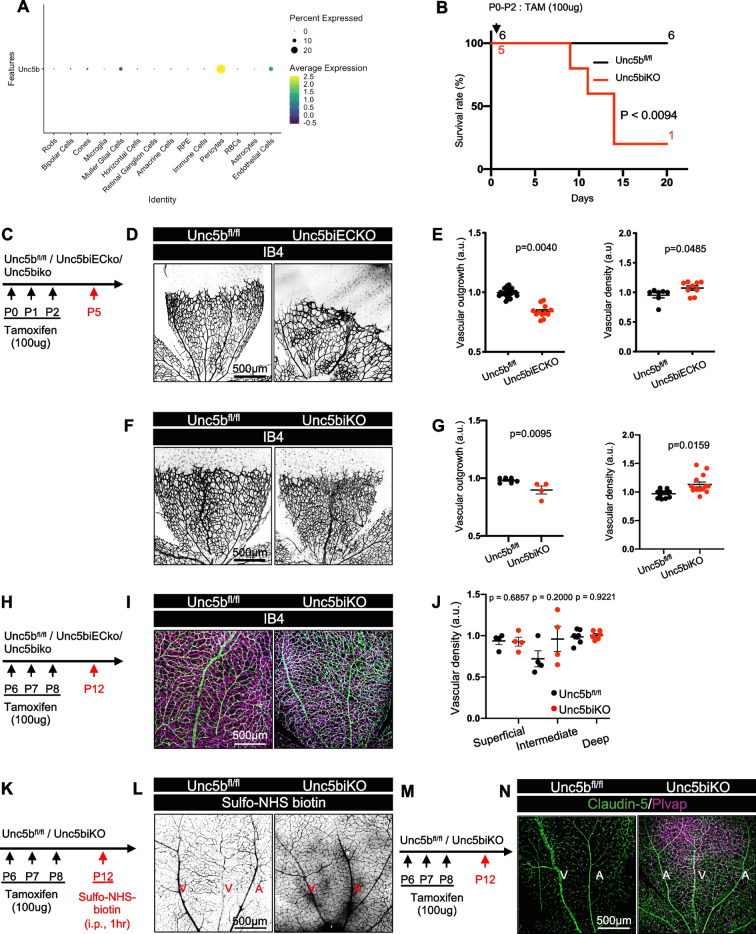 Figure 6
Figure 6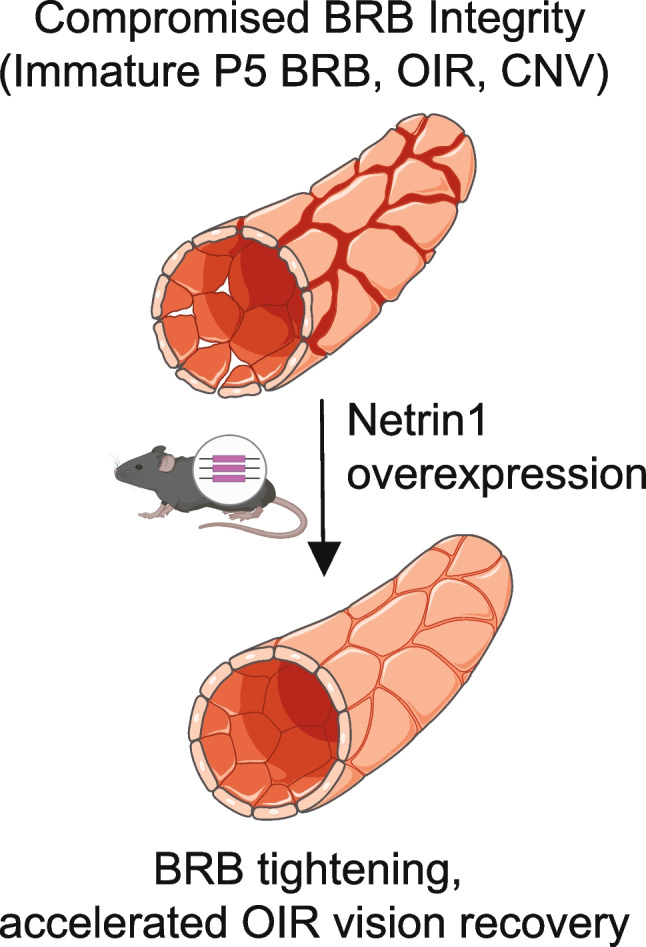 Figure 7
Figure 7- —http://dx.doi.org/10.13039/100017540NHLBI Division of Intramural Research
- —European Research Council
- —http://dx.doi.org/10.13039/100000968American Heart Association
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAxon Guidance and Neuronal Signaling · Retinal Development and Disorders · Hippo pathway signaling and YAP/TAZ
Introduction
The blood-retina barrier (BRB) is a highly selective vascular barrier that tightly regulates the movement of molecules between the bloodstream and the neural retina, thereby maintaining the specialized microenvironment necessary for retinal function [1]. The BRB consists of an inner barrier formed by retinal endothelial cells (ECs) and an outer barrier formed by retinal pigment epithelial (RPE) cells, both of which utilize tight junctions to restrict permeability while allowing controlled nutrient and waste exchange. Disruption of the integrity of the inner BRB accelerates the development of multiple diseases including diabetic retinopathy (DR) and wet age-related macular degeneration (AMD) by leading to vascular leak, fluid accumulation and inflammation that cause neuronal damage and vision loss [2]. Similar breakdown of the blood–brain barrier is involved in diseases such as multiple sclerosis, Alzheimer’s disease and stroke [2–6].
DR is the leading cause of vision loss worldwide, where functional impairments in vision are driven by retinal blood vessel leakage and macular edema [7]. More than 30% of patients with diabetes are reported to have DR [8]. Current treatments include laser photocoagulation, intravitreal steroids, and frequent intravitreal injections of vascular endothelial growth factor (VEGF) inhibitors, alongside blood sugar control. While effective, anti-VEGF therapies require repeated injections and can have off-target effects on retinal neuronal cells that express the signal-transducing VEGFR2 [9–11]. Patients receiving systemic anti-VEGF treatments reported side effects, including hypertension, bleeding, vascular injury, proteinuria, impaired wound healing, and gastrointestinal perforation [12]. To circumvent these side-effects, anti-VEGF therapy for DR and AMD is administered intravitreally [13, 14].
Neovascular ‘wet’ AMD is another major retinal disease that leads to vision loss. Two distinct forms, dry and wet AMD are distinguished. Dry AMD, the more common form, is characterized by progressive thickening of Bruch’s membrane beneath the RPE, alongside gradual degeneration of photoreceptors, the RPE itself and the underlying choriocapillaris. This progressive damage to the surrounding tissue results in disruption of the BRB, contributing to retinal ischemia and inflammation. The more severe, wet form of AMD is driven by choroidal neovascularization (CNV), where abnormal blood vessels from the choroid invade the RPE and extend into the subretinal space, leading to fluid accumulation, increased vascular permeability, and vision loss. The breakdown of the BRB in wet AMD promotes the leakage of fluid and proteins from newly formed blood vessels, exacerbating retinal damage and accelerating disease progression.
The pathogenesis of AMD remains poorly understood and current treatment options are limited. In wet AMD, frontline therapeutics include intravitreal injections of VEGF inhibitors, such as bevacizumab and ranibizumab, which target pathological neovascularization and vascular leakage, are the mainstay of therapy [15]. More recently, bispecific antibodies targeting both VEGF and angiopoietin-2 (Ang-2) have been developed to improve treatment efficacy and reduce the frequency of injections [16]. Preclinical treatment with antibodies blocking the Slit2 receptors Roundabout1 and 2 reduces ocular inflammation and synergizes with anti-VEGF treatment in preventing mouse OIR and CNV [17]. However, despite these and other advances, AMD remains a major cause of irreversible blindness, highlighting the need for novel therapeutic strategies.
Netrin-1 is a key axon guidance molecule that has emerged as a crucial regulator of angiogenesis and BRB integrity [18, 19], and its dysregulation is implicated in DR pathogenesis [20–22]. Netrin-1 enhances retinal revascularization in OIR and diabetic models, while its knockdown using shRNA- reduced pathological angiogenesis [20, 21]. Netrin-1 also modulates vascular permeability in DR [22]. MMP-9, which is upregulated in DR, cleaves Netrin-1 into a bioactive VI-V fragment that increases vascular permeability and diabetic macular edema, and inhibiting MMP-9 prevents this pathological permeability. In contrast, full length Netrin-1 preserves endothelial integrity [20, 23]; thus underscoring the therapeutic potential of preserving Netrin-1 signaling in retinal vascular disease [22].
We previously demonstrated that Netrin-1 maintains BRB integrity by activating endothelial Unc5b and enhancing Norrin–β-catenin signaling. [19, 24]. Single cell RNA sequencing (scRNA-seq) of retinal ECs identified a BRB gene expression program that is regulated by Netrin-1 and Unc5b [19]. Additionally, UNC5B is essential for maintaining stability at endothelial cell–cell junctions in HUVECs and in retinal plexus branching [25]. These findings reveal the essential role of Netrin-1 in maintaining BRB integrity and suggest that targeting Netrin-1 signaling may restore barrier function and prevent vascular leakage, offering a novel therapeutic approach for DR and related neurovascular diseases.
In this work, we used genetic NTN1 overexpressing mice to determine effects on BRB integrity. We also employed the OIR and Laser-Induced CNV models of pathological leak, and we investigated the effect of NTN1 overexpression on OIR induced vision impairment. We found that overexpressing Netrin1 stabilized the BRB in both physiological and pathological models of permeability, and improved vision recovery in OIR. In addition, we found that NTN1 overexpression minimally and transiently affected retinal angiogenesis in an Unc5b independent manner, while its BRB stabilizing effects were Unc5b dependent. Together, these findings position Netrin-1-Unc5b signaling agonists as promising therapeutic candidates for restoring vascular integrity and visual function in retinal diseases characterized by BRB breakdown.
Materials and methods
Mouse models
All animal experiments were performed under a protocol approved by the Institutional Animal Care Use Committee of Yale University. All mice were of the C57BL/6 strain. Inducible Netrin-1 gain of function mice (gift from Patrick Mehlen, Lyon University) were generated by intercrossing loxStoplox;hNTN1 [26] with RosaCre^ERT2^ (Jackson laboratories) driver to induce ubiquitous overexpression of NTN1. Ntn1^fl/fl^ mice [27] crossed with ROSACre^ERT2^ mice were described previously. Unc5b^fl/fl^ mice [24] were bred with Cdh5Cre^ERT2^ (endothelial deletion) (Taconic, Biosciences), or with RosaCre^ERT2^ (global deletion). Entire litters containing male and female mice were used for experiments analyzed at P5 and P12 because the sex of the mice could not be distinguished. Both males and females were also used for experiments analyzed at P17, P21 and P60.
Western blot
Retinas were dissected and frozen in liquid nitrogen and lysed in RIPA buffer (Research products, R26200-250.0) supplemented with protease and phosphatase inhibitor cocktails (Roche, 11836170001 and 4906845001) using a Tissue-Lyser (5 times 5 min at 30 shakes/second). Protein lysates were centrifuged 15 min at 13200 rpm at 4 °C and supernatants were isolated. Protein concentrations were quantified by BCA assay (Thermo Scientific, 23225) according to the manufacturer’s instructions. 30ug of protein was diluted in Laemmli buffer (Bio-Rad, 1610747) boiled at 95 °C for 5 min and loaded in 4–15% acrylamide gels (Bio-Rad, 5678084). After electrophoresis, proteins were transferred on a nitrocellulose membrane and incubated in TBS 0.1% Tween supplemented with 5% BSA for 1 h to block non-specific binding. The following antibodies were incubated overnight at 4 °C: Ntn1 (mouse anti-Netrin-1 R&D, AF1109, which shows cross-reactivity with recombinant human Netrin-1), β-actin (Sigma, A1978), Unc5b (Cell Signaling Technology #13851). Western blot densitometry analysis was conducted using ImageJ (version 1.54 g).
Tracer injection and immunostaining
Sulfo-NHS-biotin tracer was injected intraperitoneally in P12 mice at a concentration of 10 mg/mL (300μL per mouse) and left to circulate for 1 h. Retinas from mice injected with tracers were dissected in 3.7% formaldehyde to ensure dye fixation within the tissue during retina dissection. Each retina was placed in 3.7% formaldehyde at room temperature for 10 min, stored in 100% methanol overnight, washed 3 × 10 min with PBS, dissected and incubated with specific antibodies in blocking buffer (1% fetal bovine serum, 3% BSA, 0.5% Triton X-100, 0.01% Na deoxycholate, 0.02% Sodium Azide in PBS at pH 7.4) overnight at 4 °C. The following antibodies were used: Unc5b (Cell signaling, 13851S), Claudin-5-GFP (Invitrogen, 352588), Plvap (BD biosciences, 550563), Mfsd2a (Cell Signaling, 80302S), Esm1 (Biotechne, AF1999), Alexa-Fluor 647-Anti-ERG (abcam, AB196149), TER-119 (Invitrogen, 14–5921-82). IB4 as well as all corresponding secondary antibodies were purchased from Invitrogen as donkey anti-primary antibody coupled with either Alexa Fluor 488, 568 or 647. Streptavidin-Texas Red (Vector Laboratories, SA-5006–1) was used to detect sulfo-NHS-biotin. The next day, retinas were washed 3 × 10 min and incubated with secondary antibody for 2 h at room temperature, then washed 3 × 10 min and post-fixed with 0.1% PFA for 10 min and mounted using fluorescent mounting medium (DAKO, USA).
Microscopy and image analysis
Confocal images were acquired on a laser scanning fluorescence microscope (Leica SP8) using the LASX software. 10X, 20X and 63X oil immersion objectives were used for acquisition using selective laser excitation (405, 488, 547, or 647 nm). Quantification of vascular outgrowth and quantification of pixel intensity were performed using the software ImageJ (version 1.54 g). Vascular density was quantified with the software Angiotool by quantifying the vascular surface area normalized to the total surface area. The superficial, intermediate and deep layers were separated and colored using ImageJ z-stack functions. Quantification of pixel intensity was performed using the software ImageJ. Avascular area in retinas from mice subjected to OIR was quantified on ImageJ by measuring the avascular area and normalizing to the total area of a retina. Claudin-5 was quantified using area of green pixels divided by area of all pixels. Plvap was quantified using area of magenta pixels divided by area of all pixels. Fluorescein leak was quantified by creating an area around a lesion based on a brightfield image and then quantifying intensity using the same ROI in the fluorescence channel. Quantification of TER-119 Integrated Density was measured on ImageJ. Integrated density is Mean intensity X area. CNV quantifications were done by creating an ROI of a lesion based on the brightfield image of the lesion, and then quantifying the fluorescence intensity of that ROI in the corresponding fluorescence channel.
Oxygen-induced retinopathy
P7 mice were placed in a hyperoxia chamber with the dam. The chamber oxygen level was set to 75% and the chamber was closed from P7 to P12. On P12 the pups were returned to room air with a nursing foster mother. At P12, P13 and P14 mice were given TAM injections (200 mg/mouse per day). Eyes were then assessed at P17 for the maximum neovascular response for OIR.
Laser-induced choroidal neo-vascularization (CNV)
Adult mice were briefly anesthetized with ketamine (100 mg/kg) and xylazine (10 mg/kg). 1% Tropicamide (Somerset) was applied on eyes for 5 min to dilate pupils. The tropicamide was then wiped off and a drop of GenTeal (Alcon Professional) was applied to the eyes. Four focal burns were administrated in each eye using a 532 nm argon ophthalmic laser via a slit lamp, being careful to avoid large blood vessels. One week post injury mice were anesthetized with ketamine (100 mg/kg) and xylazine (10 mg/kg) and intraperitoneally injected with 10 mg of Sodium Fluorescein approximately 10 min prior to imaging, and lesions were imaged. Optical coherence tomography (OCT) images were acquired using a Phoenix Micron IV. Shortly after, mice were sacrificed, enucleated and eyes were fixed in 4% PFA for 10 min, before choroids were dissected for whole mount staining.
Electroretinography
Electroretinography was performed as previously described [28]. Mice were dark-adapted overnight before being anesthetized with ketamine (100 mg/kg) and xylazine (10 mg/kg). To maintain body temperature, the mice were placed on a temperature regulating heating pad. Pupils were dilated by applying 1% Tropicamide to the eyes for 5 min, followed by positioning gold wire corneal electrodes (LKC Technologies, SKU: N30). Electroretinograms (ERGs) were recorded simultaneously from both eyes. Scotopic responses were obtained using single-flash stimuli ranging from ~ 4.0 log cd·s/m^2^ to + 2.7 log cd·s/m^2^. The a-wave amplitude was calculated as the negative deflection occurring within the first 60 ms post-flash, while the b-wave amplitude was identified as the maximum positive deflection following the a-wave. A 55-Hz Bessel filter was applied to remove oscillatory potentials prior to measuring the b-wave amplitude. Mice were monitored for 24 h after anesthesia administration.
Results
NTN1 overexpression enhances BRB integrity during postnatal retinal development
Inducible Netrin-1 gain of function mice were generated by intercrossing mice harboring a human NTN1 transgene (loxStoplox;hNTN1) [26] with RosaCre^ERT2^ mice*.* Human Netrin-1 is nearly identical to murine Netrin-1 (≈99% amino acid identity) and the transgene encodes full-length Netrin-1 protein. Ubiquitous heterozygous NTN1 overexpression was induced by TAM injections from P0-2 (hereafter NTN1iGOF) and TAM-injected Cre-negative NTN1fl/WT littermates were used as controls (Fig. 1A).Fig. 1. Netrin-1 overexpression prevents vascular leak in neonatal retinas. (A) NTN1 overexpression strategy in neonates. (B) qPCR analysis of P5 retina mRNA extracts. (C) Western blot and quantification (D) of P5 retina protein extracts. (E) NTN1 overexpression strategy and (F) immunofluorescence of whole-mount P5 retinas with the indicated antibodies. (G) NTN1 overexpression and tracer injection strategy. (H) P5 retinas i.p. injected with Sulfo-NHS-biotin for 1 h and stained for Claudin-5 and (I) quantification of leak at the angiogenic front. (J) Overexpression strategy and (K) retina staining with the indicated antibodies and (L) quantification of K. (M) Overexpression strategy and (N) P12 retinas stained with indicated antibodies and (O) quantification of N. Each dot represents one retina from one mouse. At least 4 mice were used per genotype for each experiment. Data are shown as mean ± SEM. Two-sided Mann–Whitney U test was performed for statistical analysis
Subsequent qPCR analysis of P5 retinas with primers that detect human NTN1 revealed increased NTN1 mRNA levels in NTN1iGOF mice compared to littermate controls (Fig. 1B). Western blot of retina lysates using an antibody that detects mouse Netrin-1 and cross-reacts with human Netrin-1 confirmed increased levels of a 68 kDa band in NTN1iGOF mice compared to littermate controls (Fig. 1C,D). We did not detect any cleavage fragments, consistent with production of full-length Netrin-1 protein in the NTN1iGOF mice (Supp. Figure 1).
As knockout of Ntn1 and its receptor Unc5b destabilizes the BRB [19], we anticipated that conversely, overexpressing NTN1 would enhance barrier properties. Towards this goal, we investigated expression of Claudin-5, a tight junction protein, and Plvap, a permeability protein expressed in fenestrae and transcytotic vesicles in P5 retinas. The vascular front of NTN1^fl/WT^ littermate control mice was Plvap + and Claudin5- (Fig. 1E, F) and displayed higher Claudin-5 expression in more mature vessels closer to the optic nerve, and lower expression at the angiogenic front (Fig. 1F). Conversely Plvap expression was enriched in the leaky front but suppressed in mature vessels (Fig. 1F). By contrast, NTN1iGOF ECs at the vascular front converted to a Claudin-5 +/Plvap- phenotype (Fig. 1F), consistent with acquisition of barrier function. Intraperitoneal (i.p.) injection of Sulfo-NHS biotin into P5 NTN1iGOF mice showed significantly reduced BRB leakage at the angiogenic front compared to Cre-negative littermate controls (Fig. 1G,H,I), attesting to enhanced vascular barrier formation in superficial tip cells. Probing for expression of the BRB-associated lipid transporter protein Mfsd2a revealed low levels of Mfsd2a in Esm1 + and Podocalyxin + tip cells at the angiogenic front of Cre-negative littermate controls, as expected from prior studies [29, 30] (Fig. 1J). However, NTN1iGOF mice showed mildly increased levels of Mfsd2a in Podocalyxin + tip ECs at the vascular front, and reduced Esm1 levels in Podocalyxin + tip cells compared to controls, suggesting that molecules regulating barrier properties are induced [29] (Fig. 1 K,L). At P12, when retina vessels have acquired a functional BRB [19, 30], the NTN1iGOF mice still displayed increased Claudin-5 immunostaining levels compared to littermate controls (Fig. 1M-O) suggesting a reinforced endothelial barrier.
Netrin-1 exerts a minimal and transient effect on retinal angiogenesis
Netrin-1 has been previously shown to affect angiogenesis [18, 31–34]. Following TAM injections from P0-2, IB4 staining of P5 NTN1iGOF retinas revealed a small but significant increase in vascular outgrowth compared to littermate controls but no change in vascular density (Fig. 2A-C). Conversely, Ntn1iKO (inducible knockout) mice generated by intercrossing Ntn1fl/fl mice with ROSA-Cre^ERT2^ and TAM-treated at P0-2 and analyzed at P5 like the GOF mice displayed no change in vascular outgrowth and a small but significant increase in vascular density compared to their littermate controls (Fig. 2D-F). We had previously reported that TAM treatment of Ntn1iKO mice between P6 and P12 produced no change in angiogenesis of the superficial, intermediate and deep vascular layers [19]. Likewise, P12 NTN1iGOF retinas from mice injected with TAM at P6 – P8 showed similar vascular density in superficial, intermediate and deep plexus compared to controls (Fig. 2G-I). Additionally, staining for the pericyte marker Pdgrfβ revealed no differences in pericyte coverage of IB4 + vessels between NTN1iGOF retinas and controls at P12 (Fig. 2J,K). These data show that Netrin-1 has a small and transient effect on early retinal angiogenesis, but sustained effects on BRB integrity.Fig. 2. Netrin-1 promotes angiogenesis of the superficial retinal vascular plexus. (A) NTN1 overexpression strategy in neonates. (B) Immunofluorescence of whole-mount P5 retinas stained with Ib4. (C) Quantification of vascular outgrowth and density. (D) Ntn1 gene deletion strategy in neonates. (E) Immunofluorescence of whole-mount P5 retinas stained with IB4. (F) Quantification of vascular outgrowth and density. (G) NTN1 overexpression strategy, (H) P12 retina staining and (I) quantification of IB4 labeled vascular layers in indicated genotypes. (J) Immunofluorescence of whole-mount P12 retinas stained with IB4 and Pdgfrβ and (K) quantification. Each dot represents one retina from one mouse. At least 3 mice were used per genotype for each experiment. Data are shown as mean ± SEM. Two-sided Mann–Whitney U test was performed for statistical analysis between two groups, ANOVA followed by Bonferroni’s multiple comparisons test was performed for statistical analysis between multiple groups
NTN1 overexpression rescues pathological vascular leakage in OIR
To investigate whether NTN1 overexpression could inhibit leak in ocular neovascular disease, we used the OIR model [35] (Fig. 3A). Retinas of mice exposed to 75% oxygen (hyperoxia) between P7 to P12 develop vaso-obliteration and a capillary free avascular area in the center of the retina. Upon return to room air at P12, the relative hypoxia induced release of excessive amounts of angiogenic factors, which results in pathological sprouting and formation of leaky and hemorrhagic neovascular tufts (NVTs). When NTN1iGOF was induced by TAM administration during the revascularization process, retinas displayed decreased avascular areas and decreased neovascular tuft areas compared to littermate controls (Fig. 3B-F). This indicates a shift toward productive angiogenesis, facilitating revascularization, while simultaneously suppressing pathological angiogenesis and NVTs. To understand this process better, we focused on OIR tip cells that express Podocalyxin as well as high levels of Esm1 and low levels of Mfsd2a [29]. Podocalyxin + tip cells in the NTN1^fl/WT^ control OIR sprouting regions were positive for the tip-cell marker Esm1 with limited Mfsd2a expression, but Mfsd2a expression was significantly increased in Ntn1iGOF retinas, suggesting improved barrier stability (Fig. 3G,H). Additionally, while control littermate retinas demonstrated the presence of Plvap and diminished expression of Claudin-5 in the arterioles and venules of P17 OIR retinas, littermate NTN1iGOF retinas had increased Claudin-5 expression and suppressed Plvap expression (Fig. 3I,J). OIR retinas typically demonstrated hemorrhagic injury and leakage of Ter-119 + red blood cells as was seen in control NTN^fl/WT^ retinas and quantified by measuring the mean intensity times area of Ter-119 + staining. NTN1iGOF mice displayed decreased Ter-119 + cells as compared to controls (Fig. 3K,L), demonstrating protection against OIR hemorrhagic injury.Fig. 3. Oxygen-Induced Retinopathy in NTN1iGOF mice. (A) OIR and overexpression strategy. (B) Whole-mount P17 OIR retinas stained with IB4. (C) Quantification of Avascular area. (D) Whole-mount OIR P17 retinas. Red indicates NVTs and yellow boundary line delineates the avascular area. (E) One petal from P17 OIR retina, magenta outlines NVTs (F) Quantification of NVTs. (G) High magnification images of P17 OIR sprouts stained with indicated antibodies and (H) quantification of G. (I) Whole-mount P17 retinas stained with indicated antibodies. (J) Quantification of immunostaining as indicated. (K) Whole-mount P17 OIR retinas stained with indicated antibodies and (L) quantification of TER-119 Integrated Density (Mean intensity X area). Each dot represents one retina. At least 4 mice were used per genotype for each experiment. Data are shown as mean ± SEM. Two-sided Mann–Whitney U test was performed for statistical analysis
Netrin-1 overexpression improves OIR-impaired vision
Retinal dysfunction and vision loss in the OIR retina are characterized by reduced electroretinogram (ERG) amplitudes, including photoreceptor (a-wave) and postsynaptic bipolar cell responses (b-wave) [35, 36]. To determine whether a tightened barrier induced in NTN1iGOF mice improved retinal function in OIR mice, we conducted ERG recordings at P18 and P21. P18 NTN1iGOF mice exhibited enhanced a- and b-wave amplitudes, with increasing light intensities, compared to littermate controls (Fig. 4B-H). Specifically, scotopic a-waves of NTN1iGOF showed significantly increased amplitudes compared to controls (Fig. 4.D,E). Similarly, Scotopic b-waves showed a non-significant increase in step 4/5 (rod-only response) amplitude, and a significant increase in step 9/10 (mixed rod and cone response) amplitude (Fig. 4.F,G,H). Around P21 neovascularization in the retina spontaneously regresses and the retina vasculature recovers around P25 [37]. When we measured ERG amplitudes in the same mice at P21 (Fig. 4I), we observed that responses of NTN1iGOF mice trended towards higher a and b wave amplitudes compared to littermate controls (Fig. 4J-O), but the values were not significantly different due to variability in the response (Fig. 4L,N,O). Overall, these data show that NTN1iGOF accelerates the recovery of visual acuity following OIR.Fig. 4. Netrin-1 overexpression improves OIR-impaired vision. (A) ERG experimental outline. (B) NTN1 overexpression strategy and (C) Averaged scotopic combined traces at P18. (D) A wave amplitudes at P18 and (E) comparison of average response to step 9/10. (F) B wave amplitudes at P18 and (G, H) comparison at step 4/5 and step 9/10. (I) NTN1 overexpression strategy. (J) Averaged scotopic combined traces at P21. (K) A wave amplitudes at P21 and (L) comparison at step 9/10. (M) B wave amplitudes at P21 and (N, O) comparison at step 4/5 and step 9/10. Each dot is the average of the left and right eye responses per mouse per trial. Data are shown as mean ± SEM. Two-sided Mann–Whitney U test was performed for statistical analysis
NTN1 overexpression reduces pathological leakage in choroidal neovascularization
To test effects of NTN1 overexpression in another pathological model of leak, we used a laser-induced model that mimics neovascular/wet AMD [38]. The models consist of laser rupture of the Bruch’s membrane, which triggers the release of VEGF and results in the growth of new blood vessels from the choroid into the subretinal area. This neovascularization is accompanied by pathological leak assayed by measuring fluorescent tracer leak. Adult (P60) male NTN1iGOF mice and littermate controls were injected with TAM for 5 days and subjected to CNV three days later. CNV was induced by 4 distinct laser injuries per retina and vascular leak was assessed with fluorescein injection (i.p. 10 min) 5 days later, which is reported to be the timepoint with maximal leak [39] (Fig. 5A,B). Optical coherence tomography (OCT) imaging confirmed the CNV presence in both groups (Fig. 5B). NTN1iGOF mice displayed markedly decreased fluorescence compared to littermate controls, as quantified by measuring mean fluorescein intensity per lesion and per eye (Fig. 5B,C). Whole mount choroid staining showed no changes in IB4 + lesion size or smooth muscle cell coverage in NTN1iGOF mice compared to controls (Fig. 5D,E), indicating lack of an angiogenic effect in this model.Fig. 5NTN1 overexpression rescues CNV-induced leak. (A) NTN1 overexpression strategy and experimental design. (B) Fundus fluorescein angiography images 5-days post laser injury, and Optical Coherence Tomography (OCT) images 5-days post laser injury and (C) quantification of fluorescein mean intensity per lesion and per eye. Each dot corresponds to one eye, four mice per genotype were analyzed. (D) Whole mount staining of choroids with indicated antibodies and (E) Quantification, each dot corresponds to one lesion
Unc5b is dispensable for Netrin-1–Mediated angiogenesis but required for BRB integrity
To understand if Netrin-1 exerted its angiogenic effect on the superficial retinal vascular plexus via Unc5b, we queried single cell RNA sequencing data from P12 retinas for Unc5b expressing cell types. We observed Unc5b expression mainly in ECs, in mural cells, and in a few Muller glia cells [19] (Fig. 6A). To determine whether Unc5b in pericytes or in Muller glia contributed to retinal angiogenesis or BRB function, we generated global Unc5b knockout mice (hereafter Unc5biKO) by intercrossing Unc5b^fl/fl^ and RosaCre^ERT2^.Fig. 6. Effects of global and endothelial Unc5b deletion on retinal angiogenesis and the BRB. (A) Unc5b expression in retinal scRNA-seq samples. (B) Survival curve after neonatal global Unc5b gene deletion, Mantel-cox test. (C) Gene deletion strategy. (D) Whole-mount P5 retinas of indicated genotypes stained with IB4 and (E) Quantification of vascular outgrowth and density. (F). Whole-mount P5 retinas of indicated genotypes stained with Ib4 and (G) Quantification of vascular outgrowth and density. (H) Gene deletion strategy. (I) IB4 staining and (J) quantification of superficial, intermediate, and deep layers at P12. (K) Unc5b gene deletion and tracer injection strategy. (L) Whole-mount P12 retinas after i.p. injection with sulfo-NHS-biotin for 1 h. (M) Unc5b gene deletion strategy. (N) Whole-mount P12 retinas stained with the indicated antibodies. A: Artery, V: Vein. Data are shown as mean ± SEM. Two-sided Mann–Whitney U test was performed for statistical analysis between two groups, ANOVA followed by Bonferroni’s multiple comparisons test was performed for statistical analysis between multiple groups. Each dot represents one eye. At least 4 mice were used per genotype for each experiment
Neonatal TAM injections between P0-2 induced lethality of Unc5biKO mice by P15 (Fig. 6B) as previously reported with Unc5biECKO mice [24]. Immunostaining and western blot analysis of retinal lysates revealed efficient Unc5b deletion in Unc5biKO retinas (Supp. Figure 2A-C).
To assess retinal angiogenesis, we stained P5 retinal flatmounts from mice injected with TAM at P0-P2 with IB4 and measured vascular density and outgrowth. Both endothelial specific and global Unc5b deletion reduced vascular outgrowth and increased vascular density (Fig. 6C-G). These phenotypes differ from Ntn1iKO mice (see Fig. 2D-F) and lead us to conclude that Unc5b is not required for Netrin-1’s effect on angiogenesis. Global Unc5b deletion between P6-P8, and analysis of retinal vessel density at P12, revealed no significant differences between groups (Fig. 6H-J), highlighting that both Unc5b and NTN1 only affect angiogenesis of the superficial vascular plexus, but in a distinct manner.
To assess BRB permeability, we examined leakage of injected Sulfo-NHS-biotin in P12 mice after gene deletion between P6 and P8 (Fig. 6K). Unc5biKO mice exhibited BRB leakage (Fig. 6M), and ECs in the Unc5biKO mice converted to a Claudin-5-/Plvap + phenotype (Fig. 6M-N), demonstrating loss of BRB integrity. Likewise, P12 Ntn1iKO mice injected with TAM between P6 and P8 displayed Sulfo-NHS biotin leakage and EC conversion to a Claudin-5 low/Plvap high phenotype (Supp. Figure 3 A,B), indicating loss of BRB integrity as previously reported [19].
Discussion
Herein, we report that TAM-inducible, global genetic NTN1 overexpression in mice converted leaky vessels at the P5 angiogenic front into a non-leaky phenotype. Moreover, NTN1 overexpression rescued pathological vessel leak in two ocular neovascular disease models, CNV and OIR. The barrier stabilizing effect in OIR improved electroretinogram recordings as seen by increased ERG amplitudes in P18 mice, attesting that NTN1 promotes functional vision recovery along with BRB properties in this model (Fig. 7).Fig. 7. Working model. NTN1iGOF mice display BRB tightening and rescue of vascular leakage in two mouse models of pathological ocular neovascularization, but only mild transient effects on retinal angiogenesis. As global Unc5b deletion prevents BRB tightening, reinforcing Netrin-1 Unc5b signaling may be a therapeutic strategy to prevent vision loss. Figure was created using BioRender and SMART
At the molecular level, previous work revealed two distinct tip cell populations in the developing postnatal retina [29]. Superficial S tip cells are found at the leaky vascular front of P5 retinas, while D tip cells dive into the underlying neural retina to form two deeper vascular layers starting at P10. Superficial S tip cells are leaky to Sulfo-NHS biotin and express permeability proteins Esm1 and Plvap, while diving D tip cells are non-leaky and express Claudin5 and the lipid transporter Mfsd2a, which are two effectors of BRB integrity [29]. Neither Claudin5 nor Mfsd2a are expressed by superficial S tip cells, and neither Plvap nor Esm1 are expressed by diving D tip cells, hence expression of these proteins is mutually exclusive between leaky S-tip cells and BRB competent D-tip cells. The NTN1iGOF P5 superficial tip cells analyzed herein displayed loss of permeability proteins Plvap and Esm1 and upregulated Claudin5 and Mfsd2a, suggesting that NTN1iGOF converts leaky superficial tip cells into a phenotype that resembles BRB competent, non-leaky diving D-tip cells.
Pathological OIR tip cells express S-like mRNA profiles, including high Esm1 and low Mfsd2a expression [17, 29]. NTN1iGOF upregulated expression of Mfsd2a and Claudin5 in OIR tip cells, while Plvap was suppressed compared to control OIR tips cells and Esm1 remained unchanged, suggesting that NTN1iGOF confers acquisition of some BRB properties. Furthermore, hemorrhagic injury was reduced, and visual recovery occurred earlier in NTN1iGOF mice when compared to controls.
In addition to effects on OIR vascular permeability, NTN1iGOF enhanced revascularization and decreased neovascular tuft formation. Netrin-1 as well as Unc5b and Neogenin expression levels were previously shown to increase in OIR retinas and pro-angiogenic effects of Netrin-1 in OIR were reported before [40–42]. One study reported that OIR-induced retinal neovascularization was successfully treated with Netrin-1 RNAi [41]. Another study conducted in mice lacking the related Ntn4 gene showed that Ntn4 homozygous global knockout mice subjected to OIR accelerated revascularization and recovery of visual acuity measured by ERG, with no effects of loss of Ntn4 knockout on physiological retinal angiogenesis or in a CNV model were detected [43]. Here, NTN1 loss or gain of function had weak and transient effects on angiogenesis. The most pronounced effect was seen in OIR angiogenesis, where avascular area and NVT formation were both reduced by NTN1iGOF, whereas neovascularization after CNV was unaffected. During development of the superficial vascular plexus between birth and P5, NTN1iGOF slightly but significantly increased vascular outgrowth but not vascular density, while Ntn1iKO slightly but significantly increased vascular density but not vascular outgrowth. Neither GOF (this work) nor LOF mutants [19] displayed angiogenic defects when recombination was induced between P6 and P12, i.e. during formation of the deeper vascular layers of the retina. Thus, Netrin-1 has a small and transient effect on angiogenesis, while its effect on BRB integrity persists across all stages and models.
Our previous work showed that Netrin-1’s BRB stabilizing effects are mediated by endothelial Unc5b. Sc-RNA seq analysis showed that both Ntn1iKO and Unc5biECKO downregulated a common BRB gene expression program encoding tight junction proteins, nutrient and ion transporters, Wnt signaling effectors and others [19]. Herein, we extend these data by showing that global Unc5b mutants exhibit similar vascular phenotypes when compared to endothelial Unc5b mutants, including lethality and loss of BRB integrity. As both endothelial and global Unc5b increased vascular leak, additional Unc5b expression in retinal pericytes and Muller glia appears to play a minor role in BRB integrity when compared to endothelial Unc5b. Further studies using pericyte or Muller-glia specific Unc5b deletions should be conducted to confirm this finding.
Likewise, further studies are needed to elucidate the source of Netrin-1 required for its BRB-stabilizing effect. Netrin1 is secreted into the blood and is present at a concentration of 50-150 ng/mL [44, 45]. Pericytes are the only retinal cells that express Netrin-1, but only 8% of pericytes express Ntn1 at low levels [19]. Future studies are required to determine if pericyte Netrin-1 expression is required for its BRB-stabilizing effect, or if the relevant Netrin-1 source is from the bloodstream. In addition, we cannot formally rule out that the effects of NTN1GOF on BRB integrity tightening involve additional receptors besides Unc5b.
In contrast to BRB integrity, angiogenic phenotypes in the P5 retina of NTN1 and Unc5b mutants were clearly distinct. Unc5b deletion in ECs or in all cells increased vascular density, confirming earlier work in global mutants and with function blocking antibodies [18, 33, 34]. Unc5b was identified as a Notch downstream effector and loss or gain of Unc5b limited the ability of Notch activation to regulate EC behaviors [25]. Endothelial-specific Unc5b deletion using the same lines employed here increased branching complexity in the developing retina [25]. While vascular outgrowth was also increased in their series, both studies support a role for Unc5b in limiting vascular branching [25]. In addition to Netrin-1 [33, 46], Unc5b binds Robo4 [34, 47], Flrt2 [48, 49] and Flrt3 [48] via its extracellular domain, and deletion of Flrt3 phenocopied retinal hypervascularization observed in Unc5biECko retinas [50]. These data indicate that Unc5b mediated angiogenesis and barriergenesis are mediated by distinct ligands. Potential receptors mediating NTN1 effect on retinal angiogenesis include CD146 [51], which also binds VEGF [52].
Dysregulation of Netrin-1 expression in human patients with DR was shown to contribute to vascular leak and macular edema [22]. In addition to current treatments for DR and AMD that rely on blocking VEGF-A activity [53–55], recent studies developed agonists that induce BRB and BBB stabilization via activation of β-catenin [56, 57]. Approaches to enhance Netrin-1 binding to endothelial Unc5b could synergize with such therapies and offer broad application through the regulation of both the BRB and the BBB. Given the reported cleavage of NTN1 in DR eyes [22], therapeutic strategies to enhance NTN1 mediated BRB function may need to incorporate cleavage-resistant variants or MMP-9 inhibition to ensure the beneficial effects of intact Netrin-1 are preserved.
Supplementary Information
Below is the link to the electronic supplementary material.Supplementary file1 (PDF 156 KB) Validation of NTN1iGOF. Uncropped Western blot from Fig.1c shows presence of full-length Netrin-1 and absence of bands below 68kDaSupplementary file2 (PDF 1160 KB) Validation of global Unc5b deletion. (A) Whole-mount P5 retinas stained with indicated antibodies. (B) Western blot of retina protein extracts and blot quantification (C). Each dot represents one retina from one mouse. Unc5b^flfl^ n=9, Unc5biKO n= 10Supplementary file3 (PDF 947 KB) Effects of Ntn1 deletion on BRB integrity. (A) Ntn1 gene deletion and tracer injection strategy. (B). Whole-mount P12 retinas after i.p. injection with sulfo-NHS-biotin for 1h. (C) Ntn1 gene deletion strategy. (D) Whole-mount P12 retinas stained with the indicated antibodies. A: Artery, V: Vein
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Simmonds M et al (2025) Anti-VEGF drugs compared with laser photocoagulation for the treatment of proliferative diabetic retinopathy: a systematic review and individual participant data meta-analysis. Health Technol Assess 1–75. 10.3310/MJYP 657810.3310/MJYP 6578 PMC 1199524240186529 · doi ↗ · pubmed ↗
- 2Jiang G, Han X, Qiao K, Liu S (2022) Therapeutic effect of intravitreal anti-VEGF drugs on retinal neovascularization in diabetic retinopathy. Minerva Med. 10.23736/S 0026-4806.22.07943-510.23736/S 0026-4806.22.07943-535838253 · doi ↗ · pubmed ↗