Phobic responses of bull ants (Myrmecia midas) to odours introduced on their foraging route
Venkata Manvitha Kambala, Yousef Ahmed, Jasmine Lee, Anwin Jose, Sahera Syed Nadir, B. C. Priyanka, Ali Gabir, Yingdie Sun, Ken Cheng, Sudhakar Deeti

TL;DR
Bull ants avoid unfamiliar odors on their foraging routes, showing phobic responses that affect their navigation.
Contribution
This study reveals neophobic responses in bull ants to novel odors, demonstrating how chemical cues influence their vertical navigation.
Findings
Bull ants showed phobic responses to all five odors introduced on their foraging routes.
Tea-tree and Lavender odors caused the strongest detours and scanning behaviors.
Repeated exposure to Lavender reduced aversive responses, suggesting neophobia rather than inherent aversion.
Abstract
Goal-oriented learning and navigation is well known in eusocial insects. The solitary foraging of nocturnal bull ants Myrmecia midas in their visually complex environment relies on path integration and landmark learning. While this species seems to be ‘sensitive’ to handling and reacts to visual changes in their surroundings, not much is known about how added olfactory stimuli impact their route navigation on a vertical surface. In the current study, we added one of five different invisible odours on the trees on which foragers normally forage. We found that the bull ants showed phobic responses to all the odours. The Tea-tree and Lavender odours showed the strongest impact on the bull ants’ navigation by causing detours, U-turns, and avoidance of the sensory stimuli, with the ants meandering more and scanning more frequently. The odours of Olive oil, Flax-seed oil, and Eucalyptus oil…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
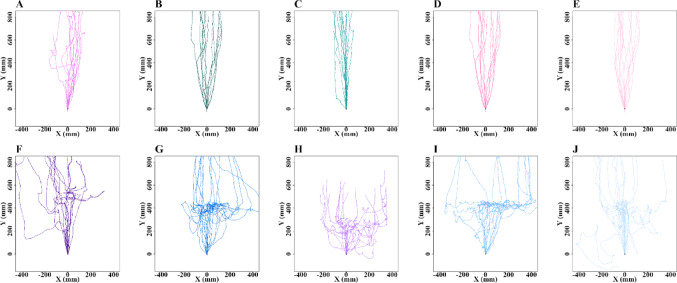 Figure 1
Figure 1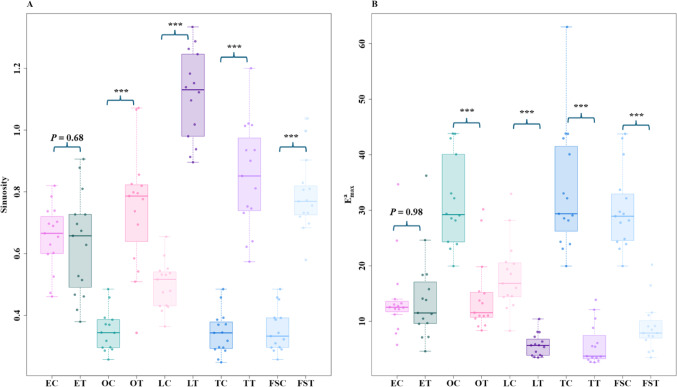 Figure 2
Figure 2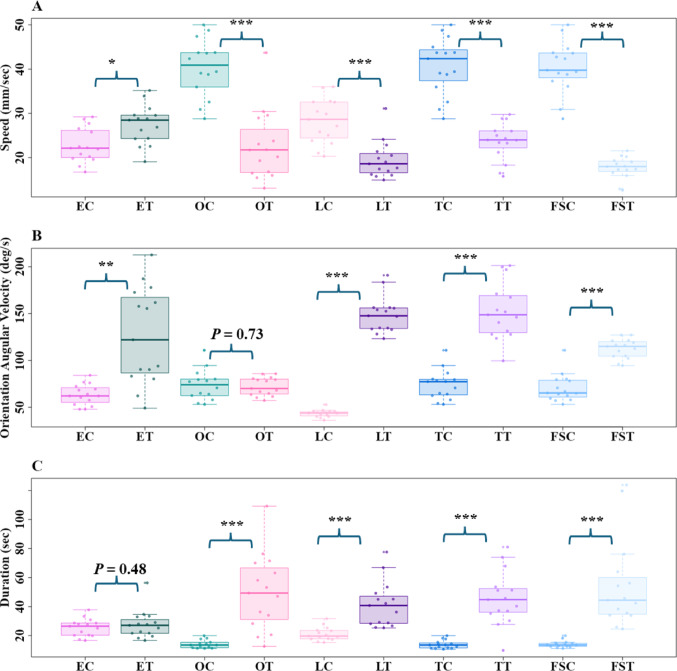 Figure 3
Figure 3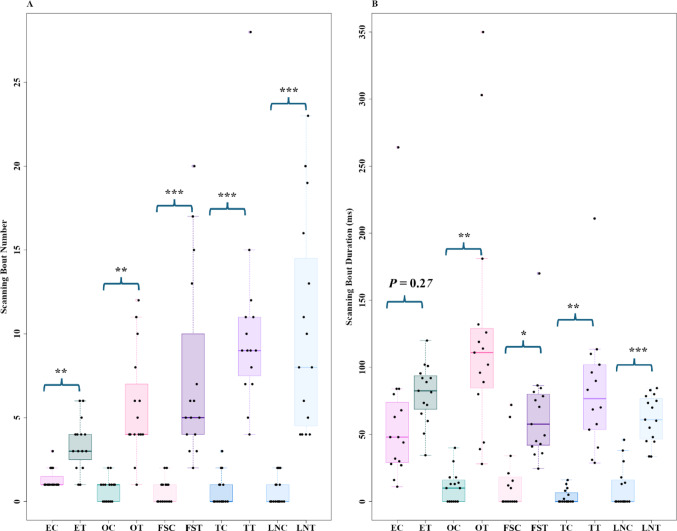 Figure 4
Figure 4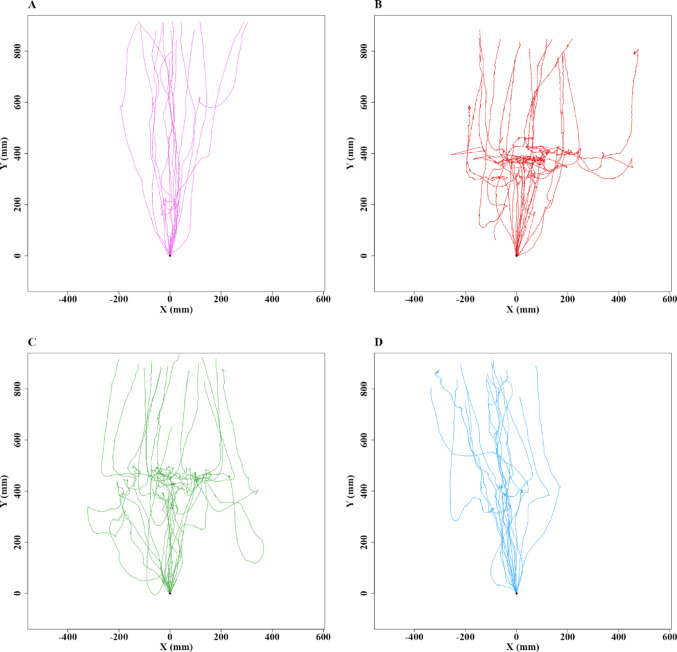 Figure 5
Figure 5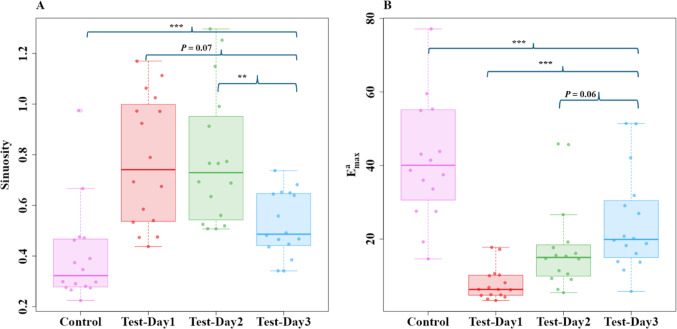 Figure 6
Figure 6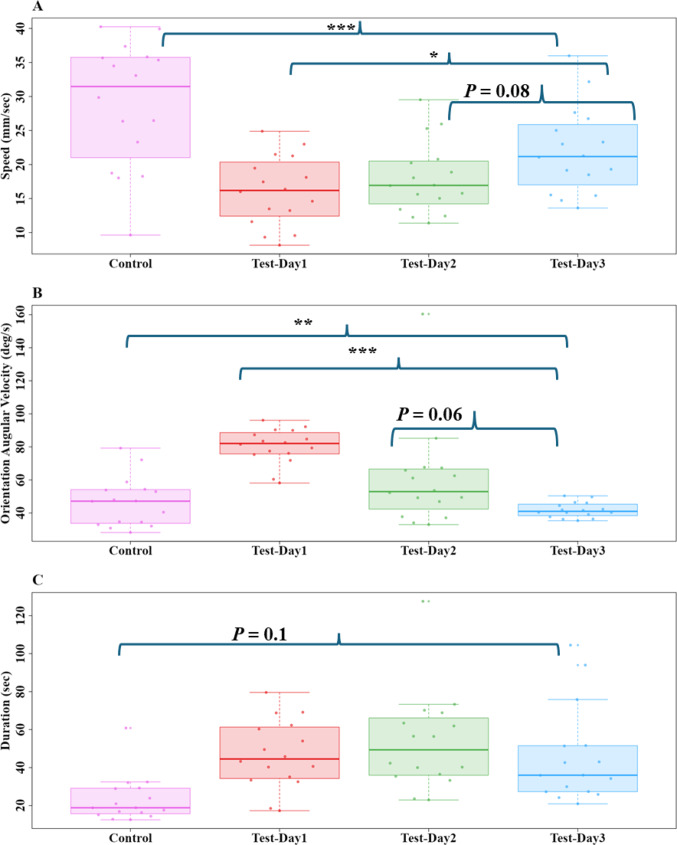 Figure 7
Figure 7- —Macquarie University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurobiology and Insect Physiology Research · Insect and Arachnid Ecology and Behavior · Animal Behavior and Reproduction
Introduction
Insects possess small brains, yet exhibit sophisticated and complex behaviours, demonstrating that efficient neural architectures and task-specific specializations can enable remarkable problem-solving and adaptability. Insects navigate efficiently from their home to their destination using a navigational tool kit consisting of multiple strategies (Buehlmann et al. 2020; Wehner 2020). Ant species that conduct group-based foraging rely on conspecific cues, typically pheromones, to navigate (Knaden and Graham 2016). A considerable number of ant species, however, forage solitarily, whereby each individual ant must complete its own search for food and return to its nest. These solitary foragers are adept at navigating on the ground, on trees, or underground. Bull ants, for example, are skilled in recognising their environments, the route to a foraging tree to locate food sources, and the way back to their nest (Deeti et al. 2023a; Narendra et al. 2013). Navigating ants use multiple strategies (Wehner 2008, 2020), including path integration (PI) (Heinze et al. 2018) and learned panorama cues (Narendra et al. 2013; Zeil 2023). Visual panoramas are thought to be learned by performing learning walks, widely observed behaviours in desert ant genera such as Cataglyphis (Fleischmann et al. 2016; Wehner et al. 2004), Ocymyrmex (Müller and Wehner 2010), and Melophorus (Deeti and Cheng 2021; Deeti et al. 2024a, b; reviews Freas et al. 2019; Zeil and Fleischmann 2019). Learning walks are not exclusive to arid environments, as they are also exhibited by non-desert ant species such as Myrmecia bull ants (Jayatilaka et al. 2018) and Formica wood ants (Nicholson et al. 1999). Ants are also known to use olfactory cues for locating food sources (Wolf and Wehner 2000) and their nest (Steck et al. 2009) and marking trails to guide nestmates to resources (Buehlmann et al. 2020; Knaden and Graham 2016). The prevalence of these behaviours in diverse ant species highlights the importance of olfactory cues in navigation, foraging, and place learning. However, the importance of olfactory cues on bull ants’ vertical navigation is largely unknown.
The bull ant species Myrmecia midas inhabits underground structured nests and journeys up surrounding trees to forage for food, then successfully descending down the tree to return to the nest (Islam et al. 2021, 2022, 2023). Nocturnal members of this genus, including M. pyriformis and M. midas, have an added challenge: conducting these three-dimensional foraging trips during evening and morning twilight when low light levels make visual cues harder to detect. Ants are known to use landmarks to navigate, and they can learn to associate landmarks with food sources (Collett and Collett 2002a). M. midas navigates visually while descending its foraging tree (Freas et al. 2018). Occasionally, while descending the tree trunk, M. midas foragers scan by bringing their head and eyes to a near upright position while their body remains vertically oriented. This behaviour suggests that whilst descending, M. midas foragers use visual cues memorised while horizontal on the ground to steer to the correct side of the tree. Blocking the panoramic views surrounding the tree leads to disorientation in the descent. Ascending foragers climb up the tree trunk to the canopy, but have not been observed to make these scans (personal observations), suggesting that they may attend to different cues, possibly ‘vertical views’ of the canopy, or gravitational cues to orient while climbing. Visual changes, such as movement or sudden appearance or disappearance of surrounding landmarks, can prompt these ants to react because bull ants, like many other ants, rely heavily on their vision to navigate their environment and find food (Collett et al. 2006; Islam et al. 2021, 2022, 2023). Ants have compound eyes that allow them to detect even the slightest movements and changes to their surroundings (Narendra et al. 2017; Warrant and Dacke 2010, 2011). Our account, however, concerns reactions to stimuli in another modality: olfaction.
Novel odours on an animal’s foraging path might generate a phobic response. Neophobia leads to an avoidance of unfamiliar stimuli, serving as a protection mechanism from possible threats heralded by the novel stimuli. The classification of novel and unfamiliar applies to anything that an organism has not experienced in the context in which the stimulus appears (Greenberg and Mettke-Hofmann 2001). Novelty can be found in food, circumstances, environments, fluids, and objects. Whilst neophobia varies across species, in birds, it increases with the degree of unfamiliarity (Miller et al. 2022). Neophobic responses may be reduced if an animal is exposed to a specific stimulus repeatedly (Greenberg and Mettke-Hofmann 2001). Bull ants M. midas exhibit phobic responses to changes in their learned routes that have been interpreted as neophobic responses (Islam et al. 2023). This includes detouring around new or unfamiliar objects, meandering more, and scanning the environment more frequently when presented with tactile and olfactory changes on the tree on which they travel to forage for food. Here we further explore their response to olfactory changes on their learned route with a range of chemical cues, examining changes in their travelling behaviour in more detail.
In this study, we explored how the ascending navigational behaviour of nocturnal bull ant Myrmecia midas foragers is influenced by introduced olfactory cues placed on their established foraging routes. Firstly, we made control or baseline observations on the travel characteristics of bull ants when travelling vertically up the eucalyptus tree trunk. Then on the following day, we examined possible phobic responses of the bull ants by applying a new olfactory cue on their foraging tree. The novel odours consisted of Eucalyptus oil, Olive oil, cold-pressed Flax-seed oil, Tea-tree oil, and Lavender oil. The direct route required walking over the added odours. We hypothesised that by introducing novel odours on their learned route, we would trigger detour or avoidance behaviour among the foragers. To assess the impact of the foreign odours, we measured path characteristics such as sinuosity, speed, angular velocity of orientation shifts, trajectory duration, the number of scanning bouts, and the scanning-bout durations, comparing control and test (odour-added) conditions. The behavioural changes will allow us to understand how phobic responses of the bull ants to the added chemical cues manifest in behaviour. A second experiment tested the second component of neophobia (Greenberg and Mettke-Hofmann 2001), that the aversive responses should diminish with repeated exposure. In contrast, a stimulus that is inherently aversive will not induce reduced aversive responses with repeated exposure. We took one of the most-avoided chemicals and presented it to the same ants over 3 days.
Materials and methods
Study location
During the Australian autumn months from March to April 2024 (Experiment 1), and March to April 2025 (Experiment 2) we conducted odour-response studies on nocturnal bull ant Myrmecia midas colonies at the Macquarie University campus, North Ryde, Sydney, Australia (33°46′18″ S, 151°06′30″ E). These bull ants typically construct their ground-nesting colonies near the base of eucalyptus trees, within a range of about 30 cm, to gain elevation and safeguard against floods (Deeti et al. 2024c). From our observations, we noticed that ants begin foraging vertically on the eucalyptus trees during the evening twilight, immediately after sunset, and their activity peaks within 20 to 30 min. They then return to the nest during the morning twilight. These experiments were conducted exclusively on trees near the nest, specifically focusing on foragers on the nest tree. Although ethical approval was not required for researching and collecting ants in Australia, we ensured that our experiments did not harm individual ants or their colonies.
Experimental setup
In Experiment 1 (autumn 2024), we used five different trees and nests. In Experiment 2 (autumn 2025), we used only one nest. Before conducting the experiments, we observed the individual trees for a week to understand the ants’ well-maintained foraging corridor while vertically travelling up on the tree, and then we marked their preferred foraging corridor. All the foragers had followed a 40-cm-wide corridor. Before conducting any experiments, we labelled the four corners of the recording area, situated 50 cm above the ground, to mark a 1-m-high and 70-cm-wide area where we could record the paths of individual foragers during their upward foraging trips, which we call ascending navigation, distinct from their ground-level movements. We used a Sony Handy camera (FDR-AX700) to record the paths of vertically ascending navigators. The camera was set on a tripod positioned 0.6 m above the ground, recording footage at 25 frames per second. The camera’s field of view covered the focal area of the tree surface, recording with a high resolution of 3860 by 2160 pixels. The tripod and camera were in place for a week before experimentation began, to habituate ants to the presence of the set up. During experiments, the camera was switched on before the first forager entered the recording area and recorded continuously for 40 min in Experiment 1 and for 2 h in Experiment 2.
Procedure
Experiment 1
In Experiment 1, we used five different odours for investigation. These included Eucalyptus oil (Coles^®^, Eucalyptus Oil 1 mL/mL), representing a local odour of the experimental tree surface, Olive oil (Remano^®^, oleic acid 0.75 mL/mL), which is a source rich in oleic acid, and cold-pressed Flax-seed oil (Coles^®^, linoleic acid, 0.63 mL/mL ), a source rich in linoleic acid, cold-pressed to preserve this fragile omega-6 fatty acid. Both these fatty acids have been recognised as odours that ants associate with death (Sun et al. 2018). We also included Tea tree oil (Thursday Plantation^®^, Melaleuca Oil 1 mL/mL) and Lavender oil (Hemani^®^, Lavender oil 1 mL/mL) as foreign odours in our study. All these odours were novel on the foraging route. Each of these odours was tested only once on the nest tree of a single nest, different nests for different odours. The experiment at each nest lasted two days, with Control observations on day 1 and Test-day observations on day 2. We did not capture or paint any ant because this process is aversive to this species and will generate avoidance responses whether any odour is added or not (Lionetti et al. 2023).
Control day. On the control recording day, we set up the camera 5 min before sunset. Once a forager reached the base of the foraging tree, the observer turned on the camera to record. The recording continued for 40 min or until the sample size reached 15 ants. We did not apply any scents on the control day.
Test day. On the test day, 5 min before sunset, we placed a scent in the middle of the recording area along the foraging path. We used 2 ml of oil on each tree, a horizontal strip 50 cm by 5 cm applied with a cotton gauze. Once the scent was applied, the observer began recording with the camera as soon as a forager ant reached the base of the foraging tree. The recording continued for the next 40 min or until 15 ants had been observed.
Experiment 2
To study how ants respond to a new smell over multiple exposures, we needed to test the same ants several times. On the first day, we collected 20 foraging ants as they walked through the recording area. Each ant was marked with a specific paint colour on its abdomen for identification and then released at the nest entrance. We then gave them three days to settle and recover from any stress caused by being captured and painted.
Control Day. After the three-day settling period, we recorded the activity of the painted ants on the fourth day as our control. We set up the camera five minutes before sunset. When a forager ant reached the base of the foraging tree, the observer started recording. The camera recorded continuously for two hours. No scent was applied on the control day.
Test days. On the fifth day, we introduced a foreign smell using lavender oil (Hemani^®^, 1 mL/mL), chosen because the ants showed the most aversive responding to it in Experiment 1. Five minutes before sunset, we applied 2 mL of lavender oil to the tree using a cotton gauze in a 50 cm by 5 cm strip along the foraging path every day. As soon as a forager reached the base of the tree, the observer started recording. The camera ran for two hours. We repeated this test for the next two days, recording a total of three trials with the lavender across three consecutive days. If the aversion to the odour is neophobia, we predict that the aversive responses would diminish over the three trials. If the odour is inherently aversive, avoidance behaviours are expected to continue unabated over the three trials.
Tracking
We used the animal tracking program DLTdv8 (version 8.2.9) in MATLAB (2022B) to extract frame-by-frame coordinates of the head and thorax—specifically the tip of the head and the middle of the thorax — for each ant in every video obtained during our recording of control and test days. These extracted frame-by-frame coordinates served as the basis for all subsequent analyses of the foragers’ movements and behaviour.
Data analysis
To understand how quickly and far the ants moved, we calculated speed. Speed refers to the magnitude of an ant’s velocity and was calculated as the average over the entire videotaped trajectory for each ant. Stopping durations, defined as periods where velocity fell below 0.01 mm/s (natural ant movement), were excluded from both distance and time calculations to accurately determine the average speed. We also calculated the orientation angular velocity by measuring the rate of change of an ant’s orientation direction over time. When walking, the orientation direction of these ants typically oscillates continuously, with the head swinging left and right. Orientation direction in any frame was determined as the direction of the straight line passing from the thorax coordinates through the head coordinates. Orientation angular velocity was calculated by dividing the change in orientation angle by the corresponding change in time, providing a measure of how quickly the ant was altering its head direction. To understand path characteristics, we used two indices of straightness: sinuosity, and E^a^max, each of which relates to the directness of navigation away from the nest. Sinuosity assesses the degree of waviness or curvature in an ant’s path as it navigates away from the nest (Batschelet 1981; Deeti et al. 2023a). A higher sinuosity value indicates a more convoluted path, while a lower value indicates a straighter path. Sinuosity is an estimate of the tortuosity in a path, calculated as \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:S=2{\left[p\left(\frac{1+c}{1-c}+{b}^{2}\right)\right]}^{-0.5}$$\end{document} , where \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:p$$\end{document} is the mean step length, \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:c$$\end{document} is the mean cosine of turning angles and \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:b$$\end{document} is the coefficient of variation of the step length. A trajectory step is the movement between the positions of the animal (thorax positions) recorded at consecutive video frames. Accordingly, step lengths are the Euclidean distances between consecutive points along a path, and turning angle refers to the change in direction between two consecutive steps (Benhamou 2004; Deeti and Cheng 2024d) The maximum expected displacement of a path, E^a^max = β/(1–β), where β is the mean cosine of turning angles, is a dimensionless value expressed as a function of number of steps, and is consistent with the intuitive meaning of straightness (Cheung et al. 2007; Deeti and Cheng 2025). Larger maximum expected displacement values indicate straighter paths, hence greater displacement, while a smaller value suggests more localised or constrained movement.
During the outbound navigation, ants frequently displayed a series of stereotypical successive fixations in different head directions by stopping and rotating on the spot at one location, known as a “scanning bout” (Deeti et al. 2023a, b). During each ant’s control and test runs, we extracted the number of scanning bouts and the scanning-bout durations, from the start of a scanning bout until the ant started walking again.
Statistical analysis
Experiment 1, conducted at the five different nests, was analysed using t-tests, one test for each nest. A repeated-measures ANOVA was conducted for Experiment 2 to determine if there are statistically significant differences in the dependent variables measures across the control and test conditions (Control, Test-Day1, Test-Day2, Test-Day3). For Experiment 2, a priori Helmert contrasts were used to assess recovery patterns across test days. Specifically, the independent contrasts compared: (1) the control day versus all test days combined, (2) Test-Day1 versus Test-Day2 and 3, and (3) Test-Day2 versus Test-Day3.
Results
Experiment 1
After emerging from the nest during the evening twilight, all the nest-tree foragers followed a stereotypical vertical path up the eucalyptus tree trunk and continued along the same foraging corridor every day. We conducted observations on these experienced foragers for two consecutive nights on their nest trees for control (N = 15) and odour-test (N = 15) purposes. The presence of odours on their foraging corridor resulted in detours for foraging (Fig. 1). Specifically, with Eucalyptus oil, 46.4% of ants walked over the odour, while 53.6% took detours to avoid it. With Olive oil and Flax-seed oil (sources of oleic acid and linoleic acid, respectively), 35% and 40% of foragers, respectively, walked over the odour, while 65% and 60% took detours. With Tea tree oil and Lavender oil, none of the foragers walked over the odour, with 26% and 22% of foragers, respectively, taking a U-turn and returning to the nest, while 74% and 78% took detours away from the odour line. These results suggest that foragers often exhibit a phobic response, with nearly half of them avoiding the familiar Eucalyptus oil, more avoiding oils containing oleic and linoleic acid, and complete avoidance of Tea tree and Lavender oils.
In path characteristics, ants exhibited a more curved and meandering trajectory in the odour-change conditions compared to the control conditions. We found differences across conditions in each of our two measures of path meander. Firstly, sinuosity increased significantly in the odour-change conditions (Olive oil control vs. Olive oil test: t = − 7.4, df = 17.4, P ≤ 0.0001; Flax-seed oil control vs. Flax-seed oil test: t = − 14.1, df = 19.4, P ≤ 0.0001; Tea tree oil control vs. Tea tree oil test: t = − 10.4, df = 18.2, P ≤ 0.0001; Lavender oil control vs. Lavender oil test: t = − 12.2, df = 22.8, P ≤ 0.00001) except for Eucalyptus oil (Eucalyptus oil control vs. Eucalyptus oil test: t = 0.4, df = 23.8, P = 0.6) (Fig. 2A). Secondly, E ^a^ max was lower in the odour-change conditions, with the ants having a smaller amount of displacement per unit length travelled compared to the control conditions. The t-tests showed a significant difference in test conditions compared to the control conditions in all comparisons except Eucalyptus oil (Olive oil control vs. Olive oil test: t = 6.1, df = 25.2, P ≤ 0.0001; Flax-seed oil control vs. Flax-seed oil test: t = 7.2, df = 16.8, P ≤ 0.0001; Tea tree oil control vs. Tea tree oil test: t = 9.08, df = 16.9, P ≤ 0.0001; Lavender oil control vs. Lavender oil test: t = 9.6, df = 23.2, P ≤ 0.0001; Eucalyptus oil control vs. Eucalyptus oil test: t = − 0.02, df = 27.5, P = 0.9) (Fig. 2B).
Odour changes had a noticeable impact on the speed, orientation angular velocity, and duration of the foragers. When the foragers sensed non-visual odour stimuli, they slowed down, frequently shifted their gaze in different directions, and stayed in the recording area longer compared to the respective controls (Fig. 3). In speed, the t-tests revealed significant differences in all comparisons (Fig. 3A), with the ants speeding up with Eucalyptus oil and slowing down with other odours (Eucalyptus oil control vs. Eucalyptus oil test: t = − 3.04, df = 27.6, P < 0.01; Olive oil control vs. Olive oil test: t = 6.5, df = 26.6, P ≤ 0.0001; Flax-seed oil control vs. Flax-seed oil test: t = 5.2, df = 27.01, P ≤ 0.0001; Tea tree oil control vs. Tea tree oil test: t = 8.5, df = 24.2, P ≤ 0.0001; Lavender oil control vs. Lavender oil test: t = 13.9, df = 19.06, P ≤ 0.00001). Changes in odour also increased the magnitude of orientation angular velocity in the foragers (Fig. 3B). The t-tests showed significant differences between all pairs in orientation angular velocity except for Olive oil (Eucalyptus oil control vs. Eucalyptus oil test: t = − 4.6, df = 15.2, P ≤ 0.001; Flax-seed oil control vs. Flax-seed oil test: t = − 20.8, df = 15.1, P ≤ 0.00001; Tea tree oil control vs. Tea tree oil test: t = − 8.9, df = 21.1, P ≤ 0.0001; Lavender oil control vs. Lavender oil test: t = − 9.06, df = 25.1, P ≤ 0.0001; Olive oil control vs. Olive oil test: t = 0.3, df = 20.9, P = 0.7). The amount of time spent in the recording area also increased, as foragers stopped just before the odour line and meandered around it (Fig. 3C). Consequently, trips took significantly more time with all odours except for Eucalyptus oil (Olive oil control vs. Olive oil test: t = − 5.3, df = 14.3, P ≤ 0.0001; Flax-seed oil control vs. Flax-seed oil test: t = − 4.8, df = 16.4, P ≤ 0.0001; Tea tree oil control vs. Tea tree oil test: t = − 6.6, df = 14.6, P ≤ 0.0001; Lavender oil control vs. Lavender oil test: t = − 5.1, df = 14.1, P < 0.0001; Eucalyptus oil control vs. Eucalyptus oil test: t = − 0.7, df = 23.4, P = 0.4).
Odour change increased scanning in foragers. In Control recordings, i.e. those without any odour change, the majority of the foragers performed a single scan across their entire recording area (Fig. 4A). In contrast, on odour trials, where ants experienced a non-visual odour change, foragers scanned at least twice, with a maximum of 28 (Fig. 4A). The t-tests revealed significant differences in the number of scanning bouts in all comparisons (Eucalyptus oil control vs. Eucalyptus oil test: t = − 4.7, df = 18.3, P ≤ 0.001; Olive oil control vs. Olive oil test: t = − 4.8, df = 15.1, P ≤ 0.0001; Flax-seed oil control vs. Flax-seed oil test: t = − 4.1, df = 15.9, P ≤ 0.0001; Tea tree oil control vs. Tea tree oil test: t = − 5.5, df = 14.9, P ≤ 0.0001; Lavender oil control vs. Lavender oil test: t = − 4.7, df = 15.9, P ≤ 0.0001). The duration of scanning bouts also increased significantly with odour changes (Fig. 4B), in all comparisons except for Eucalyptus oil (Olive oil control vs. Olive oil test: t = − 4.2, df = 14.4, P ≤ 0.001; Flax-seed oil control vs. Flax-seed oil test: t = − 2.7, df = 27.87, P < 0.01; Tea tree oil control vs. Tea tree oil test: t = − 4.8, df = 14.5, P < 0.001; Lavender oil control vs. Lavender oil test: t = − 5.4, df = 24.1, P ≤ 0.0001; Eucalyptus oil control vs. Eucalyptus oil test: t = − 1.1, df = 17.60, P = 0.2).
Experiment 2
To understand whether the detouring mechanism is triggered by neophobia or inherent aversion to an odour, we applied Lavender oil, which all the ants avoided on the Test day in Experiment 1, and recorded individual ant behaviour over three days. On the control day (N = 16), all ants went straight up the tree. On the first test day (Test-Day1) (N = 16), after applying the lavender oil odour, only one ant walked over the odour, one took a U-turn, and the rest detoured to avoid the odour. On the second test day (Test-Day2) (N = 16), five ants walked over the odour. By the third test day (Test-Day3) (N = 16), only two ants detoured, while the rest walked over the odour (Fig. 5).
To understand the impact of the odour on ants’ navigational performance, we compared the path characteristics of foragers on the Control and Test-Day1 to 3. Firstly, the sinuosity was observed to be low in the Control condition. It increased significantly on Test-Day1, decreased on Test-Day2, and showed variation in magnitude on Test-Day3. The repeated-measures ANOVA model revealed a significant difference (F3,56 = 4.4, P < 0.006) across the conditions (Fig. 6). A priori Helmert Contrast-1 (Z_3,56_ = 6.1, P = 0.0009) showed significant differences between the control and test conditions. Contrast-2 revealed a trend (Z_3,56_ = 1.7, P = 0.07), while contrast-3 revealed significant differences between the Test-Day2 and Day3 (Contrast-3: Z_3,56_ = 3.26, P = 0.002). These results show significant recovery by Test-Day3.
Secondly, E ^a^ max as observed to be lower in the Test-Day1 and it gradually increased over the next two Test Days. The repeated-measures ANOVA revealed a significant difference (F3,56 = 7.8, P ≤ 0.0001) across the days (Fig. 6). The first two a priori Helmert tests (Contrast-1: Z_3,56_ = 7.1, P ≤ 0.0001; Contrast-2: Z_3,56_ = 5.6, P ≤ 0.0008) showed significant differences between the contrasted conditions, while Contrast-3 revealed no significant differences between the Test-Day2 and Day3 (Contrast-3: Z_3,56_ = 1.85, P = 0.06). These results indicate that the experimental odour applied on their foraging route had a significant impact on the characteristics of the ants’ paths and that the ants recovered in subsequent trips.
The odour change had a noticeable impact on the speed, orientation angular velocity, and duration of the foragers from the Control Day to Test-Day3. The mean speed of foragers decreased on Test-Day1 and gradually increased over the next two Test Days. The repeated-measures ANOVA revealed a significant difference (F3,56 = 3.8, P < 0.03) across the conditions (Fig. 7). A priori Helmert Contrast-1 (Z_3,56_ = 4.4, P ≤ 0.0006) showed significant differences between the control and test conditions. Contrast-2 revealed a borderline effect (Z_3,56_ = 2.2, P = 0.04), while the other Helmert contrast revealed no significant differences between Test-Day2 and Test-Day3 (Contrast-3: Z_3,56_ = 1.7, P = 0.08).
The orientation angular velocity of foragers increased on Test-Day1 and gradually decreased over the next two Test Days (Fig. 7). The repeated-measures ANOVA revealed a significant difference (F3,56 = 10.2, P ≤ 0.00001) across the days. A priori Helmert test Contrast-1 (Z_3,56_ = 3.1, P = 0.003), Contrast-2 (Z_3,56_ = 6.2, P ≤ 0.00006), and Contrast-3 (Z_3,56_ = 3.03, P ≤ 0.003) all showed significant differences between contrasted conditions. The duration of time spent in the recording area also increased, as foragers stopped just before the odour line and meandered around it. However, the repeated measures ANOVA revealed no significant differences (F3, 56 = 2.1, P = 0.1) across the days (Fig. 7). Overall, these results indicate that the odour treatment significantly influenced the foragers’ speed and orientation angular velocity.
Discussion
This study examined whether manipulating bull ants’ natural environment with odorific oils will elicit abnormal behaviour. When exposed to foreign odours along their regular foraging path on the tree trunk, the ants immediately showcased avoidance behaviours, consistent with a previous study (Islam et al. 2023). The level of avoidance varied depending on the odour, with almost complete avoidance for Tea tree and Lavender oils, moderate avoidance for oils containing oleic and linoleic acid, and partial avoidance for Eucalyptus oil, whose odour is familiar to them, albeit much stronger than usual. The results suggest that ants are most habituated to Eucalyptus oil, showcasing the least disturbance in their daily routine. None of the foragers walked over the Tea tree oil and Lavender oil in Experiment 1; on average 76% detoured around the odour corridor, exhibiting the most avoidance. In the presence of novel odours, the ants’ paths became more curved and meandering, as indicated by increased sinuosity and lower displacement efficiency compared to the control conditions. When encountering novel odours, the ants slowed down their speed, exhibited increased head-turning angular velocity (except for Olive oil), and spent more time in the recording area, suggesting they were searching for alternative routes. In the presence of odour changes, the ants performed multiple scanning bouts, with some individuals scanning up to 28 times, in contrast to a single scan in the control conditions. The duration of scanning bouts also increased significantly with odour changes. In summary, the study demonstrates that foraging ants exhibit a suite of phobic responses when exposed to unfamiliar odours along their regular foraging paths. This suite manifests as avoidance behaviour, path deviations, reduced speed, increased scanning, and altered movement patterns as they attempt to find alternative routes around the novel odour stimuli. In a second experiment, repeated exposure to the Lavender-oil odour on the foraging tree led to marked decreases in all the phobic reactions over trials.
Our interpretation is that this suite of phobic responses stem from neophobia in the ants, a fear of new kinds of stimuli. Animals are known to exhibit neophobic responses to changes in their learned routes by detouring around new stimuli or unfamiliar objects, meandering more, and scanning the environment more frequently (Mettke-Hofmann 2022; Islam et al. 2023). The current study species, the nocturnal bull ant M. midas, showed an avoidance response when they were captured and released back at the same location after a minute (Lionetti et al. 2023), as the location became aversive to them. Although other interpretations are possible, such as a reaction to new stimuli without aversion, we interpret the avoidance response and the other behaviours as indicating fear of new kinds of stimuli. The prevalence of avoiding the added stimuli altogether and even turning back to return home tips our interpretation in favour of neophobia. Another study on this species’ reactions to changes to the tactile or olfactory environment (Islam et al. 2023) also found avoidance responses in the bull ants, interpreted as neophobia. Here we found the extent of neophobic response to particular stimuli varied. For known odours such as Eucalyptus, Flax-seed oil, and Olive oil, the ants showed a moderate phobic response. However, the foreign Tea tree and Lavender oils induced a stronger phobic response. Almost none of the foragers walked over these odours; instead, some took U-turns to returned to their nest and the ants’ paths were found to be more curved and less straight, with frequent stops and scans, likely due to their search for an alternative way to reach the tree canopy by taking detours from the original path. As a result, it took them longer to reach the final heading direction. The interpretation of neophobia, as opposed to an inherent aversion to the odours, is supported by the results of Experiment 2. Repeated exposure to an odour that was completely avoided on first exposure led to decreases in all phobic responses, consistent with one of the characteristics of neophobia (Greenberg and Mettke-Hofmann 2001). We interpret such decreases tentatively as habituation, while noting that to support an interpretation of habituation (Rankin et al. 2009), more response characteristics should be examined.
We conjecture that the longer working life of this species and the smaller number of foragers in the colony are functional reasons for exhibiting a fear response to novel stimuli (Lionetti et al. 2023). A longer working life and a smaller work force mean that each worker is more valuable to the colony. A more valuable life should be more cautious about changes in the environment, which might signal danger. Mechanistically, the ants likely perceived the non-visual unfamiliar odours as potentially harmful, causing them to avoid walking over them. Future studies should explore how these olfactory changes affect their foraging efficiency and behaviour over longer periods.
Our experiments found that non-visual odour changes on their familiar route on the foraging tree induced detour behaviour in the bull ants to avoid the odour line. This species improves over trials in detouring around a physical barrier (Islam et al. 2023). Experiment 2 here showed, however, that with repeated exposure to a foreign odour, the ants mostly abandoned the detour strategy and walked right over the odour two trials after initial exposure (on trial 3). They did not need to learn to detour better.
The cases of the oils containing key unsaturated fatty acids, Olive oil and Flax-seed oil, demand more consideration. Fatty acids are known to play a crucial role in the interactions between ants and their environment, affecting their behaviour and ecological dynamics. Many ant species dump rubbish outside the nest in the field (e.g. Pogonomyrmex barbatus, Gordon 1983). In laboratory experiments, oleic and linoleic acids have been shown to trigger removal behaviour in ants (review: Sun et al. 2018). Oleic acid, a monounsaturated fatty acid, has been tested on Pogonomyrmex badius and Solenopsis saevissima (Wilson et al. 1958), Myrmica vindex (Haskins and Haskins 1974), Myrmica rubra (Diez et al. 2013), and Solenopsis invicta (Qiu et al. 2015). Linoleic acid, an omega-6 polyunsaturated fatty acid, has been tested on Myrmica rubra (Diez et al. 2013) and Solenopsis invicta (Qiu et al. 2015). In the current experiment, Olive and Flax-seed oil (sources of the aforementioned death-associated odours) showed a moderate impact on ant navigation; some of the ants detoured around while others walked over the odour line. These fatty acids had less effect here than did the strange odours of Tea tree and Lavender oils, suggesting novelty of chemical cues as a primary driver of neophobia rather than the fatty acids associated with death. Why did oils containing fatty acids shown in previous research to be associated with corpses not have a stronger effect? The context in which these fatty acids appear comes to mind as a possible reason. These food oils have all kinds of other chemicals, possibly containing odours associated with food plants. That might make the effects of oleic and linoleic acid not as strong in causing an avoidance reaction. The context of a foraging tree might also matter, as it is not a place where rubbish is found, and the ants probably have not ever encountered any dead organisms on the tree trunk during their previous foraging expeditions. In a foraging context, these odours might even have attractive properties, indicating the availability of dead prey (Buehlmann et al. 2014), although a corpse’s odour would likely not be as strong as the oils applied in Experiment 1. In vertebrate animals (Bouton 2004; Bouton et al. 1999) and in insects (Cheng 2005; Colborn et al. 1999; Collett et al. 2002b; Pahl et al. 2007), contextual cues modulate learning, memory, and behaviour. Further studies, however, are warranted to confirm this hypothesis, including experiments that apply pure oleic acid and linoleic acid on the foraging route, the chemicals used in the studies reviewed in this paragraph.
Conclusion
In summary, this study provides important insights into the neophobic responses displayed by the bull ant Myrmecia midas when encountering different odours along their standard foraging routes. The results demonstrate that when exposed to unfamiliar odours, these ants exhibit a range of behaviours: avoidance of the chemicals with path deviations, scanning, and meandering movement patterns. The responses varied in intensity, with complete avoidance of foreign odours like tea tree and lavender oils, moderate avoidance of odours containing oleic and linoleic acids (which have been linked to cues associated with death), and less but still moderate avoidance for the slightly familiar eucalyptus oil. In the presence of novel odours, the ants’ paths became more curved and meandering, with increased sinuosity and lower displacement efficiency. They often decreased their speed and increased head-turning angular velocity, likely searching for alternative routes around the odour stimuli. The neophobic responses to a novel odour diminished over repeated trials with the same odour. The observed neophobic responses highlight the importance of olfactory cues in shaping the ants’ behaviour and decision making during their navigation. This study highlights the link between sensory perception and behavioural plasticity which enables these insects to thrive in their environments. Future research could explore the long-term effects of these odour changes on foraging efficiency.
Fig. 1. Trajectories of ants in Experiment 1 in control and test conditions with various odours added. A–J show the Control (N = 15) and Test (N = 15) paths of the bull ants during vertical navigation. A–E represent the control paths before Eucalyptus oil, Olive oil, cold-pressed Flax-seed oil, Tea tree oil, and Lavender oil, respectively, were added. F–J depict the test paths after Eucalyptus oil, Olive oil, cold-pressed Flax-seed oil, Tea tree oil, and Lavender oil, respectively, were added. Each trajectory plot represents the movement path of an individual ant. The y-axis points upwards. The coordinates (0,0) represent the starting point of each ant trajectory on the videorecording
Fig. 2. Path characteristics of bull ants in Experiment 1 during the control and test odour trials. Shown are the A Sinuosity and B E^a^max under control and test-odour observations. The paired Box plots represent the median (line inside the box), interquartile range (box), and extreme values excluding outliers (whiskers). Individual data points are shown as dots. Each point represents a single trajectory measure. EC, OC, FSC, TC, LNC represent Eucalyptus oil, Olive oil, Flax-seed oil, Tea tree oil, and Lavender oil controls whereas ET, OT, FST, TT, LNT represent Eucalyptus oil, Olive oil, Flax-seed oil, Tea tree oil, and Lavender oil tests respectively. The Pairwise comparisons between control and test groups shows the significance in path characteristics for several odours, as indicated by the annotations on the plots. *** denotes P ≤ 0.0001
Fig. 3. Comparison of Speed, Orientation Angular Velocity and Duration of the entire path of bull ants in Experiment 1 during the control and test odour trials. Box plots represent the median (line inside the box), interquartile range (box), and extreme values excluding outliers (whiskers). Individual data points are shown as dots. Each point represents a single trajectory measure. EC, OC, FSC, TC, LNC represent Eucalyptus oil, Olive oil, Flax-seed oil, Tea tree oil, and Lavender oil controls and whereas ET, OT, FST, TT, LNT represent Eucalyptus oil, Olive oil, Flax-seed oil, Tea tree oil, and Lavender oil tests, respectively. The pairwise comparisons between control and test groups show the significance in path characteristics for several odours, as indicated by the annotations on the plots. *** denotes P ≤ 0.0001, ** denotes P ≤ 0.001, and * denotes P ≤ 0.01
.
Fig. 4. Number of scanning bouts (A) and the duration of scanning bouts (B) of the bull ants in Experiment 1 during the control and test observations. The paired Box plots represent the median (line inside the box), interquartile range (box), and extreme values excluding outliers (whiskers). Individual data points are shown as dots. Each point represents a single trajectory measure. EC, OC, FSC, TC, LNC represent Eucalyptus oil, Olive oil, Flax-seed oil, Tea tree oil, and Lavender oil controls whereas ET, OT, FST, TT, LNT represent Eucalyptus oil, Olive oil, Flax-seed oil, Tea tree oil, and Lavender oil tests, respectively. The pairwise comparisons between control and test groups show the significance in path characteristics for several odours, as indicated by the annotations on the plots. *** denotes P ≤ 0.0001, ** denotes P ≤ 0.001, and * denotes P ≤ 0.01
Fig. 5. Trajectories of ants in Experiment 2 in control and test conditions with presence of odour. A–D show the Control and Test paths of the bull ants during vertical navigation. A represents the Control paths before Lavender oil was added. B–D depict the paths from Test-Day1 to 3 after Lavender oil was added. Each trajectory plot represents the movement path of an individual ant. The y-axis points upwards. The coordinates (0,0) represent the starting point of each trajectory on the videorecording
Fig. 6. Path characteristics of bull ants in Experiment 2 during the control and test odour trials. Shown are the A Sinuosity and B E^a^max on Control and Test Days. Box plots represent the median (line inside the box), interquartile range (box), and extreme values excluding outliers (whiskers). Individual data points are shown as dots. Each point represents a single trajectory measure. The independent contrasts compared: (1) the control day vs. all test days combined, (2) Test-Day1 vs. Test-Day2 and 3, and (3) Test-Day2 vs. Test-Day3. *** denotes P ≤ 0.0001, ** denotes P ≤ 0.001, and * denotes P ≤ 0.01
Fig. 7. Comparison of Speed, Orientation Angular Velocity and Duration of the entire path of bull ants in Experiment 2 during the control and test odour trials. Box plots (A–C) represent the median (line inside the box), interquartile range (box), and extreme values excluding outliers (whiskers) of the duration of the ant on the video. Individual data points are shown as dots. Each point represents a single trajectory measure. The independent contrasts compared: (1) the control day vs. all test days combined, (2) Test-Day1 vs. Test-Day2 and 3, and (3) Test-Day2 vs. Test-Day3. *** denotes P ≤ 0.0001, ** denotes P ≤ 0.001, and * denotes P ≤ 0.01
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Deeti S, Cheng K (2024 d) Desert ant (Melophorus bagoti) dumpers learn from experience to improve waste disposal and show spatial fidelity. Insects 15(10):814. Retrieved from https://www.mdpi.com/2075-4450/15/10/81410.3390/insects 15100814 PMC 1150899339452390 · doi ↗ · pubmed ↗
- 2Deeti S, Mc Lean DJ, Murray T, Cheng K (2024 b) Route learning and transport of resources during colony relocation in Australian desert ants. Learn Behav 1–13. 10.3758/s 13420-024-00652-110.3758/s 13420-024-00652-1PMC 1209254139438403 · doi ↗ · pubmed ↗