Morphological Ambiguity between Chambersiella Cobb, 1920, and Geraldius Sanwal, 1971: A Taxonomic Dilemma Solved through a Note from N. A. Cobb
Jonathan D. Eisenback, Paulo Vieira

TL;DR
This paper resolves a taxonomic confusion between two nearly identical nematode genera by reevaluating their distinguishing features and historical records.
Contribution
The paper proposes synonymizing Geraldius with Chambersiella based on new archival evidence and field observations.
Findings
Field surveys found no monodelphic specimens, contradicting the original diagnosis of Chambersiella.
Archival notes from N. A. Cobb support synonymizing Geraldius with Chambersiella.
Morphological evidence consistently aligns with the revised classification.
Abstract
The genera Chambersiella and Geraldius (Nematoda) are nearly morphologically identical, differing primarily in female ovary number: Chambersiella was described as monodelphic, while Geraldius was diagnosed as didelphic. This note reevaluates the validity of that distinction, incorporating original descriptions and a previously overlooked archival note from N. A. Cobb. Field observations failed to recover monodelphic specimens, even in type localities. We propose synonymizing Geraldius with Chambersiella, supported by Cobb’s archived observations and consistent morphological evidence.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
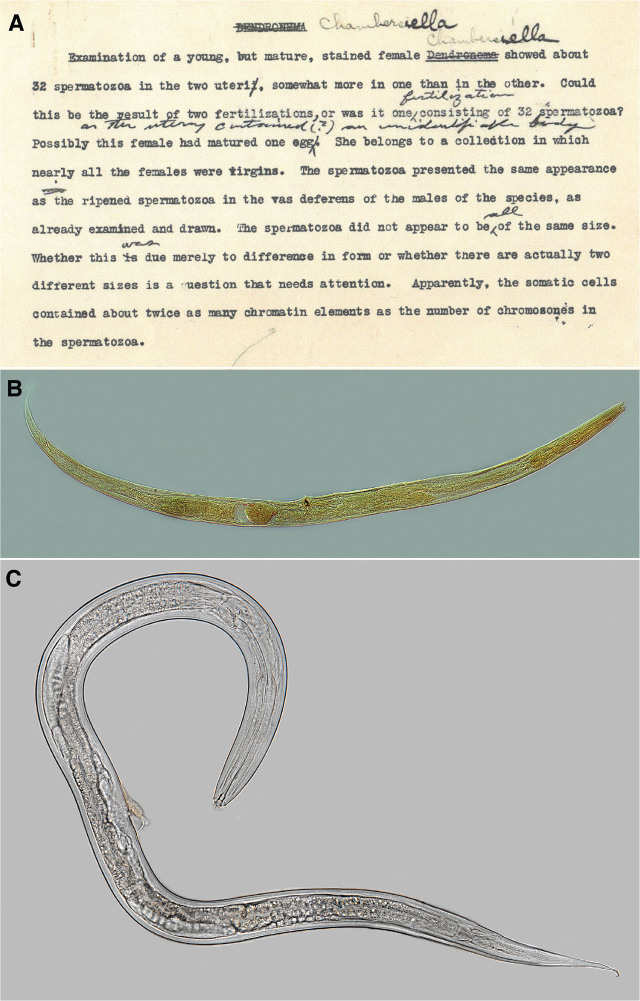 Figure 1
Figure 1|
|
|
|
|
|
|
|
|
|---|---|---|---|---|---|---|---|
|
| 1 | 5 | 5 | – | 10 | 19 | 16 |
| L | (670) | 905–1,354 (1,077) | 1,000–1,200 (1,100) | 1,100–1,300 (1,200) | 1,001–1,200 (1,100) | 800–1,200 (1,000) | 1,040–1,306 (1,163) |
| a | (27.0) | 19.6–24.0 (21.4) | 24.6–28.8 (26.9) | – | 17.6–23.5 (20.7) | 19.3–35 (25.0) | 23.6–30.3 (26.9) |
| b | (5.0) | 4.1–5.8 (4.6) | 4.5–5.1 (4.9) | 4.4–5.3 (4.9) | 4.4–5.3 (4.9) | 4.9–6.8 (5.7) | 4.9–5.9 (5.4) |
| c | (15.6) | 7.2–10.7 (9.0) | 8.9–14.5 (10.3) | 8.0–9.5 (8.8 | 8.0–9.5 (8.9) | 8.1–10.8 (9.6) | 7.0–7.9 (7.4) |
| c′ | (2.1) | 3.7–5.4 (4.6) | 3.3–5.8 (5.0) | 4.1–6.3 (5.2) | 4.1–6.3 (5.1) | 3.7–6.3 (4.7) | 5.7–7.2 (6.3) |
| V% | (58) | 42.0–53.3 (51.1) | 47.3–53.9 (49.9) | 48.8–52.3 (50.0) | 48.8–52.3 (50.2) | 39–50 (47) | 46.8–49.3 (48.0) |
| Esophagus l. | (135.3) | 218.5––262.0 (235.9) | 215–247 (228.0) | 224–248 (236) | 224–248 (238.0) | 141–207 (179) | 207.0–227.0 (217.2) |
| Corpus | – | 130.9––136.3 (135) | 134–143 (139.4) | 120–138 (129) | 120–138 (131) | 112–145 (125) | 123.5–140.0 (131.4) |
| Excretory pore | – | 163.2––202.0 (178.7) | 155–185 (16(6.5)) | 171–190 (180) | 171–190 (178) | 73–161 (133) | 148.0–183.0 (164.9 |
| Tail length | (42.9) | 102.0––142.0 (120.4) | 116–127 (121.3) | 121–144 (132) | 121–144 (131) | 81–121 (105) | 146.0–177.0 (156.4) |
| BDA | (20.8) | 25.0––28.8 (27.0) | 21–27 (22.6) | 23–31 (27) | 23–31 (26) | 18–28 (24.4) | 22.5–27.0 (24.8) |
| Ant. to NR | (85.1) | 140.1––151.0 (146.1) | 145–166 (154.5) | – | 144–165 (157) | 110–145 (130.5) | 131.0–155.5 (145.0) |
| Phasmid to anus | – | 27.1––43.0 (37) | 40–47 (43.2) | 33–46 (39) | 33–46 (39.8) | 32–51 (40.5) | 55.0–78.5 (64.8) |
| Ant. to deirid | – | – | 154–166 (159.0) | 166–190 (178) | 166–190 (178) | 129–152 (138) | 149.0–183.0 (165.4) |
| BD at vulva | (24.8) | 43.6––58.9 (48.2) | 42–50 (45.6) | – | – | 35–64 (49) | 33.5–45.5 (40.4) |
| BD maximum | (24.8) | 45.9––60.8 (50.1) | 38–48 (41.4) | 46–67 (57) | 46–67 (55.8) | 30–55 (41) | 38.0–48.0 (43.3) |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNematode management and characterization studies · Parasite Biology and Host Interactions · Marine Biology and Ecology Research
Accurate delimitation of nematode genera is essential for biodiversity assessments and ecological studies. Chambersiella (Cobb, 1920) and Geraldius (Sanwal, 1971) are two morphologically similar genera within the family Chambersiellidae. Their distinction is based solely on the number of ovaries in females – a character whose diagnostic value is here reconsidered. This note integrates classical descriptions, field collections, and archival notes from Cobb to reexamine the status of these genera.
Materials and Methods
We reviewed the original descriptions of Chambersiella rodens Cobb, 1920 (Cobb, 1920) and Geraldius bakeri Sanwal, 1971 (Sanwal, 1971), and a typewritten note by N. A. Cobb preserved in the USDA Nematode Collection, Beltsville, MD (Fig. 1). Unfortunately, the type specimen could not be found. Comparisons focused on ovary number, the sole feature used to differentiate the genera. Morphological data were compiled from published illustrations, morphometrics, and direct field observations.
(A) Typewritten note by N. A. Cobb and filed with the type specimen of Chambersiella rodens Cobb, 1920 pointing out that the specimen had two uteri. (B) Photomicrograph of a female of Chambersiella sp. from entry number 11034, slide G-1671, from the USDANC, collected by J. Saunders via Gerald Thorne or Mae Noffsinger from the beetle gallery of Scolytus mutistriatus Marsham, 1802 showing a didephic ovary. (C) Photomicrograph of a female of Chambersiella rodens Cobb, 1920 collected from a seed cone of Japanese cedar, Cryptomeria japonica (L. f.) D. Don.
Additionally, specimens were collected from the tree bark of pine (Pinus spp.) and oak (Quercus spp.), and Spanish moss, Tillandsia usneoides (L.) L. in Alabama, Florida, Georgia, Maine, New Hampshire, North Carolina, South Carolina, Tennessee, Vermont, and Virginia.
Results and Discussion
Cobb’s original description includes the phrase, “The gently tapering ovary contains 50–100 ova; reflexed to near anus,” suggesting a posteriorly reflexed second ovary. This interpretation is confirmed by a note from Cobb stating, “Examination of a young, but mature, stained female Chambersiella showed about 32 spermatozoa in the two uteri, somewhat more in one than the other” (Fig. 1A). If Cobb observed “spermatozoa in two uteri,” this suggests that either the female was didelphic (most likely), or the second structure was mistakenly called a uterus, but could have been a seminal receptacle, uterine diverticulum, or an abnormality.
Only two slides of Chambersiella sp. were found in the USDANC from entry number 11034, slides G-1671 and G-1672, collected by J. Saunders via Gerald Thorne or Mae Noffsinger from the beetle gallery of Scolytus mutistriatus Marsham, 1802, which shows a didelphic ovary (Fig. 1B). Unfortunately, the type specimen was not found.
All specimens that were collected along the eastern coast of the United States from both oak and pine trees were all didelphic. No monodelphic specimens were recovered, even in areas where C. rodens was originally reported. Measurements (Table 1) and photomicrographs (Fig. 1C) of specimens identified by the authors as C. rodens are presented here as further support that Chambersiella is didelphic.
Table 1:: Morphometric of females of Chambersiella rodens Cobb, 1920, C. inserrai Cid del Prado et al., 2021 C. bakeri Sanwal, 1957, C. galapagoensis Cid del Prado, 2012 and C. jejuensis Mwamula et al., 2025 from the original descriptions, additional measurements of C. bakeri by Holovachov, and original measurements of C. rodens by Eisenback and Vieira.
After more than 100 years, Chambersiella remains monotypic, whereas Geraldius contains at least five species (Cobb, 1920; Sanwal, 1957; Cid del Prado, 2012; Cid del Prado et al., 2021; Mwamula et al., 2025). These findings cast doubt on the use of ovary number as a valid generic character and suggest that the genera are not distinct. The cardial morphology, cephalic cirri, and esophageal anatomy are otherwise indistinguishable between the two groups.
Preliminary molecular studies on Geraldius inserrai Cid del PradoVera et al., 2021 and G. jejuensis Mwamula et al., 2025 (Cid del Prado et al., 2021; Mwamula et al., 2023) suggest close phylogenetic affinity with other Geraldius species, lending support to their inclusion within the same genus. If Chambersiella does indeed have two ovaries, it has not been reported since its description; therefore, molecular studies cannot be completed on nematode that cannot be found. Moreover, integrative taxonomic frameworks employing multiple loci and species delimitation methods (e.g., General Mixed Yule-Coalescent [GMYC] (Zhang et al., 2013), Automatic Barcode Gap Discovery [ABGD] Puillandre et al., 2012), or Bayesian Poisson Tree Processes [bPTP]) can further substantiate the collapse of these two genera into a unified Chambersiella sensu lato (Derycke et al., 2010).
Therefore, the integration of classical morphology, historical taxonomic intent (Cobb, 1920), and modern molecular diagnostics represents a comprehensive path forward for resolving this generic controversy in Chambersiellidae.
Conclusion
The historical separation of Chambersiella and Geraldius based solely on ovary number is undermined by Cobb’s archived notes and extensive field observations. We propose that Geraldius is a junior synonym of Chambersiella, and we formally transfer the following species: Geraldius bakeri; G. galapagoensis Cid del Prado, 2012; G. inserrai; and G. jejuensis to Chambersiella.
Proposed Taxonomic Changes
Chambersiella Cobb, 1920 syn. Geraldius Sanwal, 1971Type species: Chambersiella rodens**Chambersiella bakeri Sanwal, 1957Chambersiella galapagoensis. (Cid del Prado, 2012) n. comb.Chambersiella inserrai (Cid del Prado et al., 2021) n. comb.Chambersiella jejuensis (Mwamula et al., 2025) n. comb.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cid del Prado V. I.2012 Two new species of nematodes (Cephalobida: Chambersiellidae) from moss from North and South America Nematropica 42108114
- 2Cid del Prado V. I.Ferris H.Subbotin S. A.2021 A new species of Geraldius inserrai sp. n (Rhabditida: Chambersiellidae) from Mexico Nematropica 516777
- 3Cobb N. A.1920 One hundred new NEMAS Contributions to a Science of Nematology 9217343
- 4Derycke S.De Ley P.Tandingan De Ley I.Holovachov O.Rigaux A.Moens T.2010 Linking DNA sequences to morphology: Cryptic diversity and population genetic structure in marine nematodes Molecular Phylogenetics and Evolution 56122423310.1016/j.ympev.2010.01.027 · doi ↗
- 5Holovachov O.Esquivel A.Bongers T.2003 Free-living nematodes from nature reserves in Costa Rica. 4. Cephalobina Nematology 5111510.1163/156854102765216632 · doi ↗
- 6Mwamula A. O.Bae C. H.Lee D. G.Kim Y. S.Lee Y. D.Lee D.2025 Description and molecular characterization of Geraldius jejuensis n. sp. (Nematoda: Chambersiellidae) from Korea Journal of Nematology 5712025202310.2478/jofnem-2025-0023 PMC 1218193440547570 · doi ↗ · pubmed ↗
- 7Puillandre N.Lambert A.Brouillet S.Achaz G.2012 ABGD, automatic barcode gap discovery for primary species delimitation Molecular Ecology 2181864187710.1111/j.1365-294X.2011.05239.x 21883587 · doi ↗ · pubmed ↗
- 8Sanwal K. C.1957 Chambersiellidae n. fam. (Nematoda) with emended diagnosis of the genus Chambersiella Cobb, 1920, description of C. bakeri n. sp., and discussion of taxonomic position Canadian Journal of Zoology 3561562110.1139/z 57-054 · doi ↗