Genome-wide identification and expression analysis of Phenylalanine Ammonia-Lyase (PAL) in cucumber
Jiaojiao Zhang, Kunyang Wang, Shuai Meng, Qiushuang Han, Ming Gao

TL;DR
This study identifies and analyzes 13 Phenylalanine Ammonia-Lyase genes in cucumber, linking their expression to fruit astringency and tannin content.
Contribution
The study provides new insights into the role of PAL genes in cucumber fruit astringency through genome-wide identification and expression analysis.
Findings
Thirteen PAL genes were identified in cucumber with conserved domains and under purifying selection.
Six CsaPAL genes showed differential expression between high- and low-astringency fruits and correlated with tannin content.
Collinear relationships were found between cucumber and Arabidopsis PAL genes, suggesting evolutionary conservation.
Abstract
Phenylalanine ammonia-lyase (PAL) is the key catalytic enzyme that initiates the phenylpropanoid metabolic pathway. This study identified 13 members of the cucumber PAL family to elucidate their characteristics and their potential role in the development of cucumber fruit astringency. All identified members exhibited high conservation and contained three conserved domains: the MIO domain, the core domain, and the insertion shielding domain. Evolutionary pressure selection analysis suggested that purifying selection was the primary driving force behind the evolution of PAL family members. Cis-acting element analysis demonstrated that CsaPALs responded to light, hormones, and stress. Collinearity analysis revealed collinear relationships between AtPAL2 and CsaPAL2, as well as between AtPAL4 and CsaPAL8. Transcriptome sequencing exhibited significant differences in the expression levels of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
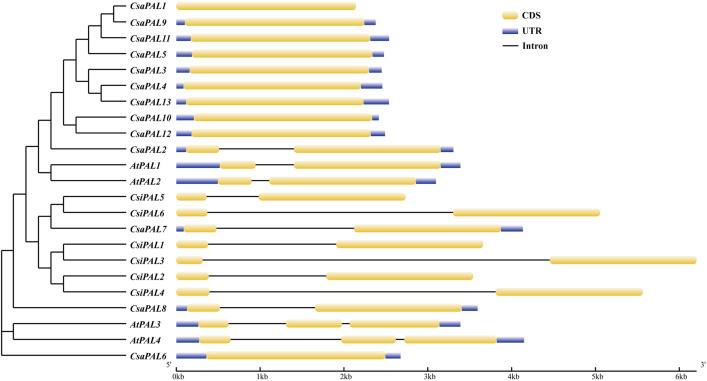 Fig 1
Fig 1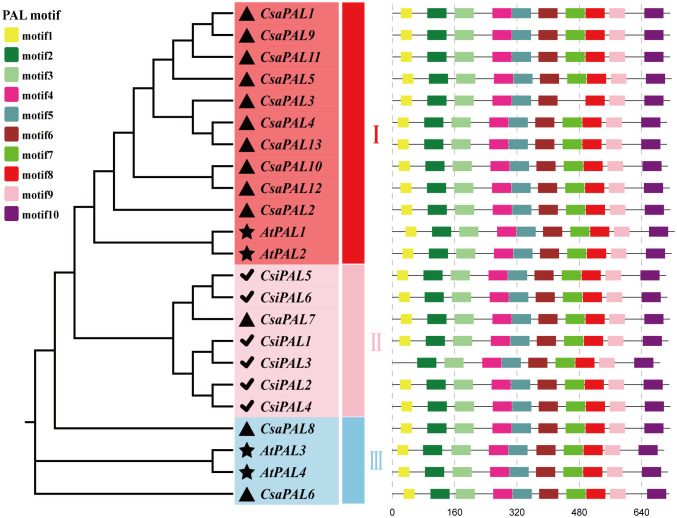 Fig 2
Fig 2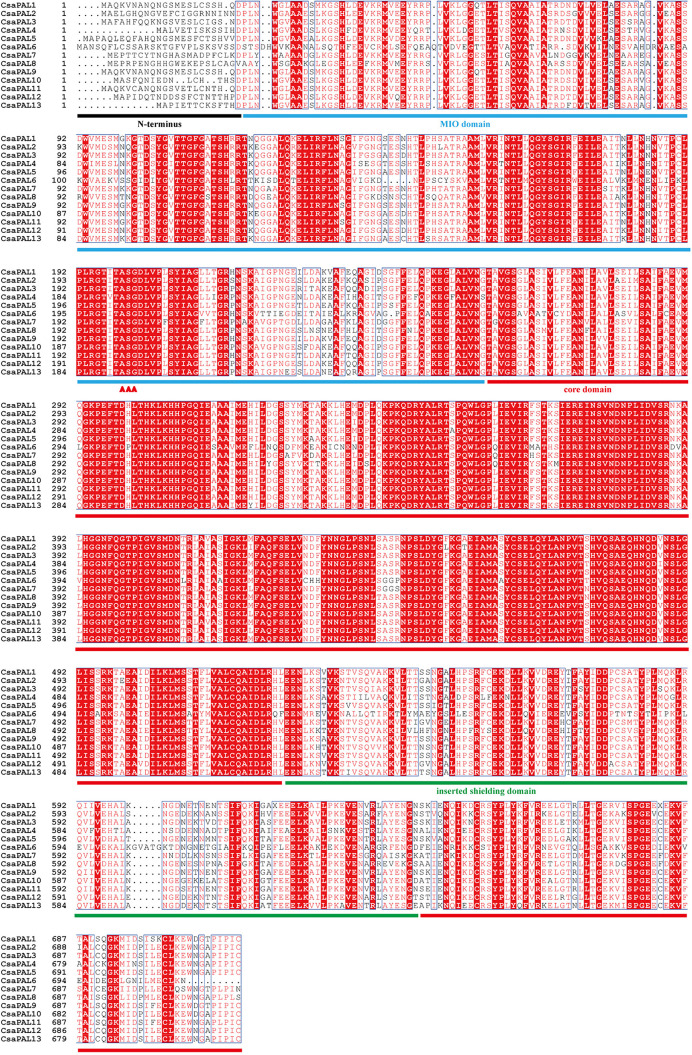 Fig 3
Fig 3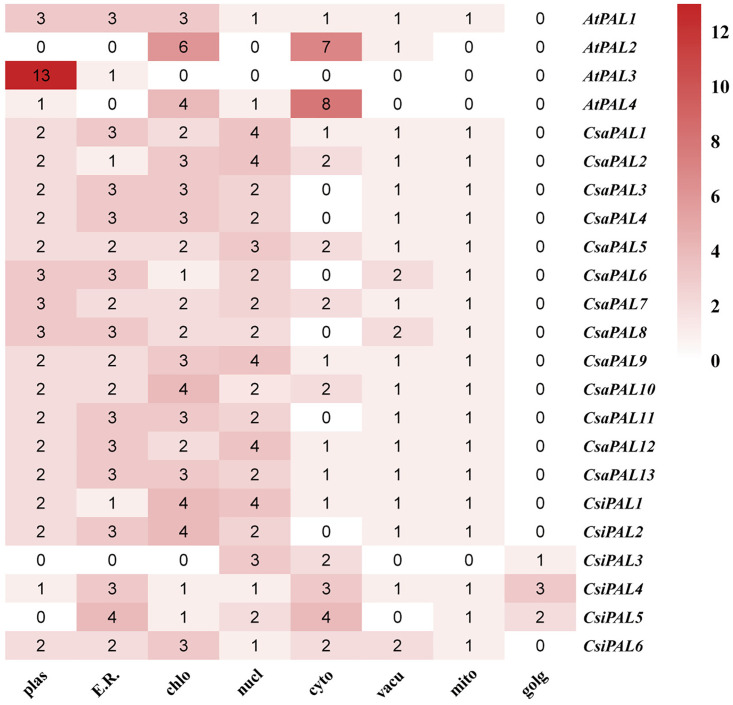 Fig 4
Fig 4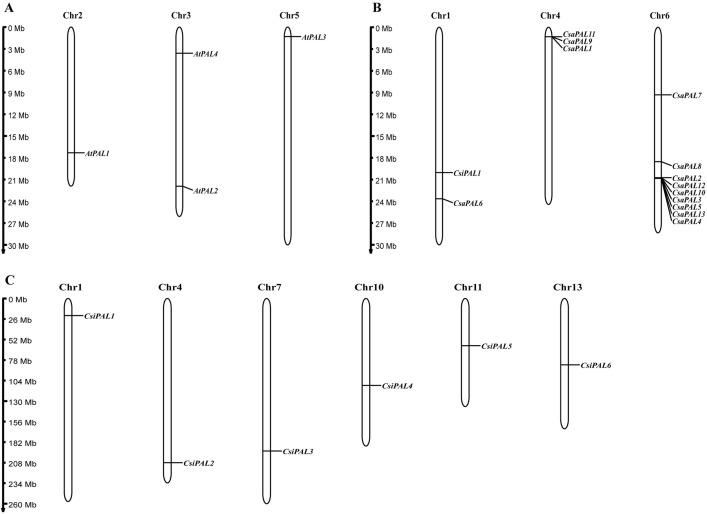 Fig 5
Fig 5 Fig 6
Fig 6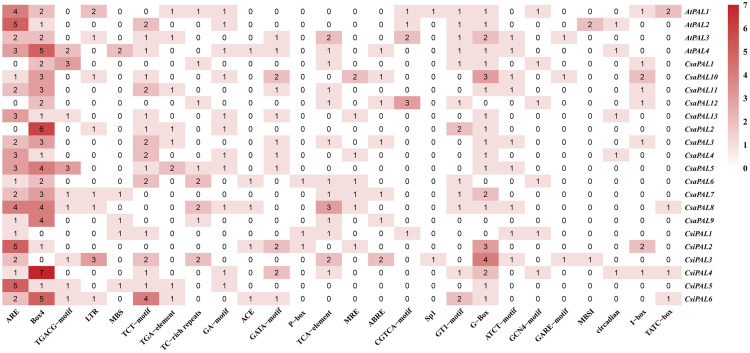 Fig 7
Fig 7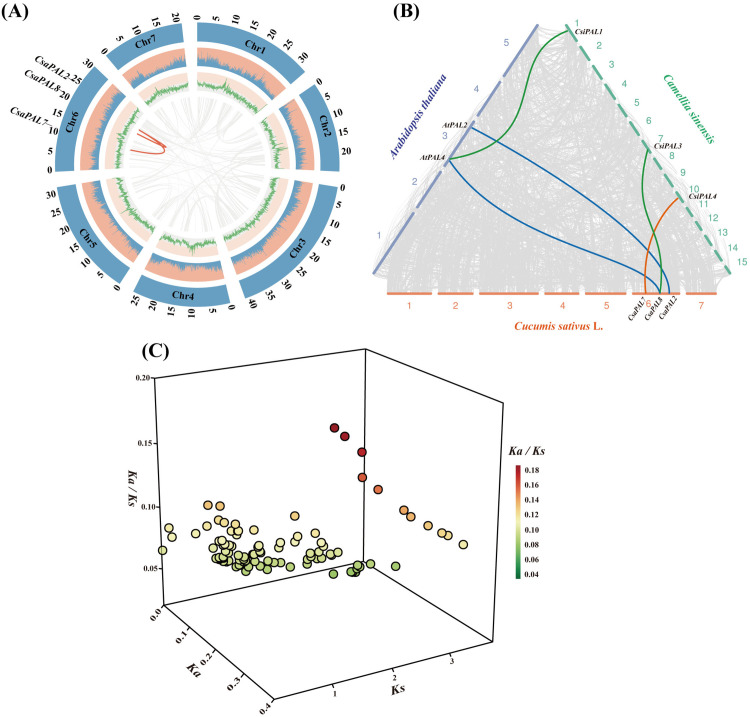 Fig 8
Fig 8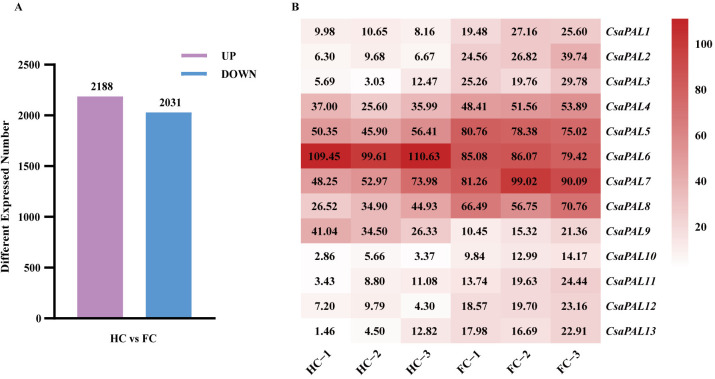 Fig 9
Fig 9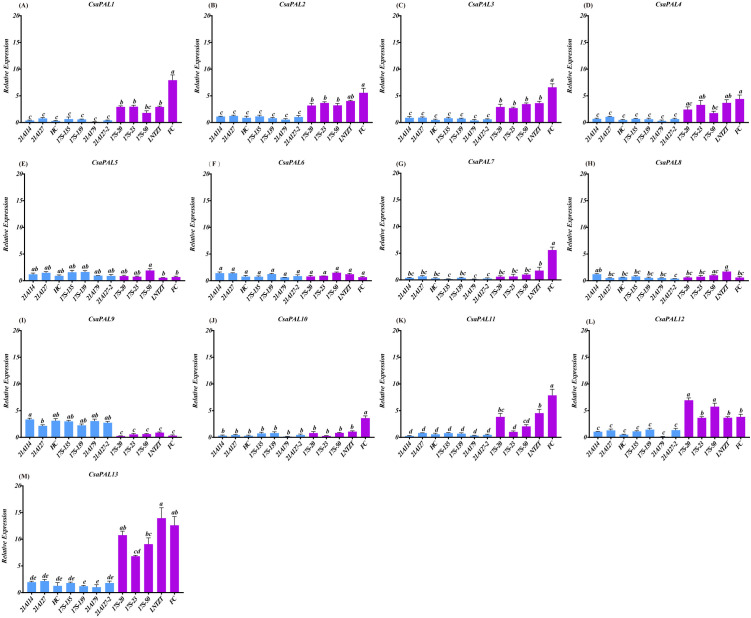 Fig 10
Fig 10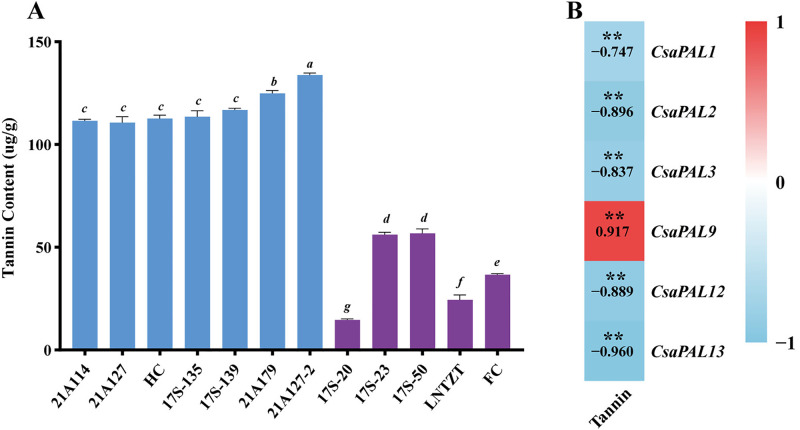 Fig 11
Fig 11- —http://dx.doi.org/10.13039/100008970Hebei Normal University of Science and Technology
- —http://dx.doi.org/10.13039/100016073Key Technologies Research and Development Program of Anhui Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Gene Expression Analysis · Postharvest Quality and Shelf Life Management · Plant biochemistry and biosynthesis
Introduction
In recent years, rising living standards and improved quality of life have driven greater demand for high-quality horticultural products with taste emerging as a key priority [1]. Among various quality-related concerns, astringency in fruits and vegetables has garnered particular attention. Astringency is perceived through sensations such as dryness, roughness, contraction, and lingering adhesion in the oral cavity [2]. The main compounds responsible for this characteristic include polyphenols, organic acids, inorganic acids, and certain proteins [3–7]. In plant-based foods, astringency primarily results from polyphenolic compounds, including flavonoids, phenolic acids, and tannins [8–10]. Several studies have established tannins as the primary contributors to astringency in most horticultural crops [4,10,11]. For instance, grapes contain substantially higher levels of tannins in their pericarp, seeds, and pedicels compared to other tissues, which makes them particularly astringent [12,13]. In tea, the principal compounds responsible for astringency are tea polyphenols and flavonoid glycosides [12,14]. Similarly, the progressive accumulation of proanthocyanidins in persimmons leads to a significant increase in the astringency of persimmon fruits [15,16]. Likewise, in walnuts, the gradual increase in the content of hydrolysable tannins in the walnut seed coat during maturation significantly enhances their astringency [17].
Phenylalanine Ammonia-Lyase (PAL) functions as the initial enzyme in the phenylpropanoid metabolic pathway [18–20]. It catalyzes the conversion of phenylalanine into trans-cinnamic acid [21], which is then hydroxylated by Cinnamate-4-hydroxylase (C4H) to form p-Coumaric acid. This p-Coumaric acid is subsequently activated by 4-Coumaroyl-CoA ligase (4CL) to produce p-Coumaroyl-CoA [22]. In the flavonoid biosynthesis pathway, chalcone synthase (CHS) catalyzes the reaction between p-Coumaroyl-CoA and malonyl-CoA forming chalcone, the key precursor that initiates flavonoid production. Chalcone is further isomerized by chalcone isomerase (CHI) and hydroxylated by flavanone 3-hydroxylase (F3H) to form dihydroflavonol. Dihydroflavonol serves as the precursor for anthocyanins, tannins, and other flavonoids. Through the action of dihydroflavonol 4-reductase (DFR), dihydroflavonol is reduced to leucoanthocyanidin, which is subsequently transformed into proanthocyanidins in plants [23–25]. This metabolic pathway highlights the crucial role of PAL genes in driving the biosynthesis of anthocyanins, tannins, and various flavonoids in plants [26]. PAL was initially discovered in barley [27], and later research has identified PAL genes across a wide range of plant species, including Arabidopsis [28], banana [29], rice [30], wheat [31], walnut [32], and grape [33]. PAL enzymes are highly conserved and typically feature an MIO domain, which houses the enzyme’s active site. In most plant PAL enzymes, the MIO domain contains a conserved active site sequence ‘GTITASGDLVPLSYIAG’, with the characteristic Ala-Ser-Gly (ASG) motif as a key component localized within this sequence [34–36].
Cucumber (Cucumis sativus L.) is an important horticultural crop cultivated worldwide for its fresh and edible fleshy tissues [37]. As a result, its flavor plays a critical role in determining its commercial value. Despite this, the molecular mechanisms underlying astringency development in cucumber remain inadequately understood, and the role of PAL family genes in this process has not yet been reported. To date, research on cucumber PAL genes has focused mainly on stress responses [38]. In tea, astringency is mainly caused by tea polyphenols, and elevated expression of PAL in green tea increases polyphenols concentrations, leading to greater astringency [39]. To elucidate the role of PAL genes in cucumber astringency development, this study conducted a comprehensive genome-wide analysis of the PAL gene family in cucumber. This included gene structure characterization, protein secondary structure prediction, subcellular localization prediction, chromosomal mapping, cis-acting element identification, collinearity assessment, and evaluation of evolutionary selection pressures. In addition, transcriptional expression profiling of PAL genes was performed in cucumber fruits with varying levels of astringency, and the correlation between tannin content in cucumber fruits and PAL genes expression was examined. This study aims to advance the understanding of the molecular functions of PAL family genes in cucumber and to reveal their potential roles in the development of astringency in cucumber fruits.
Materials and methods
Plant materials
The cucumber specimens employed in this study were sourced from the greenhouse of the College of Horticultural Science & Technology at Hebei Normal University of Science & Technology, located at 119°10’ E, 39°42’ N. The experimental samples included strongly astringent cucumber varieties (‘21A114’, ‘21A127’, ‘HC’, ‘17S-135’, ‘17S-139’, ‘21A179’, ‘21A127-2’) and mildly astringent ones (‘17S-20’, ‘17S-23’, ‘17S-50’, ‘FC’, ‘LNTZT’), all of which belong to the South China-type cucumber. The astringency index (AI) of each cucumber material is shown in S1 Table [40]. Grading criteria for the cucumber astringency index (AI): AI = 0 indicates no astringency, AI < 40 indicates slight astringency, 40 ≤ AI < 70 indicates moderate astringency, and AI ≥ 70 indicates strong astringency. For each sample, fruit materials (sampled as a mixture of peel and pulp) were harvested 9 days post pollination and three biological replicates. These samples were promptly frozen in liquid nitrogen and temporarily stored at −80°C to facilitate subsequent experimental analyses.
Genome-wide identification of the PAL gene family members
The PAL family members of Arabidopsis, tea, and cucumber were obtained from the online database TAIR (https://www.arabidopsis.org/), TPIA (https://tpia.teaplants.cn/) and CuGenDB (http://cucurbitgenomics.org), respectively. The Arabidopsis genome is from TAIR10.1, and its corresponding NCBI RefSeq assembly is GCF_000001735.4. The cucumber genome is from Cucumber_9930_V3, with the corresponding NCBI RefSeq assembly being GCF_000004075.3. The tea genome data is from the relevant information of Tieguanyin in the TPIA database. The HMMER software (version 3.4) was used for verification to obtain a gene set with high credibility. The final PAL gene set was obtained based on the conserved domains of PAL proteins.
Gene structure and chromosomal localization
Based on the gene sequences of the PAL family, the structure of each PAL genes was obtained on the GSDS2.0 online software (https://gsds.gao-lab.org/index.php) [41]. Meanwhile, with the help of the results of local BLAST analysis, all PAL genes in each species were mapped to their respective chromosomal positions. Subsequently, each gene was systematically renamed to reflect its chromosomal location, so as to improve the clarity and organization of the genomic data for further analysis.
Protein conserved domain and secondary structure analysis
We employed the online software MEME to analyze the conserved motifs within the PALs (https://meme-suite.org/meme/) [42]. The conserved motifs were visualized to highlight the conserved domains of the PAL family proteins using the online tool Evolview (https://www.evolgenius.info/evolview/#/) [43]. The online resource Expasy (https://web.expasy.org/) [44] was used to analysis of the secondary structure of these proteins.
Phylogenetic analysis
The amino acid sequences of PALs were imported into the MEGA software (version 11.0.13) for sequence alignment using the ClustalW algorithm [45]. Subsequently, the Neighbor-Joining method was employed to construct a phylogenetic tree with a Bootstrap parameter set to 1000 replicates. The resulting newick (nwk) data were then analyzed further using the online software Evolview (https://www.evolgenius.info/evolview/#/), providing a visual representation of the evolutionary relationships among the PAL family members.
Cis-acting element analysis
We developed a Python script (version 3.8.1) to extract 2000 base pairs upstream from each gene to obtain the promoter sequence. The cis-acting elements within these promoter regions were identified using the Plant Care online tool (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/) [46] and visualized with the Evolview software (https://www.evolgenius.info/evolview/#/). The pheatmap package (version 1.0.12) in the R programming language (version 4.3.3) was employed to generate a heatmap analyzing the distribution and frequency of these cis-acting elements.
Subcellular localization prediction
The amino acid sequences of all PALs were analyzed using WoLF PSORT (https://wolfpsort.hgc.jp/) [47] to predict their subcellular localization. The pheatmap package (version 1.0.12) in the R programming language (version 4.3.3) was utilized to create a heatmap.
Collinearity analysis and Ka/Ks analysis
The collinearity analysis was carried out using software or tools such as jcvi software (version 1.4.16) [48], seqtk software (version 1.4), Bedtools (version 2.31.1) [49], Perl (version 5.22.0), and Circos (version 0.69.9) [50] respectively. For the Ka/Ks analysis, we first used the Simple Ka/Ks Calculator of TBtools (version 2.142) [51] to calculate the Ka, Ks, and Ka/Ks values for each gene pair. Based on these values, we employed the R programming language (version 4.3.3, with the plot3D package version 1.4.1 and RColorBrewer package version 1.1–3) to generate a three-dimensional scatter plot for further analysis.
RNA isolation, RNA-seq, reverse transcription and qRT-PCR
In a previous report, we examined the astringency levels of fruits (9 days post-pollination, dpp) from 228 cucumber germplasm resources, ‘FC’ (low-astringency) and ‘HC’ (high-astringency) were screened [40]. To identify the major genes influencing cucumber astringency, we selected cucumber materials ‘HC’ and ‘FC’ for transcriptome sequencing. First, we use the RNAprep Pure Plant Kit (TIANGEN, China) to extract total RNA from cucumber fruits (9 days post-pollination), and assessed the extraction quality on a 1% agarose gel. Sample purity was determined using a NanoDrop ND2000 spectrophotometer. Select the total RNA from high-astringency cucumber material ‘HC’ and low-astringency cucumber material ‘FC’ for transcriptome sequencing. The total RNA was used to synthesize cDNA with the FastQuant RT reagent kit (Tiangen, China) for subsequent experiments. Then, to further verify the association between PAL genes and the formation of cucumber astringency, we selected 12 cucumber materials with extreme astringency traits (7 high-astringency cucumbers and 5 low-astringency cucumbers), and obtained the expression levels of CsaPALs in these materials through qRT-PCR. Real-time quantitative PCR was performed on a BIO-RED instrument using the SYBR^®^ Plus Universal qPCR Kit. The qRT-PCR program comprised an initial denaturation at 95°C for 2 minutes, followed by 40 cycles of 95°C for 5 seconds, 60°C for 10 seconds, and 72°C for 15 seconds (S2 Table). The CsUBI gene was selected as the internal reference gene, and relative expression levels were calculated using the 2^-ΔΔCt^ method. Statistical analysis (p < 0.05, one-way ANOVA, Tukey test) and graph generation were conducted using GraphPad Prism software.
Determination of tannin content and correlation analysis
The tannins in cucumber fruits (sampled as a mixture of peel and pulp) were extracted with dimethylformamide (DMF) solution, followed by determination using a colorimetric method. Correlation analysis was carried out using SPSS software (Version 27.0.1.0).
Results
Genome-wide identification and gene structure analysis of PAL
In this study, a total of 23 PAL genes were identified (Table 1) across three species: Arabidopsis thaliana (4 genes), Camellia sinensis (6 genes), and Cucumis sativus L. (13 genes). The amino acid sequences encoded by these PAL genes ranged in length from 687 amino acids (CsiPAL3) to 725 amino acids (AtPAL1), with molecular masses varying between 74889.74 Da (CsiPAL3) and 78725.73 Da (AtPAL1). The physicochemical properties of these proteins were then analyzed, revealing that all PALs are acidic in nature (pI < 7). In addition, their GRAVY index values were all below zero, indicating that the proteins are highly hydrophilic. The analysis of chromosomal loci showed that PAL genes are predominantly clustered on cucumber chromosome 6, implying that potential tandem duplication events may have contributed to their expansion on this chromosome [52].
Table 1: Gene information of the PALs in various species.
To gain a deeper understanding of the PALs, gene structure diagrams were initially constructed. The results revealed that the gene lengths in the PAL family genes range from 2 kb to 6 kb. Specifically, in cucumber, the fragment sizes of PAL genes are predominantly between from 2 kb and 3 kb. In contrast, the tea gene CsiPAL3 exceeds 6 kb, with introns comprising more than 50% of its length. The number of exons in PAL genes typically range from 1 to 3. In cucumber, most PAL genes contain a single exon, except for CsaPAL2, CsaPAL7, and CsaPAL8, which have two exons each (Fig 1).
The gene structures of the PALs.The yellow rectangles represent exons, the black solid lines represent introns, and the blue rectangles represent UTR.
Analysis of conserved motifs and phylogenetic relationships of the PAL family proteins
To explore the phylogenetic relationships among genes in the PAL families, the amino acid sequences of PAL proteins from Arabidopsis, tea, and cucumber were compared, and subsequently phylogenetic trees were constructed for each species (Fig 2). The analysis revealed that the 23 PALs clustered into three distinct groups, containing 12, 7, and 4 members, respectively. The evolutionary analysis indicated that 10 out of the 12 genes in group Ⅰ were derived from cucumber PAL genes (CsaPAL1-CsaPAL5 and CsaPAL9-CsaPAL13), implying a close evolutionary relationship and high homology among them. In contrast, the 6 PAL genes from tea were clustered in group Ⅱ, showing a moderate degree of homology with the CsaPAL7 gene in cucumber. Within the PAL gene families, 10 conserved protein motifs were identified (Fig 2, S3 Table). Motif 1 and 9 were 29 and 41 amino acids long, respectively, while all other motifs were 50 amino acids in length. Notably, CsaPAL3 lacked motif 7, and CsiPAL3 lacked motif 1. The presence of these conserved motifs across the PAL family proteins highlights the high degree of homology among PAL genes.
The composition of conserved motifs and phylogenetic tree of the PAL family proteins.The amino acid sequences of the PAL family are from Arabidopsis (4), tea (6), cucumber (13). The PALs of Arabidopsis were marked with black star, the tea is marked with a black check, and the cucumber is marked with a black triangle.
Previous studies have established that eukaryotic PAL proteins contain three conserved domains: the MIO domain, the core domain, and the inserted shielding domain. The MIO domain encompasses the MIO motif, which includes the catalytic triplet Ala-Ser-Gly [53–55]. To investigate cucumber PALs, we performed multiple sequence alignment of their amino acid sequences. All CsaPAL proteins were found to contain the N-terminal domain, the MIO domain, the core domain, and the inserted shielding domain, with the ASG tripeptide motif located in the MIO domain. These domain characteristics closely matched those previously reported for PAL proteins in plants (Fig 3).
Multiple sequence alignment of cucumber PALs.The black solid line represents the N-terminus, the blue solid line represents the MIO domain, the red solid line represents the core domain, and the green solid line represents the inserted shielding domain. The conserved Ala-Ser-Gly (ASG) tripeptide motif specific to the PALs are marked with red triangles.
Analysis of the protein secondary structure and prediction of subcellular localization
The secondary structure of proteins plays a crucial role in determining their function. To better understand the potential functional characteristics of PAL genes, we analyzed the secondary structures of PAL family members (S4 Table), including α-helix, extended strand, β-turn, and random coil. The relative proportions of these structures varied among the PAL family members, with α-helix accounting for 50.19%−88.51%, extended strands for 4.91%−7.67%, β-turn for 1.15%−5.93%, and random coils for 10.34%−39.15%.
Predictions of subcellular localization indicate that these PALs are primarily found in eight cellular compartments (Fig 4); the plasma membrane, endoplasmic reticulum, chloroplast, nucleus, cytoplasm, vacuole, mitochondrion, and Golgi apparatus. In cucumber, CsaPAL proteins are predominantly localized in the plasma membrane, endoplasmic reticulum, chloroplast, and nucleus indicating that they may play important functional roles in these specific cellular locations.
Subcellular localization prediction of the PAL family members.The larger the value, the higher the possibility that the protein is located in the organelle. ‘plas’ represents the plasma membrance, ‘E.R.’ represents the endoplasmic reticulum, ‘chlo’ represents the chloroplast, ‘nucl’ represents the nucleus, ‘cyto’ represents the cytosol, ‘vacu’ represents the vacuole, ‘mito’ represents the mitochondrion, ‘golg’ represents the golgi apparatus.
Chromosomal localization of the PAL genes
To visually examine the 23 PAL genes, their chromosomal locationswere mapped out. In Arabidopsis, the 4 PAL genes are distributed across chromosomes 2, 3, and 5 (Fig 5). In cucumber, the 13 PAL genes are located on chromosomes 1, 4, and 6, while in tea, the 6 PAL genes are positioned on chromosomes 1, 4, 7, 10, 11, and 13, respectively. Notably, in cucumber, CsaPAL1, CsaPAL9, and CsaPAL11 form a cluster on chromosome 4, while CsaPAL2, CsaPAL3, CsaPAL4, CsaPAL5, CsaPAL10, CsaPAL12, and CsaPAL13 cluster on chromosome 6. These two PAL gene clusters suggest high sequence similarity and potential functional conservation within the cucumber PAL gene family.
Visualized localization of the PAL genes on the chromosomes of each species.Chromosomal localization analysis of Arabidopsis (A), cucumber (B), and tea (C).
Analysis of Cis-acting elements
The analysis of cis-acting elements within the promoter sequences (from −2000 bp to −1 bp) of PAL genes revealed 26 distinct cis-acting elements. These elements can be grouped into three main categories: light-responsive elements (Box4, G-Box, TCT-motif, GT1-motif, GATA-motif, MRE, I-box, ATCT-motif, GA-motif, Sp1, ACE), hormone-responsive elements (ABRE, TGACG-motif, CGTCA-motif, TCA-element, P-box, TGA-element, GARE-motif, TATC-box), and stress-responsive elements (ARE, LTR, TC-rich repeats, MBS, circadian). Of these, 24 elements were detected in the promoter sequences of cucumber PAL genes (Figs 6 and 7).
The distribution of cis-acting elements in the promoter regions of PALs.Different colors represent different cis-acting elements.
The distribution of the number of cis-acting elements in the promoter regions of the PAL genes.
The cis-acting element analysis demonstrated that the Box4 element is present in the promoter regions of all cucumber PAL genes. Among them, CsaPAL2, CsaPAL3, CsaPAL5, CsaPAL6, CsaPAL7, CsaPAL8, CsaPAL9, CsaPAL10, and CsaPAL11 showed relatively strong responses to the Box4 element. Additionally, CsaPAL1 exhibited the most significant response to the TGACG-motif element, while CsaPAL4 and CsaPAL13 primarily responded to the ARE element (Fig 7). Overall, these findings suggest that members of the cucumber PAL gene family are involved in regulating responses to light, hormones, and various stresses.
Analysis of gene duplication, collinearity and evolutionary selection pressure
To explore the mechanisms of gene amplification within the PAL gene family, we conducted a collinearity analysis across the cucumber genome (Fig 8A). This analysis revealed that CsaPAL2 has undergone tandem duplication events in conjunction with both CsaPAL7 and CsaPAL8. This duplication could potentially influence the expression levels of the PAL gene. Furthermore, we performed a cross-species horizontal collinearity analysis of PAL family genes using cucumber, Arabidopsis, and tea. Collinearity blocks were constructed to visualize these relationships (Fig 8B). The results showed that CsaPAL2 is collinear with AtPAL2, CsaPAL8 is collinear with AtPAL4 and CsiPAL3, and CsaPAL7 is collinear with CsiPAL4. Additionally, AtPAL4 exhibits collinearity with CsiPAL1. These relationships suggest that collinear genes may share similar or complementary functions across species.
Analysis of gene duplication, collinearity and gene selection pressure of the PAL family.(A) Collinearity analysis within the cucumber genome. From the outside to the inside, they represent the distribution of each chromosome of cucumber, the distribution of the number of genes on each chromosome of cucumber, and the distribution of GC content in the cucumber genome. The gray and red arcs are the connections of blocks with collinearity. (B) Inter-species collinearity analysis of Arabidopsis, cucumber and tea. The gray, orange, green and blue arcs are the connections of blocks with collinearity. (C) Analysis of evolutionary selection pressure on homologous gene pairs of the PAL family. The x-axis, y-axis and z-axis represent the Ka value, Ks value and Ka/Ks value of the gene pairs respectively.
To investigate the selection pressures acting on the PAL family genes during evolution, we calculated key evolutionary indicators for each gene pair: Ka (frequency of non-synonymous mutations), Ks (rate of synonymous mutations), and the Ka/Ks ratio. The Ka/Ks ratio serves as an indicator of selection pressures exerted on genes over time. The analysis revealed that all PAL gene pairs have Ka/Ks ratios below 1, implying that purifying selection has predominantly shaped their evolutionary trajectory.
Expression profile analysis of CsaPALs genes and their correlation with tannin content
According to the RNA-Seq results, the comparison between ‘HC’ and ‘FC’ revealed 4219 differentially expressed genes (DEGs), including 2188 upregulated and 2031 downregulated genes (Fig 9A). Furthermore, the expression levels of 13 PAL family members showed significant variation between the two lines (Fig 9B). Specifically, CsaPAL6 and CsaPAL9 were expressed at significantly higher levels in ‘HC’ compared to ‘FC’, whereas the remaining CsaPALs genes displayed higher expression in ‘FC’ (Fig 9B).
Analysis of differentially expressed genes in cucumber with different astringency levels.(A) The number of up-regulated and down-regulated DEGs of ‘HC’ vs ‘FC’. (B) Gene expression heatmap drawn based on the FPKM data from transcriptome sequencing results.
The qRT-PCR results showed that CsaPAL1, CsaPAL2, CsaPAL3, CsaPAL12, and CsaPAL13 were expressed at significantly higher levels in low-astringency cucumbers compared to high-astringency ones. In contrast, CsaPAL9 exhibited significantly higher expression in high-astringency cucumbers (Fig 10). These expression patterns aligned with the transcriptome sequencing results (Figs 9 and 10). Furthermore, high-astringency cucumbers generally contained more tannins than low-astringency ones, and the tannin content was significantly correlated with the expression levels of the six PAL genes (Fig 11). These findings suggest that CsaPAL1, CsaPAL2, CsaPAL3, CsaPAL9, CsaPAL12, and CsaPAL13 are potential candidate genes involved in regulating cucumber astringency.
The expression profiles of the CsaPALs in cucumber with different astringency.Blue represents materials with high astringency, purple represents materials with low astringency. The cucumber UBIQUITIN gene was used as the reference transcript. Error bars represent the standard error from three independent experiments. Symbols a, b, c, d, and e indicate a significant difference at the p-value of 0.05.
Correlation between tannin content and expression levels of PAL candidate genes in cucumber fruit materials with different astringency levels.(A) Tannin content in cucumber fruit materials with different astringency levels. Symbols a, b, c, d, e, f, and g indicate significant differences at the p-value of 0.05. (B) Correlation heatmap of tannin content and expression levels of PAL candidate genes. The values in the boxes represent Pearson correlation coefficients. ** indicates a significant correlation at the level of p < 0.01.
Discussion
In the phenylpropanoid metabolic pathway, the enzyme PAL catalyzes the conversion of phenylalanine into cinnamic acid. As the initial enzyme in this metabolic pathway, PAL serves as a critical rate-limiting enzyme for the production of downstream metabolites and plays an essential role in tannin biosynthesis in plants [37]. Previous studies have extensively documented the genome-wide identification of PAL gene family members in various plant species [32,56–59]. In this study, we identified 13 PAL genes in cucumber (Fig 1). These genes exhibit structural variation, with exons counts ranging from 1 to 3 and variations in intron length and number (Fig 1, Table 1). This structural diversity may contribute to the functional differences, suggesting that different CsaPAL genes could participate in the synthesis of distinct secondary metabolites. For instance, in Arabidopsis, the AtPAL1 and AtPAL2 genes are mainly associated with flavonoid biosynthesis, while AtPAL3 and AtPAL4 are primarily involved in lignin biosynthesis [60]. Therefore, further research is required to clarify whether CsaPAL genes play a role in the biosynthesis of flavonoids and lignin in cucumber.
The plant PAL family genes are typically classified into three main sub-families [32,61,62]. In this study, we constructed a phylogenetic tree to illustrate the evolutionary relationships among PAL proteins from Arabidopsis, tea, and cucumber, and identified three distinct groups (Fig 2). The results showed that the 13 PAL proteins in cucumber are mainly distributed across two sub-families (Group I and Group III), a pattern consistent with findings in walnut [63], sorghum [64], and melon [65]. Furthermore, the CsaPAL genes in Group I share similar exon-intron structures and protein motifs, and cluster with AtPAL1 and AtPAL2, which are known to function in flavonoid biosynthesis in Arabidopsis (Fig 2). This suggests that the CsaPAL genes in Group I may also play a role in flavonoid biosynthesis in cucumber.
The identification and analysis of cis-acting elements in gene promoter regions provides insight into gene transcriptional regulation and expression patterns. Research has demonstrated that the promoter regions of PAL family genes contain numerous cis-acting elements involved in hormone and stress responses [31,36]. In this study, the cis-acting elements in the promoter regions of CsaPALs genes were categorized into three main groups (Fig 6): light-responsive elements, hormone-responsive elements, and stress-responsive elements. Each CsaPAL gene promoter region contains at least one hormone response-related cis-acting element (Fig 7). Interestingly, while the ERE element is found in all PAL genes promoters in Hevea brasiliensis [64], it was absent in the promoter regions of CsaPAL genes, indicating that cis-acting elements in the promoters of the PAL family vary between species. Certain cis-acting elements appear exclusively in the promoter regions of specific CsaPAL genes (Fig 7). For instance, the P-box was detected in the promoter region of the CsaPAL6 gene, the CGTCA-motif was found in the promoter region of CsaPAL12, and the GARE-motif was identified in the promoter region of CsaPAL10 (Fig 7). Several studies have shown that light can influence the astringency of fruits. For example, during the growth of grapes, increasing light exposure and temperature through pruning can significantly enhance the astringency of grapes [53]. Conversely, reducing ultraviolet light exposure can lower the astringency of tea leaves [54]. Cherries grown outdoors exhibit stronger astringency compared to those grown in greenhouses [55]. High temperatures can intensify the astringency of fruits. Swede, a root vegetable, experiences an increase in fruit astringency at high temperatures, while the astringency is greatly reduced at low temperatures [66]. In addition, gibberellin and auxin also affect the astringency of tea leaves [67]. Therefore, the light-responsive, hormone-responsive, and stress-responsive cis-acting elements in the promoter regions of cucumber PAL genes may affect the expression level of the PAL gene, and thus influence the astringency of cucumbers. The exact functions of these cis-acting elements in the promoter regions of these genes still need to be further studied and clarified.
Analyzing genetic selection pressure plays an important role in clarifying the evolutionary processes of species. In this context, Ka represents the rate of non-synonymous mutations, referring to the frequency of mutations in a gene sequence that cause amino acid changes, while Ks represents the rate of synonymous mutation, reflecting the frequency of mutations that do not affect the encoded amino acid sequence [68]. The Ka/Ks ratio represents the relative frequency of these two mutation rates and is commonly used to assess the selection pressure a gene experiences during evolution. A Ka/Ks value less than 1 indicates that the main driving force for gene evolution comes from purifying selection, which removes deleterious mutations [68]. In this study, we analyzed the Ka/Ks ratios of PAL genes in Arabidopsis, tea, and cucumber. The results showed that the Ka/Ks ratios of all CsaPALs genes were less than 1 (Fig 8C), indicating that these CsaPAL genes are evolutionarily conserved, with stable structures and consistent functions [69]. Additionally, we conducted a genomic collinearity analysis of Arabidopsis, tea, and cucumber, which revealed collinear relationship between AtPAL2 and CsaPAL2, as well as between AtPAL4 and CsaPAL8 (Fig 8B). Previous studies have demonstrated that the AtPAL2 and AtPAL4 genes of Arabidopsis are involved in the synthesis of flavonoids and lignin, respectively [60]. However, additional research is needed to determine whether CsaPAL2 and CsaPAL8 perform similar functions in the metabolism of flavonoid and lignin compounds in cucumber.
Catechins and flavonoids are the main substances that cause the astringency of tea [14]. During the maturation of tea leaves, the abundance of catechins is significantly positively correlated with the expression of PALs [70,71]. In albino tea plants (AnJiBaiCha), the expression of PAL is negatively correlated with the content of catechins [72]. Here, the expression levels of the 13 cucumber PAL genes in materials with different astringency levels showed that 6 CsaPAL genes might be related to the astringency of cucumber. Based on the expression patterns of CsaPAL in cucumber materials, the differential expression of CsaPAL genes between high- and low-astringency cucumber materials shows a “bidirectional regulation” pattern: the expression level of CsaPAL9 in high-astringency materials (‘HC’) is significantly higher than that in low-astringency materials (‘FC’), while CsaPAL1, CsaPAL2, CsaPAL3, CsaPAL12, and CsaPAL13 show the opposite trend (higher expression in low-astringency materials) (Fig 10). Additionally, the expression levels of these six genes are significantly correlated with the fruit tannin content (Fig 11B). These results suggest that there may be a “functional division” in the regulation of astringency within the cucumber PAL gene family: CsaPAL9 may contribute to the high-astringency phenotype by positively regulating tannin synthesis, while the other five genes may indirectly affect the astringency level by negatively regulating tannin accumulation or diverting the metabolic flux towards other phenylpropanoid derivatives (such as lignin). Similar functional differentiations have been reported in other plants. For example, in Arabidopsis, AtPAL1 and AtPAL2 are mainly involved in the synthesis of flavonoids, while AtPAL3 and AtPAL4 are more involved in lignin synthesis [60]. This functional division may be an adaptive strategy developed by the PAL gene family through gene duplication and functional diversification during long-term evolution to meet the diverse demands for phenylpropanoid metabolites in different tissues and at different developmental stages. It is worth noting that the seven CsaPAL genes (CsaPAL4, CsaPAL5, CsaPAL6, CsaPAL7, CsaPAL8, CsaPAL10, CsaPAL11) with no significant expression differences detected in this study are not necessarily non-functional. They may play roles in other developmental stages or specific tissues of cucumber. For example, CsaPAL8 shows collinearity with AtPAL4 in Arabidopsis, which is mainly involved in lignin synthesis during stress responses (Fig 8B). It is speculated that when cucumbers face adverse conditions (such as low temperature, diseases), CsaPAL8 may be highly expressed to promote lignin accumulation and improve plant resistance, rather than regulating fruit astringency. The specific functions of these genes in the formation of cucumber astringency still need to be further verified.
Compared with previous studies on the phenylalanine ammonia-lyase (PAL) gene family [38], we combined the cucumber-specific online database CuGenDB with the hidden Markov model of the PAL domain (PF00221.24) to identify 13 cucumber PAL genes (CsaPAL1-CsaPAL13), covering the cucumber PAL gene family more comprehensively. Additionally, the study by Amjad et al. focused on the regulatory role of PAL in cucumber thermotolerance, while our study emphasizes the regulation of cucumber astringency quality by PAL family genes and has identified candidate genes. The results of this study not only provide new perspectives for a deeper understanding of the molecular mechanism of cucumber astringency formation but also offer target genes for further improving cucumber astringency traits through methods such as gene editing and marker-assisted breeding.
Conclusions
In this study, we identified 13 members of the PAL gene family in cucumber and investigated their physicochemical properties, protein structures, conserved motifs and domains, cis-acting elements, phylogenetic relationships, evolutionary selection pressures, collinearity, and expression patterns, and correlation analysis between the tannin content and the expression levels of PAL genes. Our findings demonstrated that cucumber PAL genes are highly conserved and contain specific conserved domains characteristic of eukaryotic plant PAL genes. Transcriptome sequencing analysis revealed substantial differences in the FPKM expression levels of 13 PAL genes between the high-astringency cucumber ‘HC’ and the low-astringency cucumber ‘FC’. Furthermore, qRT-PCR verification confirmed that six genes, CsaPAL1, CsaPAL2, CsaPAL3, CsaPAL9, CsaPAL12, and CsaPAL13, were significantly differentially expressed between ‘HC’ and ‘FC’. Additionally, the expression levels of these six genes were significantly correlated with cucumber fruit tannin content. These results suggest that these six genes are likely the candidates underlying cucumber astringency. This study laid a foundation for further exploration of the biological functions of cucumber PAL genes and provided valuable references for cucumber breeding aiming at astringency traits.
Supporting information
S1 TableEvaluation information of astringency index for the experimental materials of cucumber.(XLSX)
S2 TableThe primers used in qRT-PCR.(XLSX)
S3 TableThe detailed information of the conserved motifs of the PAL family proteins.(XLSX)
S4 TableDetailed information on the secondary structure of the PAL family proteins.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gao L, Hao N, Wu T, Cao J. Advances in understanding and harnessing the molecular regulatory mechanisms of vegetable quality. Front Plant Sci. 2022;13:836515. doi: 10.3389/fpls.2022.836515 35371173 PMC 8964363 · doi ↗ · pubmed ↗
- 2Tezotto-Uliana JV, de Paula JT, Tessmer MA, Kluge RA. Ethanol vapor is efficient for reduction of astringency compounds in cashew apple. Postharvest Biol Technol. 2018;145:117–24. doi: 10.1016/j.postharvbio.2018.07.002 · doi ↗
- 3Wen L, Sun L, Chen R, Li Q, Lai X, Cao J, et al. Metabolome and microbiome analysis to study the flavor of summer black tea improved by stuck fermentation. Foods. 2023;12(18):3414. doi: 10.3390/foods 12183414 37761123 PMC 10527649 · doi ↗ · pubmed ↗
- 4Luo Y, Zhang X, Luo Z, Zhang Q, Liu J. Identification and characterization of micro RN As from Chinese pollination constant non-astringent persimmon using high-throughput sequencing. BMC Plant Biol. 2015;15:11. doi: 10.1186/s 12870-014-0400-6 25604351 PMC 4308916 · doi ↗ · pubmed ↗
- 5Lee CA, Ismail B, Vickers ZM. The role of salivary proteins in the mechanism of astringency. J Food Sci. 2012;77(4):C 381-7. doi: 10.1111/j.1750-3841.2012.02644.x 22515235 · doi ↗ · pubmed ↗
- 6Yang HH-L, Lawless HT. Descriptive analysis of divalent salts. J Sens Stud. 2005;20(2):97–113. doi: 10.1111/j.1745-459X.2005.00005.x 16614749 PMC 1435863 · doi ↗ · pubmed ↗
- 7Sano H, Egashira T, Kinekawa Y, Kitabatake N. Astringency of bovine milk whey protein. J Dairy Sci. 2005;88(7):2312–7. doi: 10.3168/jds.S 0022-0302(05)72909-X 15956294 · doi ↗ · pubmed ↗
- 8Soares S, Brandão E, Guerreiro C, Soares S, Mateus N, de Freitas V. Tannins in food: insights into the molecular perception of astringency and bitter taste. Molecules. 2020;25(11):2590. doi: 10.3390/molecules 25112590 32498458 PMC 7321337 · doi ↗ · pubmed ↗