Predation on scyphozoan polyps and selective hydrozoan nematocyst incorporation dynamics in an alien aeolid nudibranch
Hila Dror, Tamar Lotan, Dror Angel

TL;DR
A nudibranch species, Caloria militaris, preys on jellyfish polyps but does not use their stinging cells for defense, which is a new finding in nudibranch feeding behavior.
Contribution
This is the first report of complete selectivity in nematocyst sequestration in a nudibranch with a functional cnidosac.
Findings
Caloria militaris consumes various scyphozoan polyps and can survive on them for up to 258 days.
Despite intense predation, C. militaris does not incorporate scyphozoan nematocysts into its cnidosac.
Predation by C. militaris may help regulate jellyfish blooms in the eastern Mediterranean.
Abstract
Aeolid nudibranchs prey on various cnidarians and incorporate nematocysts from their prey into the cnidosac, located at the tip of their cerata for self-defense. This study examined the predator–prey interactions between two nudibranch species, Caloria militaris and Flabellina affinis, and various cnidarians, focusing on scyphozoan polyps from common eastern Mediterranean medusae, including Aurelia sp. Cassiopea andromeda, Phyllorhiza punctata, and Rhopilema nomadica. An initial short-term experiment assessed predation by both nudibranch species, after which subsequent experiments focused exclusively on C. militaris. Short-term experiments for 24 h and long-term experiments lasting up to 258 days were conducted to determine whether C. militaris consumes a variety of cnidarians, and if it incorporates the prey nematocysts into its cnidosacs. Laboratory experiments indicate that while F.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2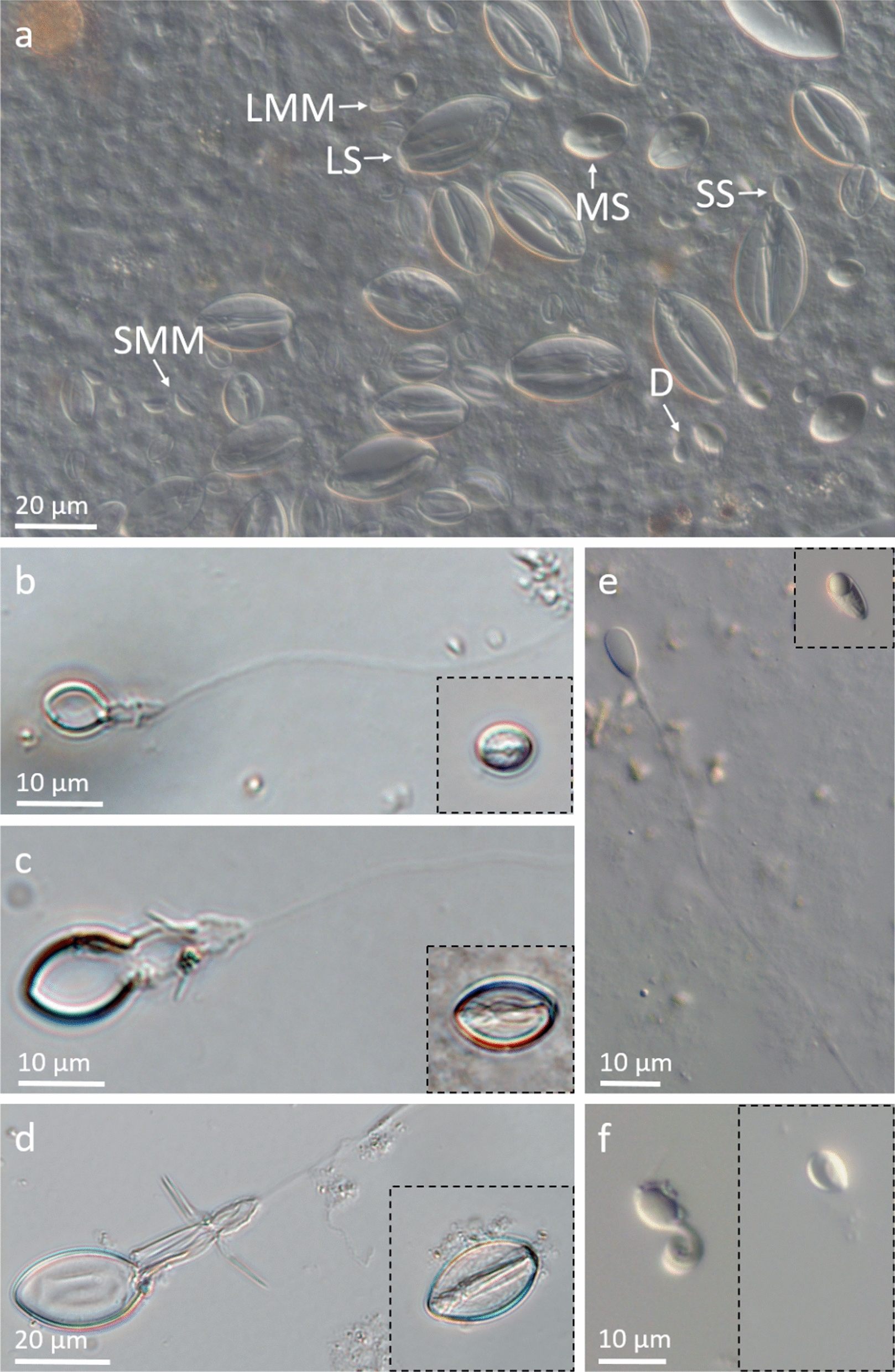 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine Invertebrate Physiology and Ecology · Marine Toxins and Detection Methods · Marine Ecology and Invasive Species
Background
The eastern Mediterranean has been experiencing massive scyphomedusa blooms during the last several decades, which have negative ecological, economic, and societal effects [1–5]. Scyphozoans typically exhibit a complex life cycle that includes a pelagic medusa stage and a benthic polyp stage. Polyps are considered a primary driver of jellyfish outbreaks since they reproduce asexually and can persist for extended periods, releasing numerous young medusae each season [6, 7]. However, due to their small size and cryptic nature, the location of natural populations of polyps of most Scyphozoa remains unknown. Here we concentrated on Scyphozoa species common to the eastern Mediterranean. These include Cassiopea andromeda (Forskål, 1775) (Family: Cassiopeidae), Phyllorhiza punctata (von Lendenfeld, 1884) (Family: Mastigiidae), and Rhopilema nomadica (Galil, Spanier and Ferguson, 1990) (Family: Rhizostomatidae), which are Indo-Pacific species alien to the eastern Mediterranean [8], whereas Aurelia (Lamarck, 1816) (Family: Ulmaridae) is a cosmopolitan genus [9]. Since the 1980’s, R. nomadica has become the dominant jellyfish species in the eastern Mediterranean, with populations present throughout most of the year [10]. It is considered one of the "100 worst invasive species" in the Alien Invasive Species Inventories for Europe [11] due to its negative ecological and economic effects and rapid expansion westward [1, 12].
Aeolid nudibranchs (Gastropoda: Nudibranchia) are shell-less marine gastropods that prey on cnidarians such as jellyfish, anemones, and hydroids [7, 13–16]. Many nudibranchs are highly specialized feeders, targeting specific prey, and their larvae settle where prey is abundant [7, 14, 17, 18]. Nudibranchs have evolved defenses against the stinging cells of cnidarians, which contain specialized, syringe-like organelles called nematocysts [19, 20]. These defenses include protective gut linings, specialized epithelial structures, and mucus secretion [13, 21–25]. Through kleptocnidy nudibranchs retain and use their prey’s nematocysts for defense, storing them in cnidosacs at the tips of their cerata (dorsal appendages) [21, 23, 26]. Not all nematocysts are retained, indicating selective sequestration [27–29], which may vary as a result of prey availability [30] and the presence of nudibranch predators [31], with some species preferring to incorporate scyphozoan nematocysts when multiple prey types are available [32].
Typically, selectivity in nematocyst incorporation is manifested as shifts in the relative abundance of nematocyst types between the prey and the cnidosac [25, 33, 34]. For instance, C. verrucosa preying on Aurelia sp. polyps, incorporated a higher ratio of rhopaloids to a-isorhizas, compared to that present in the polyps [35] and preferentially retained scyphozoan nematocysts over those from other prey species [30].
Kleptocnidy varies widely among nudibranchs, not only in terms of nematocyst selectivity, but also in transfer time, turnover rate, and storage capacity [36, 37]. Transfer may occur within hours (e.g., Cratena peregrina) [38] to days (e.g., Berghia stephanieae (Valdés, 2005)) [39], and complete turnover within days (e.g., Spurilla neapolitana) (Delle Chiaje, 1841) [25] to weeks (e.g., H. crassicornis) [40]. These variations highlight species-specific strategies.
The retained nematocysts may persist within the nudibranch's body for extended periods, ranging from days to weeks. Since the inventory of nematocysts (termed cnidome) facilitates taxonomic identification of Cnidaria [41], the presence and composition of nematocysts within the cerata can provide insights into the nudibranch's dietary history, serving as a record of the past and more recently consumed prey [21, 28, 42, 43], as well as the location of these prey.
Jellyfish blooms have significant ecological, economic, and social impacts worldwide [44] with scyphopolyps playing a key role in driving these bloom formations [6, 9, 45]. However, the location of the polyps of most scyphozoan species in situ is unknown. Since this is a major challenge with respect to understanding polyp ecology, many methods (mainly molecular approaches) have been employed to address this problem [42, 46–48]. Mills and Miller [49] were able to identify the prey of the ctenophore Haeckelia rubra (Kölliker, 1853) through nematocyst analysis in its predator. Thus, analysis of the nematocysts in the cerata of predator nudibranchs may aid in locating scyphopolyp populations.
Here, we focus on two aeolid species, Flabellina affinis (Gmelin, 1791) (Family: Flabellinidae) (Fig. 1a), a native aeolid to the Mediterranean, and Caloria militaris (Alder and Hancock, 1864) (Family: Facelinidae) (Fig. 1b), a Lessepsian migrant [50]. F. affinis, measuring up to 40 mm in length, is present throughout the Mediterranean and along the Atlantic African coasts. It is known to inhabit various habitats including seaweed beds, rocky reefs, and sandy seafloors, up to 10 m deep, and was found to feed on hydroids of the genus Eudendrium (Ehrenberg, 1834) [51]. C. militaris was originally described in the Bay of Bengal, India, and has been recorded since throughout the Indo-West Pacific [50, 52]. This species is known to inhabit natural hard substrates, although it has also been found on artificial substrates up to 30 m deep [50]. C. militaris, measuring up to 40 mm in length, is known to feed on hydroids such as Calyptospadix cerulea (Clarke, 1882) (Family: Bougainvilliidae), Ectopleura larynx (Ellis and Solander, 1786) (Family: Tubulariidae), and Pennaria disticha (Goldfuss, 1820) (Family: Pennariidae) [14, 52]. Along the Israeli Mediterranean coast, C. militaris is now commonly found in large numbers during winter and spring.Fig. 1. The aeolid nudibranchs a F. affinis and b* C. militaris*. Arrows mark the tips of the cerata. Photos: Amit Zeevi
We examined the interactions between aeolid nudibranchs and cnidarian, focusing on polyps of scyphozoan species commonly found in the eastern Mediterranean. We hypothesized that certain nudibranch species exhibit a more generalist feeding behavior than previously reported, capable of surviving solely on scyphozoan polyps when their known food (Hydrozoa) is scarce. Initial short-term experiments were conducted to compare predation between the two nudibranch species, after which subsequent experiments focused exclusively on C. militaris to assess its predation on various cnidarian polyps and to evaluate whether these can sustain the nudibranch as a primary food source over extended periods. In the next step, the incorporation of nematocysts from prey scyphozoan polyps into the cnidosac of C. militaris was tested. Several nudibranch species are capable of effectively reducing scyphozoan polyp populations through predation [7, 32, 53–55]. Since polyps represent a key life stage driving jellyfish blooms, their removal may lead to a subsequent decline in bloom formations [56]. Therefore, understanding the feeding behavior of nudibranchs and their predatory impact on scyphozoan polyps is important in our quest to understand the dynamics of scyphozoan populations.
Methods
Scyphozoan polyp cultures
Cultures of Rhopilema nomadica polyps were produced by laboratory fertilizations of four to eight sexually mature male and female jellyfish collected near Mikhmoret, Israel, eastern Mediterranean Sea (32°24′23″N 34°52′24″E). To enable planula settlement and polyp production, glass microscope slides were placed in a fertilization tank with the adult medusae following our described protocol [57]. Cassiopea andromeda medusae and polyps have been maintained in the laboratory since the discovery of their polyps on a rock collected near Jisr az-Zarqa, Israel, eastern Mediterranean (32°53′32″N, 34°90′10″E). Polyps of Phyllorhiza punctata were produced after collecting planulae from the oral arms of a gravid female medusa collected near Mikhmoret and allowing them to settle onto glass slides. Polyps of an as yet unidentified Aurelia sp. were cultivated to mature medusae and maintained in the laboratory from three polyps that appeared in our flowing seawater aquaria. All scyphozoan medusae were identified based on descriptions from Galil et al. (and references therein) [8]. Each species' polyps (size 0.5–3 mm) were kept in separate containers in a 50 μm filtered flowing seawater aquarium system, maintained at ambient sea surface temperature (SST) ranging annually from 17 to 29 °C. The polyps were fed daily with newly hatched Artemia salina (Linnaeus, 1758) nauplii and subjected to a 12-h illumination cycle.
Nudibranch sampling and maintenance
Between 2020 and 2024, 34 Caloria militaris and 4 Flabellina affinis nudibranchs were collected by SCUBA diving near Mikhmoret. Identification of the nudibranch species was based on [50] and [58]. The nudibranchs were found on natural rocky (kurkar rock) reefs at depths of 6–12 m during winter and spring (SST 17–23 °C). Each nudibranch was carefully hand-collected into a 50 ml tube and brought to the laboratory for handling. The nudibranchs were kept in 1 L containers of 50 μm filtered flowing seawater, aerated and maintained at SST, subjected to a 12-h illumination cycle. All 34 nudibranchs were used in the feeding experiments, and 13 of these were also used in the nematocyst incorporation experiments.
Short-term predation of nudibranchs on various cnidarian
Short-term feeding experiments were conducted in 200 ml glass bowls of 50 μm filtered seawater with a glass slide (7.5 × 2.6 cm^2^) holding the prey items. The prey items were chosen from various cnidarian classes based on availability in our laboratory.
The nudibranchs were starved for 72 h, then four C. militaris (size 1.5–2.5 cm) and four F. affinis (size 1–1.5 cm) individuals were each provided with 13–28 R. nomadica polyps (Scyphozoa, size 1–2 mm). In addition, four C. militaris individuals (size 1.5–2.5 cm) were each provided with a different cnidarian prey – seven Exaiptasia diaphana (Rapp, 1829) (Anthozoa anemone, size 1–2 cm), 14 polyps of Oculina patagonica (de Angelis D'Ossat, 1908) (Anthozoa coral, polyp size 1.5–2 mm), eight polyps of C. andromeda (Scyphozoa, 2–3 mm), six branches of Pennaria disticha (Hydrozoa, polyp size < 1–2 mm) (Supp. Fig. S1). Another set of three C. militaris (size 2–3 cm) were first starved for 24 h and then individually introduced into bowls containing three–five 10 cm long tentacles removed from R. nomadica medusa (40 cm bell diameter).
The nudibranchs were observed by a Motic SMZ-171 dissecting microscope for three hours to study their feeding behavior, including time to the first encounter with a polyp, response to the encounter, and the manner of feeding. Following this initial close-up examination, the nudibranchs were each placed in an aerated flowing seawater container, maintained at SST, and were allowed to feed for an additional 21 h (overall 24 h) to record predation rate (number of polyps consumed day^−1^), and the predation proportion (number polyps consumed / number polyps provided).
Long-term predation of C. militaris on scyphozoan polyps
The long-term experiments were conducted using only C. militaris nudibranchs since F. affinis did not prey on scyphozoan polyps in the short-term experiment. Each of the 34 C. militaris nudibranchs (sizes 15–30 mm, measured on collection day) were starved for 24 h and then placed in a 1 L container with flowing 50 µm filtered seawater along with polyps of different scyphozoan species on glass slides. In order to decrease size bias, various sizes of nudibranchs were examined for each prey species. The provided prey polyps included Aurelia sp., C. andromeda, P. punctata, and R. nomadica (according to availability of polyps at the time of nudibranch collection). To study the predatory capacity of the nudibranchs, polyps were provided at four dose levels per day, from low to very high (≤ 20, ≤ 50, ≤ 80, and > 80 polyps d^−1^, respectively). The polyps were replenished twice weekly, and the predation rate d^−1^, predation proportion d^−1^, and survival period of C. militaris were documented. During the experiment all nudibranchs appeared in good condition: feeding continuously, growing, moving, and some laid eggs.
Review of literature
To better understand the potential effect of predation of C. militaris on scyphozoan polyps, we scanned the literature for nudibranch species known to feed on scyphozoan polyps and their reported predation rates. The search was performed during October 2024 using Google Scholar with the search words: nudibranch, predator, scyphozoa, polyp, scyphistoma.
Prey nematocysts in nudibranch cerata
Microscopy and imaging
Examination of cerata and nematocyst samples was performed using Differential Interference Contrast (DIC) microscopy on a Zeiss AXIO Imager.M2 microscope (20X or 40X magnification) and images were captured with a digital Axiocam 503 color camera. Measurements of nematocysts were conducted using a Zeiss Zen-Pro 2.5 program.
Identification of incorporated nematocysts in the cnidosac of C. militaris
The nematocyst incorporation experiment was conducted on 13 freshly collected C. militaris nudibranchs. Prior to the experiment, 3 to 5 cerata from each of the nudibranchs were removed and stored at −20 °C for identification of nematocysts in nudibranchs collected at sea. Next, the nudibranchs were starved for 96 h and immersed in 5% KCl for 30 s to clear previously-obtained nematocysts from their cnidosacs [59]. After this treatment, cnidosacs in cerata of three nudibranchs were inspected under a microscope to confirm that all nematocysts were ejected.
The nudibranchs were used in two consecutive experiment sets with various prey species (12 days each). Between the first and the second sets, the nudibranchs were treated with 5% KCl to eject previously-incorporated nematocysts from the cerata. In the first set, nudibranchs were provided with polyps of the scyphozoan Aurelia sp. (n = 5 nudibranchs) and C. andromeda (n = 5 nudibranchs), and the hydrozoan Nemalecium lighti (n = 2 nudibranchs). In the second set, nudibranchs were provided with polyps of the scyphozoan R. nomadica (n = 6 nudibranchs) and the hydrozoan P. disticha (n = 2 nudibranchs) (see supp. Table S1 for details). P. disticha was collected from a pier in Jaffa port, Israel (32°03′09″N 34°44′57″E) and N. lighti was sampled by SCUBA diving from a 6–12 m deep reef near Mikhmoret. For hydrozoan DNA identification see below. P. disticha served as a positive feeding control as C. militaris is known to feed on this hydrozoan [14, 52]. Two–three cerata were removed from each nudibranch on the last day of the experiment (day 12) and stored at −20 °C. In addition, for nudibranchs feeding on Aurelia sp., C. andromeda, and P. disticha, cerata were removed from two nudibranchs on day 3, 6, and 9 of the experiment to determine the timeline for nematocyst incorporation into the cnidosac (see supp. Table S1 for sample details). The number of polyps consumed by each nudibranch were counted each time food items were replaced to check that the nudibranchs were actively feeding (supp. Table S2). Squash preparations of the cerata were examined and photographed to check for the incorporation of nematocysts of the various food items in the nudibranch’s cnidosacs. Nematocysts were identified based on the nomenclature proposed by Weill [60] and modified by Östman [41], and the relative abundances of the various nematocyst types in the cnidosacs were documented. For each nematocyst type, width and length of undischarged capsules (n = 10) were measured.
Nematocyst isolation and characterization from the hydrozoan P. disticha
Colonies of the hydrozoan P. disticha were sampled from a pier in Jaffa port. Nematocysts were isolated from homogenate of 3–5 branches (2–3 cm long) of P. disticha (n = 7) using 50% Percoll (Sigma-Aldrich, USA) followed by centrifugation at 1000 g for 10 min at 4 °C [61]. The resulting pellet was re-suspended in double distilled water (DDW) and stored at −20 °C. Nematocysts discharge was induced by addition of 1 µl of 0.1 M NaOH to 3 µl nematocyst suspensions placed on glass slides. For each sample, the relative abundances of the various types of nematocysts were documented. The identification and description of each nematocyst type based on Östman [41] included measurements of capsule width and length of discharged and undischarged capsules (n = 10). The nematocyst complement of the other prey species used are not presented here because they were not incorporated into the cnidosacs.
DNA extractions and sequencing of prey Hydrozoa
For the identification of prey hydrozoa collected from the wild, DNA extractions were performed using the DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) and the ZymoBIOMICS DNA Miniprep Kit (Zymo Research, USA) following the manufacturer’s protocol. A 600 bp region of the mitochondrial 16S gene was amplified using the primer set SHA/SHB [62]. PCR amplifications were carried out in 25 µl reactions containing 0.5 µl of each primer, (10 µM) 12.5 µl 2X Dream Taq Green Mix (Thermo Scientific), 1 μl BSA (20 mg/ml), 0.5 µl of template DNA (10 ng), and 10 μl of nuclease-free water. Cycling conditions were: 95 °C for 2 min, 35 cycles of 95 °C for 30 s, 50 °C for 30 s, and 72 °C for 1 min with a final extension at 72 °C for 5 min. PCR products were sequenced by "HyLabs" Israel. The taxonomic identities of the sequences obtained were confirmed using BlastN in the NCBI GenBank. Hydrozoan prey species were identified as P. disticha and N. lighti.
Data analysis
Data analysis and graphs were performed using RStudio (version 2023.06.0, R Development Core Team 2023). Analysis of variance (ANOVA) was conducted to check for significant differences in predation rate and proportion of polyps consumed between the various scyphozoan polyps offered as food. All tests were conducted at the α = 0.05 significance level. Assumptions for ANOVA were checked using Levene’s test for homogeneity of variance and a Shapiro–Wilk test for normality. Tukey’s HSD tests were conducted for multiple pairwise post-hoc comparisons of means for the significantly different ANOVA results. Pearson’s correlation tests were conducted to assess the relationship between nudibranch predation rate and survival duration; and between number of polyps provided and predation rate. Permutational multivariate ANOVA (PERMANOVA) (9999 permutations) were used to test for significant differences between the cnidome of P. disticha and the nematocysts found in the cnidosacs of nudibranchs feeding on P. disticha. Permutational multivariate analyses of dispersion (PERMDISP) were conducted to test significant PERMANOVA results, to ensure that results were due to structural differences and not to unequal dispersion among groups. Similarity percentage analyses (SIMPER) were calculated to determine the main contributors to the observed dissimilarity in the cnidomes. Results are presented as % or as mean ± SD.
Results
Short-term predation of nudibranchs on various cnidarians
All nudibranchs that were placed in the bowls explored the bottom, sides, water surface, and the slide with polyps. Flabellina affinis avoided contact with Rhopilema nomadica polyps, and no polyps were consumed during the 24 h experiment (Supp. Fig. S2). In contrast, Caloria militaris retracted when its oral tentacles first contacted the R. nomadica polyp, as if stung, and either moved away or remained still. To avoid being stung when feeding, the nudibranch elevated its head, and engulfed the polyp from the top, consuming the goblet and leaving the stalk (Supp. Video S1).
During the close-up observation, three out of four C. militaris individuals were observed feeding on R. nomadica polyps. The first R. nomadica polyp was consumed 63 min into the experiment. The nudibranch consumed the goblet within a few seconds, then rested for five minutes until it continued crawling. Twenty-four hours later, between 58% (10/17) and 100% (28/28) of the polyps offered had been consumed by C. militaris nudibranchs (19 ± 8.83 polyps d^−1^). However, when C. militaris individuals were offered tentacles of adult R. nomadica medusae, they kept their distance. Upon contact with tentacles, the nudibranchs would curl up, roll and quickly crawl away, probably due to being stung.
Predation of C. militaris on Cassiopea andromeda polyps was less efficient than on R. nomadica polyps, which are usually smaller. Upon first contact, the polyp attacked the nudibranch causing it to retreat. The first C. andromeda polyp was approached only 82 min after C. militaris was added to the experimental bowl, and it took nearly 30 min for the nudibranch to fully swallow it. By the end of the 24 h experiment, the nudibranch had consumed only three out of eight C. andromeda polyps (37.5%).
In contrast to the behavior of C. militaris that was offered scyphozoan polyps, when presented with the hydrozoan, Pennaria disticha, the nudibranch began feeding immediately and continued throughout the observation period. The nudibranch would crawl on the hydrozoan branch, reach a polyp, remove it from the stalk, and swallow it whole. The sting of the hydrozoan polyps did not seem to affect the nudibranch as it didn’t retract when coming into contact with the polyp tentacles.
Unlike the hydrozoan and scyphozoan polyps, none of the anemone, Exaiptasia diaphana and coral, Oculina patagonica polyps were consumed by C. militaris. The nudibranch made some attempts to approach the anemone but was severely stung and kept away thereafter.
Long-term predation of C. militaris on scyphozoan polyps
C. militaris nudibranchs were offered four species of scyphozoan polyps as prey at four dose levels and a summary of the predation rate, proportion of polyps consumed, and survival duration are provided in Table 1 (see Supp. Table S3 and S4 for details). Table 1. Rate and proportion of tested scyphozoan polyp species (Aurelia sp*., C. andromeda, P. punctata,* and R. nomadica) consumed by C. militaris nudibranchs per day during the long-term feeding experimentsPolyp speciesNumber of polyps provided d^−1^NPredation rate (polyps d^−1^)Predation proportionMaximum survival (d)Aurelia sp.Total98.02 ± 4.690.35 ± 0.3038Low (≤ 20)78.21 ± 4.980.55 ± 0.30Medium (≤ 50)69.26 ± 5.280.29 ± 0.19High (≤ 80)31.83 ± 3.180.02 ± 0.03Very high (> 80)313.50 ± 2.290.11 ± 0.04Cassiopea andromedaTotal1714.63 ± 12.710.68 ± 0.31258Low (≤ 20)166.20 ± 3.970.69 ± 0.26Medium (≤ 50)1722.86 ± 11.120.69 ± 0.32High (≤ 80)733.40 ± 28.000.53 ± 0.42Very high (> 80)754.80 ± 37.020.54 ± 0.37Phyllorhiza punctataTotal442.84 ± 10.020.87 ± 0.0539Low (≤ 20)215.00 ± 2.361.00Medium (≤ 50)435.14 ± 9.560.85 ± 0.09High (≤ 80)361.87 ± 1.800.88 ± 0.11Very high (> 80)195.200.96Rhopilema nomadicaTotal164.67 ± 7.780.73 ± 0.1894Low (≤ 20)163.17 ± 2.030.73 ± 0.18Medium (≤ 50)131.310.89High (≤ 80)167.710.98Very high (> 80)183.170.92Presented as mean ± SD. N = number of nudibranchs in each treatmentProvided polyps: low ≤ 20, medium ≤ 50, high ≤ 80, very high > 80Each nudibranch was provided with various dose levels during the experiment, thus the total number of nudibranchs (N) at the top row of each prey species is lower than the sum of nudibranchs in the dose treatments
The nudibranchs started feeding almost immediately upon encounter with prey. When examining the predation on the different prey species, the mean (and maximum) number of polyps consumed by the nudibranchs per day was 8.02 ± 4.69, 14.63 ± 12.71, 42.84 ± 10.02, and 4.67 ± 7.78 (23, 130, 105, and 91) for Aurelia sp., C. andromeda, Phyllorhiza punctata, and R. nomadica, respectively (Supp. Table S4). The proportion of Aurelia sp. polyps consumed was lowest (ANOVA, F_(4,42)_ = 5.8, p < 0.005) and the predation rate on P. punctata polyps was highest among the prey species tested (ANOVA, F_(4,42)_ = 17.3, p < 0.005) (Supp. Fig. S3). Nudibranch survival was longest when feeding on C. andromeda polyps (78.8 ± 54.8, max 258 days) and shortest when feeding on Aurelia sp. polyps (27.5 ± 7.0, max 38 days) (Supp. Table S4). Generally, survival was longer when the predation rate was higher (Pearson’s correlation, R_(32)_ = 0.52, p < 0.005). However, this result was affected mostly by the lower predation rates (< 20 d^−1^) (R_(24)_ = 0.51, p < 0.01) and insignificant at higher predation rates (R_(6)_ = − 0.25, p > 0.5). Whereas 3,382 R. nomadica polyps were consumed during the experiment, the podocysts of R. nomadica were left untouched.
When provided with ample food (> 80 polyps d^−1^), the nudibranch consumed between 13 and 95 polyps d^−1^, depending on the prey species. Predation rates of P. militaris compared to other nudibranchs species known to feed on scyphozoan polyps are provided in Table 2. The predation rate increased when nudibranchs were provided with increasing numbers of prey (Pearson’s correlation, R_(81)_ = 0.82, p < 0.005) but did not reach a plateau. Generally, the nudibranchs did not consume all of the provided polyps even when the provided amount of food was low. Table 2. Nudibranch species that prey on scyphozoan polyps and predation rate (when available)NudibranchSchyphopolypReferencePredation rate (polyps d^−1^) Caloria militaris (Alder and Hancock, 1864)Aurelia sp., Cassiopea andromeda, Phyllorhiza punctata, Rhopilema nomadicaThis study50.20 ± 37.58Aurelia* sp.This study13.50 ± 2.29Cassiopea andromedaThis study54.80 ± 37.02Phyllorhiza punctataThis study95.20Rhopilema nomadicaThis study83.17Ceratodoris plana (Baba, 1960) (as Okenia plana) Nemopilema nomurai, Aurelia coerulea, Rhopilema esculentumTang et al. 2021 [63]30–118 **Coryphella verrucosa (Sars, 1829)Aurelia auritaHernroth and Gröndahl 1985 a, b [7, 56]200Cyanea capillataGröndahl and Hernroth 1987 [64]AureliaÖstman 1997 [35](as Flabellina verrucosa)AureliaFrick 2005 [30]Cratena pilata (Gould, 1870) (as Coryphella sp.)Chrysaora quinquecirrhaCargo and Schultz 1967 [65]11 polyps in 10 minChrysaora quinquecirrhaVogel 1969 [66]Chrysaora quinquecirrhaOakes and Haven 1971 [54]Cuthona sp. (Alder and Hancock, 1855)Chrysaora quinquecirrhaSoranno 2016 [67]Dendronotus dalli (Bergh, 1879)Aurelia labiataHoover et al. 2012 [32]Dendronotus rufus (O’Donoghue, 1921)Aurelia labiataKozloff 1983 [68]Aurelia labiataHoover et al. 2012 [32]**Eubranchus spp.**Chrysaora quinquecirrhaSorrano 2016 [67]Facelina bostoniensis (Couthouy, 1838) (as Facelina drummondi) Aurelia auritaThiel 1962 [69]Flabellina fusca (Bergh, 1894)Aurelia labiataHoover et al. 2012 [32]Goniobranchus tinctorius (Ruppell and Leuckart, 1830) (as Chromodoris tinctoria) Nemopilema nomurai, Aurelia coerulea, Rhopilema esculentumTang et al. 2021 [63]Hermissenda crassicornis (Eschscholtz, 1831)Aurelia auritaKeen 1991 [53] < 120Aurelia labiataHoover et al. 2012 [32]8.6–31.3 polyps h^−1^ **Aurelia auritaTakao et al. 2014 [55]43–535 **Pleurobranchaea maculata (Quoy and Gaimard, 1832) (as Pleurobranchaea novaezealandiae) Aurelia sp.1Feng et al. 2017 [70]393.8Nemopilema nomurai, Aurelia coerulea, Rhopilema esculentumTang et al. 2021 [63]27–78 Sakuraeolis enosimensis (Baba, 1930)Chrysaora quinquecirrhaTakao et al. 2014 [55]8–45 Aurelia sp.1Feng et al. 2017 [70]26.6Sakuraeolis sakuracea (Y. Hirano, 1999)Chrysaora quinquecirrhaTakao et al. 2014 [55]26–131 **^*^Predation rates are provided as polyps day^−1^, unless otherwise stated. **Dependent on body size
Morphological identification of prey nematocysts in the cnidosac of C. militaris
C. militaris nudibranchs were offered polyps of various Scyphozoa and Hydrozoa as prey, and the incorporation of nematocysts into the cnidosac was examined (see Supp. Table S1 for details). When nudibranchs were immersed in 5% KCl to clear cnidosac from previously incorporated nematocysts, the nudibranchs recoiled and raised their cerata, ejecting a mix of mucus and nematocysts without autotomizing their cerata. Ten minutes after returning to seawater, the animals resumed normal behavior. While the cnidosacs of freshly collected nudibranchs, at sea, contained hundreds of nematocysts (Fig. 2a), most cnidosacs examined after the KCl treatment were completely empty of nematocysts and in others, between 1 and 3 nematocysts were detected (Fig. 2b).Fig. 2. Cnidosacs in the cerata of C. militaris nudibranchs. a Naturally feeding b After treatment with 5% KCl. c Fed with polyps of the scyphozoan, R. nomadica on day 12. d Fed with the hydrozoan P. disticha on day 9
Following cnidosac clearing, C. militaris preyed on all prey species provided during the 12-day feeding experiment (supp. Table S2). However, the cnidosacs of nudibranchs that fed on the polyps of Aurelia sp., C. andromeda, and R. nomadica, and on the hydrozoan Nemalecium lighti showed no signs of nematocyst incorporation during the experiment at days 3, 6, 9 and 12 (Fig. 2c shows a ceras of C. militaris that fed on R. nomadica polyps). In contrast, nudibranchs feeding on P. disticha, used as a control due to its established role as a natural prey species [14, 52], incorporated the prey nematocysts into their cnidosacs within 3–6 days. Incorporation of hydrozoan nematocysts increased temporally, until by day 9 of the experiment, several hundred hydrozoan nematocysts of various types were concentrated in each of the cnidosacs examined (Fig. 2d).
The nematocysts found in the cnidosacs of P. disticha fed nudibranchs were identified as stenoteles (large, medium, and small), desmonemes, and microbasic mastigophores (large and small) (Fig. 3a). The desmonemes were the most abundant (35.9%), and the large microbasic mastigophores, least abundant (1.5%) (Table 3). The same types and sizes of nematocysts were identified in the hydrozoan, P. disticha [71] (Fig. 3b–f, Table 3). However, the relative abundance of the various nematocyst types in P. disticha was significantly different than that found in the cnidosacs of nudibranchs fed with this hydrozoan (PERMANOVA, R^2^ = 0.56, p < 0.0001). The small stenoteles (Fig. 3a) were the main contributors to the differences observed between the nematocysts in P. disticha and the cnidosac samples (SIMPER, 39%); only the desmonemes exhibited similar relative abundances in the cnidosacs and in the Hydrozoa.Fig. 3. Similar nematocyst types observed in the cnidosacs of C. militaris feeding on the Hydrozoa P. disticha, and occurring in P. disticha a Nematocysts in the cnidosacs of C. militaris feeding on P. disticha (day 9). SS, small stenoteles; MS, medium stenoteles; LS, large stenoteles; LMM, large microbasic mastigophores; SMM, small microbasic mastigophores; D, desmonemes. b–f Discharged and intact nematocysts types observed in P. disticha. b–d Small, medium, and large stenoteles, e Microbasic mastigophores, f DesmonemesTable 3Measurements and relative abundance of the various intact nematocyst types found in the cnidosacs of C. militaris nudibranchs after 9 days of preying on the hydrozoan P. disticha and in P. distichaCnidosacs of C. militarisHydrozoan P. distichaNematocystLength (µm)Width (µm)AbundanceRelative abundance (%)Length (µm)Width (µm)AbundanceRelative abundance (%)Small stenotele7.3 ± 0.15.1 ± 0.223.1 ± 14.418.2 ± 9.86.6 ± 0.65.2 ± 0.573.0 ± 15.957.0 ± 10.5Medium stenotele13.6 ± 0.88.9 ± 0.714.6 ± 8.111.9 ± 6.613.7 ± 1.69.5 ± 0.85.3 ± 8.54.1 ± 6.8Large stenotele30.1 ± 2.515.9 ± 1.222.7 ± 9.519.0 ± 8.629.0 ± 2.116.1 ± 2.35.3 ± 8.02.3 ± 3.2Desmoneme5.3 ± 0.13.5 ± 0.143.0 ± 16.435.9 ± 14.55.1 ± 0.24.0 ± 0.234.6 ± 15.827.1 ± 12.5Small microbasic mastigophores7.23 ± 0.53.8 ± 0.315.3 ± 10.313.3 ± 10.110.1 ± 0.84.5 ± 0.53.0 ± 4.62.3 ± 3.7Large microbasic mastigophores12.1 ± 0.56.3 ± 0.41.9 ± 2.31.5 ± 2.011.1 ± 1.16.0 ± 0.58.6 ± 4.01.8 ± 3.1Measurements are presented as mean ± SD (n = 10) and relative abundance is presented as percent of > 100 nematocysts counted (n = 7)^*^Smaller size microbasic mastigophores (n = 6, 7.2 ± 0.9 X 3.0 ± 0.5), similar to the ones found in the cnidosacs were also detected in the hydrozoan samples. However, these were very rare as previously described by Östman [71]
A comparison between nudibranchs in the feeding experiments and wild-caught animals showed that the cnidosacs of wild-caught nudibranchs contained various nematocyst types and sizes (Supp. Fig. S4). Many of the nematocysts found in cnidosacs of wild-caught nudibranchs were small, medium, and large stenoteles (sizes: 5–9 × 4–8 µm, 13–17 × 7–14 µm, and 22–26 × 14–20 µm, respectively), similar in size and shape to the cnidome described above in the cnidosacs of nudibranchs feeding on the hydrozoan P. disticha. The most abundant and distinct nematocysts found were bean-shaped microbasic euryteles (ca. 10–16 × 5–8 µm) with a short shaft, divergent from the lateral capsule axis, matching the nematocyst complement of the hydrozoan, Eudendrium merulum (Watson, 1985) [72]. Additional nematocysts found only in the cnidosacs of naturally feeding C. militaris were round stenoteles (9–12 × 8–11 µm) and small isorhizas (5.5–6.5 × 4–5 µm).
Discussion
The aeolid nudibranch, C. militaris, has previously been reported to feed on several hydrozoan species [14, 52]. Our study showed that C. militaris is a generalist and can also prey on various scyphozoan polyps of common eastern Mediterranean species (Supp. videos S1–S3; Table S5), and that these polyps can serve as an exclusive and sustained food source over extended periods.
In the feeding experiments C. militaris did not consume all the available polyps even at low prey levels (< 20 polyps d^−1^). The feeding rate of the nudibranch was not constant and appeared to increase with higher food availability. Density-dependent feeding behavior, where predation rate accelerates in response to increased availability of prey, up to a saturation point, has been observed in many invertebrates as well as vertebrates [73, 74]. A similar functional response was observed in the nudibranch Coryphella verrucosa, demonstrating density-dependent feeding while not consuming 100% of the prey, even at low prey densities [7, 75]. In our experiments, the predation rate of C. militaris did not reach saturation even when provided with > 80 polyps d^−1^. Therefore, the maximum rate of 95 polyps d^−1^ reported here is likely an underestimation of its predatory capacity. The predation rate of C. militaris on jellyfish polyps is comparable to rates reported for other nudibranch species (see Table 2), suggesting that it may significantly impact polyp populations in situ through predation [7, 9, 53]. Nevertheless, since these nudibranchs appear to be opportunistic predators [7], and scyphozoan polyp populations may be patchy, predicting the ecological impact of C. militaris on polyp populations in the wild remains challenging.
Like other nudibranch species, C. militaris does not graze the podocysts of R. nomadica [55, 65, 70, 76]. Thus, podocysts may serve as a protective stage, enabling polyp population recovery even after grazers have substantially reduced polyp abundances [77].
While C. militaris appeared unaffected by hydrozoan nematocysts, it visibly retracted upon contact with scyphopolyp nematocysts, yet continued feeding. In contrast, contact with anemone nematocysts prompted full retreat. C. militaris exhibited a response to scyphozoan polyps attack similar to that of Cratena pilata, a known predator of polyps of the scyphozoan Chrysaora quinquecirrha (Desor, 1848) in Chesapeake Bay, which has been recorded recoiling upon contact with the polyps’ tentacles yet consuming these nonetheless [65]. These differences in nudibranch responses to prey contact may be influenced by both prey- and predator-related factors. On the one hand, nematocyst toxins vary considerably among prey classes [61]; on the other hand, nudibranchs may secrete prey-specific mucus that inhibits nematocyst discharge [24].
Many aeolid nudibranchs sequester nematocysts from their cnidarian prey for use as defense against predators [13, 21, 23]. This incorporation is selective, influenced by species, prey choice, and predation pressure on the nudibranchs [27, 29, 30, 46, 62–64]. Analysis of cnidosacs from freshly-collected nudibranchs revealed that C. militaris exhibits a generalist feeding behavior, such that, in addition to a nematocyst complement consistent with that of the hydrozoans P. disticha and E. merulum [71, 72], multiple nematocyst types and sizes were observed. Laboratory experiments confirmed that when C. militaris fed on P. disticha, it retained all of the hydrozoan's nematocyst types in its cnidosac [71]. However, the proportions of these nematocysts differed markedly from those in the hydrozoan itself and the incorporated small-microbasic mastigophores were notably smaller (Table 3). In contrast, the nudibranch C. pilata incorporated only microbasic mastigophores from the same hydrozoan [34]. Similar patterns of selective nematocyst incorporation have been documented in many other nudibranch species [25, 27, 29, 30, 33–35, 78]. Our experiments clearly demonstrate that C. militaris selectively incorporates nematocysts of specific types or sizes from the hydrozoan P. disticha, and when provided with scyphozoan polyps the nudibranch did not sequester any of the prey nematocysts.
Some nudibranchs rely on cnidarian and non-cnidarian prey for different purposes; the non-cnidarian prey provides nourishment and the cnidarian prey a source of nematocysts for defense [32]. In Phyllodesmium spp., however, the cnidosac is non-functional and these aeolids rely instead on secondary metabolites from their cnidarian prey rather than incorporating kleptocnides for defense [79]. Here we show, for the first time, that C. militaris, despite possessing a functional cnidosac, utilizes scyphozoans exclusively as a food source, disregarding their defensive nematocysts. This distinct strategy of diversified prey consumption yet exclusive nematocyst incorporation is unique among nudibranchs.
C. militaris has only recently been introduced into the eastern Mediterranean and is now regularly observed during winter and spring [50]. Therefore, the predation preferences and long-term ecological effects of this species on jellyfish polyp populations require further study. Given the role of scyphopolyps as pivotal contributors to jellyfish proliferations [6], documenting predator–prey interactions between nudibranchs and scyphopolyps is essential to our understanding of medusan population dynamics.
Conclusions
We found that in addition to feeding on hydrozoans, C. militaris preys on a range of scyphozoan polyps including Aurelia sp., C. andromeda, P. punctata, and R. nomadica, and can rely solely on these, as food for extended periods of time. Despite its efficient consumption of scyphopolyps, C. militaris does not incorporate their nematocysts into the cnidosac, displaying a distinct selectivity in nematocyst sequestration. When prey is abundant, this nudibranch exhibits high predation rates, exceeding 95 polyps d^−1^, indicating its potential to influence jellyfish population dynamics by reducing polyp numbers.
Supplementary Information
Supplementary Material 1Supplementary Material 2Supplementary Material 3Supplementary Material 4Supplementary Material 5
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Restaino D. Molecular ecology and management of nuisance and invasive marine species: Chrysaora chesapeakei, Moerisia sp., and Gonionemus vertens; 2018.
- 2Dror H, Angel DL. Substrate preferences and the effect of temperature on planulae settlement of the scyphozoan Rhopilema nomadica. Hydrobiologia. 2025; 1–14.
- 3Vogel RM. Motion picture of the courtship and early embryology of the eolid nudibranch Cratena pilata Gould. Bull Am Malacol Union. 1969;18–9.
- 4Sorrano AL. Settling patterns of Chrysaora quinquecirrha polyps on common vinyl construction material: potential implications for jellyfish blooms and coastal development in Barnegat Bay New Jersey. Montclair State University; 2016.