A minimal ion–chemistry model for predicting benign paroxysmal positional vertigo risk based on endolymphatic calcium and pH
Dong-Gyun Han

TL;DR
This paper introduces a new model to predict the risk of benign paroxysmal positional vertigo by analyzing endolymph calcium and pH levels.
Contribution
The study presents a novel biochemical model integrating calcium and pH to predict otoconia stability and BPPV risk.
Findings
The model uses endolymph saturation index (Ω) to determine otoconia stability or dissolution.
Systemic factors like pH and calcium concentration influence Ω and BPPV risk.
Simulations show a right-skewed risk distribution with most individuals having a high relative risk.
Abstract
Benign paroxysmal positional vertigo (BPPV) arises from detachment of otoconia—calcium carbonate (CaCO3) crystals embedded in a protein matrix—whose stability depends on endolymph ionic composition and pH. Age-related calcium metabolism, acid–base imbalance, and hormonal factors can impair otoconia integrity, yet, to our knowledge, no prior quantitative model integrates these biochemical parameters to predict BPPV risk. Beyond the established mechanical mechanisms of canalithiasis and cupulolithiasis, we introduce a parsimonious biochemical model in which the endolymphatic saturation index (Ω), governed by pH and ionized calcium [Ca2+], delineates an otoconia stability–dissolution boundary (Ω≈1) and complements the mechanical framework. Using carbonate-equilibrium chemistry and the CaCO3 solubility product (Ksp), we compute Ω and derive the critical calcium concentration Ccrit(pH) at Ω…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
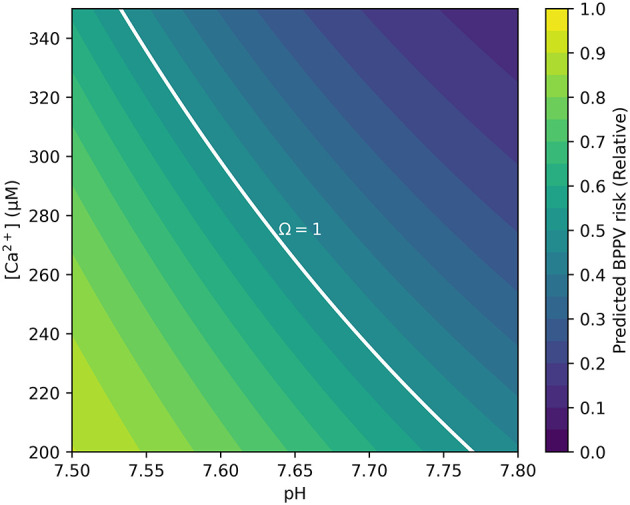 Figure 1
Figure 1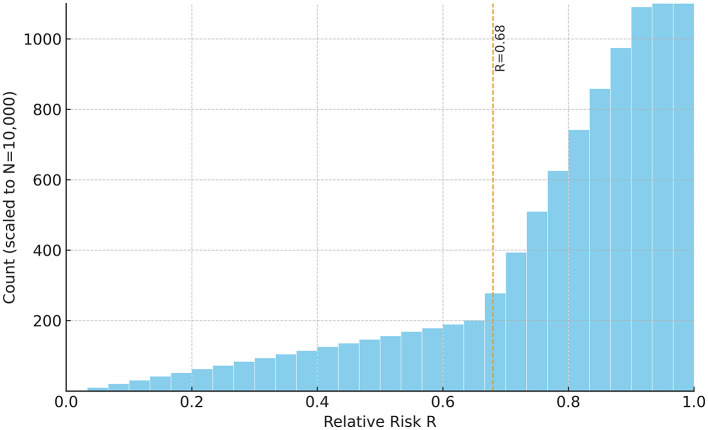 Figure 2
Figure 2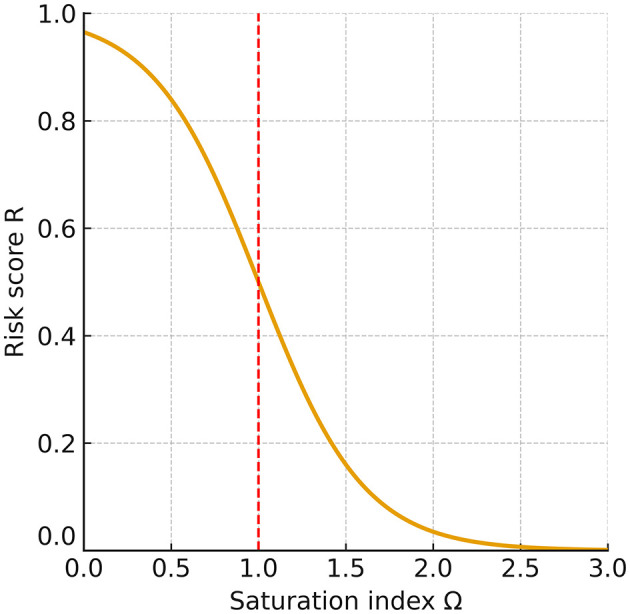 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVestibular and auditory disorders · Glaucoma and retinal disorders · Olfactory and Sensory Function Studies
1 Introduction
Benign paroxysmal positional vertigo (BPPV) is the most common peripheral vestibular disorder, with a lifetime prevalence of ~2.4% (1). It results from displacement of otoconia from the utricular macula into the semicircular canals (2, 3). Otoconia are biomineralized structures composed mainly of calcium-carbonate (CaCO_3_) crystals embedded in an organic (proteinaceous) matrix (4, 5). In contrast to bone mineral—largely hydroxyapatite Ca10(PO4)6(OH)2, a calcium phosphate with a highly stable lattice and strong ionic bonding—calcite CaCO_3_ exhibits weaker ionic bonding and greater chemical reactivity. This lower intrinsic stability renders otoconia more vulnerable to dissolution under acidic conditions or shifts in ionic composition, whereas hydroxyapatite confers long-term structural integrity to bone.
Otoconia stability is critically modulated by calcium metabolism and endolymph pH (6, 7). Age-related reductions in intestinal calcium absorption, estrogen deficiency, and systemic disorders such as osteoporosis, migraine, and Ménière's disease perturb endolymphatic calcium homeostasis (8–12). Endolymph pH, governed by systemic respiratory and metabolic acid–base balance, sets carbonate speciation; acidification shifts the equilibrium toward bicarbonate/dissolved CO_2_, increasing CaCO_3_ solubility. Experimental and clinical reports indicate that pH reduction—whether due to metabolic acidosis, hypoventilation, or local inflammation—destabilizes otoconia and increases the likelihood of BPPV episodes (12, 13).
Prior models have emphasized biomechanical factors, otolithic-membrane integrity, and systemic comorbidities (14, 15), but none have quantitatively linked the biochemical determinants—endolymphatic free calcium [Ca^2+^]and pH —to BPPV risk. Here, we present a carbonate saturation index–based model in which pH and [Ca^2+^] jointly determine the stability–dissolution boundary (Ω≈1). This framework complements (rather than replaces) the mechanical paradigm by providing a quantitative map from systemic conditions to predicted otoconia stability and BPPV risk.
2 Methods
2.1 Model formulation
We quantified otoconia stability using the carbonate saturation index (Ω):
Here, a denotes activity (ai = γ_i[i]); γi_ are activity coefficients (dimensionless) and [i] are molar concentrations. Carbonate is obtained from pH-dependent speciation as , where CT is total inorganic carbon. Ksp is the effective solubility product of calcite CaCO_3_ under the prevailing temperature and ionic-strength conditions. For a minimal model at near-physiological ionic strength (I ≈ 0.15–0.20 M) and 37 °C, we approximate activities by concentrations (γ≡1) and treat Ksp as an effective constant; thus Ω>1 denotes supersaturation favoring precipitation, whereas Ω < 1 denotes undersaturation favoring dissolution. This formulation follows diffusion-controlled CaCO_3_ crystallization (16) and biomineralization studies (17).
Numerator— :
[Ca^2+^] is determined by the balance between trans-epithelial calcium influx, efflux, passive diffusion, and protein binding within the vestibular endolymphatic compartment (referring to soluble proteins in endolymph that reversibly bind Ca^2+^, distinct from the structural matrix of otoconia). The term is derived from total inorganic carbon (CT) and the pH-dependent dissociation fraction α_2_, such that:
Even small changes in pH can produce disproportionately large shifts in because α_2_ contains an exponential term, making pH a sensitive determinant of the numerator.
Denominator—Ksp:
The Ksp term is the solubility product constant for CaCO_3_ at body temperature and ambient pressure, representing the thermodynamic equilibrium condition at which precipitation and dissolution rates are balanced. While temperature and pressure are stable in vivo, ionic strength alters ion activities, resulting in a larger effective Ksp when expressed in concentrations. In biological fluids, macromolecules, chelators, and changes in protein composition can also modify the effective Ksp by altering ion activities.
Under stable temperature and pressure, variations in Ω are primarily driven by changes in [Ca^2+^], pH (via ), CT, and ionic strength.
Carbonate speciation and dissociation fractions:
At 37 °C, carbonate species distribution is determined by the first (pKa1≈6.12) and second (pKa2≈10.3) dissociation constants of carbonic acid, together with the CO_2_ solubility coefficient in plasma (SCO2≈ 0.0306 mmol L mmHg at 37 °C) (18). Defining:
the normalization constant
represents the sum of equilibrium terms for dissolved CO_2_, bicarbonate, and carbonate ions, and serves as the common denominator in the fractional speciation expressions:
where α_0_, α_1_, and α_2_ are the fractions of total inorganic carbon (CT) present as dissolved [CO2(aq)], bicarbonate ions , and carbonate ions , respectively. Total inorganic carbon is defined as:
When is available (e.g., from blood gas analysis), CT can be obtained from
2.1.1 Relative risk score (R)
The relative risk score was calculated by applying a logistic transformation to the deviation of measured [Ca^2+^] from the critical concentration Ccrit(pH ):
where σ_C_ is a dispersion parameter representing inter-individual variability in tolerance to deviations of [Ca^2+^] from Ccrit. A smaller σ_C_ produces a steeper logistic curve, indicating high sensitivity to small departures from Ccrit, whereas a larger σ_C_ yields a shallower curve, reflecting lower acute sensitivity but a greater probability of prolonged residence in destabilizing states. Biologically, σ_C_ captures heterogeneity in physiological tolerance, arising from factors such as genetic variation, hormonal status, comorbidities, age, and environmental influences. Empirical estimation of σ_C_ can be performed by measuring [Ca^2+^] and pH in a representative cohort, calculating Ccrit for each individual, and fitting the resulting risk distribution to the logistic function–based risk model using regression analysis or maximum likelihood estimation.
2.1.2 Relative critical calcium concentration index
For comparative purposes under constant Ksp, we defined a dimensionless index:
This index allows prediction of relative shifts in Ccrit between physiological and pathological states without invasive sampling of endolymph.
Systemic acid–base disturbances measured in blood predictably shift Ccrit, thereby linking systemic physiology to inner-ear carbonate equilibrium. For example, acidosis (pH↓, pCO_2_↑) decreases α_2_, shifting carbonate speciation toward CO2/ dominance, lowering , and raising Ccrit, favoring dissolution (Ω < 1). In contrast, alkalosis increases and lowers Ccrit, promoting supersaturation (Ω>1). Ionic-strength effects computed with Davies activity coefficients (γ) are illustrated in Supplementary Figure S1. These examples highlight, in a qualitative manner, how systemic biochemical states may alter otoconia stability. The framework should be regarded as hypothesis-generating, serving to guide future studies rather than as a validated clinical tool.
2.1.3 Critical calcium at the saturation boundary
We start from the activity-based definition of the saturation index:
At the precipitation–dissolution boundary (Ω = 1), the critical free calcium is
Using (Equation 2; α_2_ given in Equation 5), we obtain
2.1.4 Interpretation
Acidification lowers α_2_(pH) and/or reductions in CT decrease , thereby raising Ccrit(pH) (more [Ca^2+^] needed to keep Ω = 1).
2.2 Mapping of systemic factors
We conducted a targeted literature review to identify systemic and local conditions that influence endolymph [Ca^2+^] and pH, such as calcium homeostasis disorders, acid–base disturbances, endolymphatic pathological processes, genetic variants, and pharmacological agents (19–33). The mechanistic implications of these factors for CaCO_3_ saturation and otoconia stability are further elaborated in Section 4.2 of the Discussion.
2.3 Simulation study
We conducted an in silico simulation to examine how endolymph chemistry modulates otoconia stability. A cohort of N = 10,000 virtual individuals was generated. Free calcium values were drawn within 200–350 μM and pH within 7.5–7.8, with these windows anchored to vertebrate vestibular reports of [Ca^2+^]≈ 250–280 μM and pH ≈ 7.6–7.7 (7, 34–37) and extended to encompass plausible pathological deviations related to barrier dysfunction, inflammation, or systemic acid–base disturbance. The total inorganic carbon CT was calibrated so that Ccrit (7.65) = 265 μM, providing a physiologically anchored reference point. Unless otherwise stated, the dispersion parameter in the risk mapping was fixed at σ_C_ = 80 μM. To reproduce the empirically right-skewed risk shape with mean R≈ 0.68 while keeping the same windows and σ_C_, the calcium draw within 200–350 μM was implemented with a mild asymmetry (details and code are provided in Supplementary Data S1); no biological sampling was performed.
For each simulated case, the pH-dependent critical calcium concentration Ccrit(pH) was computed from carbonate equilibrium chemistry (pK1 = 6.1, pK2 = 10.3 at ~37 °C; Ksp = 4.47 × 10^−9^). The saturation index was then evaluated as Ω = ( )/Ksp; in the minimal model we set activities to unity. Finally, a dimensionless relative risk score was obtained by mapping the calcium distance to threshold through a logistic transform, R = 1/{1 + exp (([Ca^2+^] – Ccrit(pH))/σ_C)}. By this convention, R → 1 indicates undersaturation ([Ca^2+^] < Ccrit, Ω < 1) and a dissolution-prone state, whereas R → 0 indicates supersaturation ([Ca^2+^] > Ccrit_, Ω>1) and relative mineral stability.
Two primary outputs were produced: (i) a risk heatmap on the pH[Ca^2+^] plane—computed with CT, T, I, and Ksp held at physiological reference values with targeted sensitivity analyses—that visualizes Ω and its Ω = 1 boundary separating stable (Ω>1) from unstable (Ω < 1) regions (Figure 1); and (ii) a 30-bin histogram of R summarizing the population distribution (Figure 2). The histogram uses a count axis labeled 0–1,000, with the tallest bar slightly exceeding 1,000, making the right-skew explicit. Under the baseline windows and σ_C_ = 80 μM, the distribution is right-skewed (mean ≈0.78; 80% with R>0.68), consistent with many cases lying just below the saturation boundary near the calibration point (pH 7.65, Ccrit = 265 μM). The non-linear pH-dependence of Ccrit and the logistic mapping further accentuate this skew.
Contour map showing BPPV risk as a function of endolymph pH and free calcium [Ca2+]. The solid white curve denotes the Ω = 1 dynamic-equilibrium boundary, at which precipitation and dissolution of CaCO3 are balanced. Regions above this boundary (Ω>1) represent supersaturation and relative otoconia stability, whereas regions below the boundary (Ω < 1) indicate undersaturation and increased dissolution risk. Simulation parameters covered the physiological range of animal vestibular endolymph (250–280 μM [Ca2+], pH 7.6–7.7) and were extended to 200–350 μM and pH 7.5–7.8 to encompass plausible pathological deviations. Risk is displayed on the pH[Ca2+] plane; CT, T, I, and Ksp are held at physiological references; sensitivity analyses are provided in Supplementary Figures.
Histogram of simulated relative BPPV risk (R) for the synthetic cohort (N = 10,000). R was computed via Equation 8 with σC= 80 μM (i.e., R = 1/[1+exp(−ΔC/σC)], ΔC≡Ccrit(pH)-[Ca2+]). Higher R ( → 1) indicates undersaturation ([Ca2+] < Ccrit; Ω < 1) and greater dissolution propensity, whereas lower R ( → 0) reflects supersaturation ([Ca2+] > Ccrit; Ω>1) and relative mineral stability. The distribution is right-skewed (mean ≈ 0.78; 80% of cases with R> 0.68). The count axis is labeled 0–1,000, and the tallest bar slightly exceeds 1,000. Sampling windows: [Ca2+] 200–350 μM; pH 7.5–7.8; calibration: Ccrit(7.65)= 265 μM. Interpreting the metrics: a cohort mean of R≈0.78 implies that, on average, free calcium lies roughly 100μM (≈10−4M) below Ccrit(pH); moreover, R>0.68 corresponds to ΔC ≳ 60 μM (i.e., [Ca2+]-Ccrit≲-60μM), consistent with a majority occupying the undersaturated (Ω < 1) regime.
Robustness was assessed in a consolidated sensitivity analysis (Supplementary Figure S2). Expanding the windows to [Ca^2+^] 180–380 μM or pH 7.4–7.9 did not move the Ω = 1 boundary (only the plotted axes were extended). Varying σ_C_ between 40 and 120 μM steepened or flattened the logistic mapping of R without shifting Ω = 1, consistent with the boundary's independence from the logistic transform. In contrast, scaling CT by ±10% displaced the Ω = 1 boundary modestly while preserving the qualitative risk-contour structure. For completeness, ionic-strength effects computed with Davies activity coefficients at ~37 °C are shown in Supplementary Figure S1: increasing I (0.15, 0.20, 0.30 M) lowers γ and shifts the Ω = 1 boundary upward (higher [Ca^2+^] required to maintain Ω = 1) across pH 7.4–7.9, while the qualitative topology is preserved.
2.3.1 Reproducibility
All analyses were performed in silico; the synthetic cohort (N = 10,000) and plotting code are provided as Supplementary Data S2 and reproduce Figure 2 under the parameters described herein (CT calibrated so that Ccrit (7.65) = 265 μM; σ_C_ = 80 μM; pH 7.5–7.8; [Ca^2+^] 200–350 μM).
2.4 Core mathematical method
We formulate a deterministic, three-step mapping from (pH, [Ca^2+^]) to (Ω, R); no empirical training is required.
Step 1—Carbonate speciation. Given pH, carbonate dissociation constants (pK1, pK2) define the fractional species α_0_(pH), α_1_(pH), α_2_(pH). The carbonate available for precipitation is
where CT is total inorganic carbon (see Section 2.1 for definitions and constants).
Step 2—Saturation index. The carbonate saturation index is
Step 3—Logistic risk mapping.
Let
The probability-like, dimensionless risk score is
so that R = 0.5 at ΔC = 0 (i.e., Ω = 1), R → 1 for ΔC>0 (undersaturation), and R → 0 for ΔC < 0 (supersaturation).
2.4.1 Calibration and separation of roles
For simulations we scale CT so that Ccrit(pH = 7.65) = 265μM (physiological anchor). The CO_2_ solubility coefficient is not used in theoretical simulations; it is invoked only in the clinical translation pipeline when inferring CT from blood-gas variables. This separation keeps the theoretical equations transparent while clarifying how clinical inputs are mapped onto the same framework.
2.4.2 Clinical translation using a blood-derived proxy
To validate directional and temporal predictions without endolymph sampling, we define a blood-derived Ω proxy from serum ionized calcium and blood-gas . The proxy is intended to test associations (e.g., BPPV status) and within-person phase concordance, not to estimate absolute endolymph states. All claims of clinical applicability are therefore conditional on prospective calibration of blood– endolymph relationships.
3 Results
The model predicted that systemic factors influencing pH or [Ca^2+^] produce distinct shifts in the carbonate saturation index (Ω) and thereby alter otoconia stability. Increases in pH–as in hyperventilation, prolonged vomiting, or with thiazide diuretics—elevated Ω. Increases in [Ca^2+^]–e.g., in hyperparathyroidism or vitamin D excess—also raised Ω. Conversely, decreases in pH–as in metabolic or respiratory acidosis, and with acetazolamide—lowered Ω. Loop diuretics reduced [Ca^2+^] and, when combined with volume depletion, further depressed Ω. Likewise, hypocalcemic states (e.g., vitamin D deficiency or hypoparathyroidism) reduced Ω.
Simulation outputs were consistent with these predictions. The risk map showed that low pH combined with low [Ca^2+^] markedly expanded the Ω < 1 region (Figure 1). The Ω = 1 contour cleanly delineated the transition between stable (Ω>1)and unstable (Ω < 1) states. The histogram of relative risk scores (R) was right-skewed with mean ≈0.78, and 80% of the cohort had R>0.68, indicating that a large fraction of the synthetic population resides near or below the saturation boundary (Figure 2). Consistent with the plotting parameters of Figure 2, the count axis was labeled 0–1,000 and the tallest bar slightly exceeded 1,000, making the right-skew visually explicit.
To aid clinical interpretation, we also examined a blood-derived proxy: hypocalcemia (e.g., ionized calcium ≲ 0.96 mM, with severe cases ~0.68 mM) or acidemia (pH 7.30–7.35) typically corresponds to Ω < 1 under the minimal model assumptions and therefore maps to high R. This proxy is illustrative rather than a direct measurement of endolymph chemistry, and we emphasize that definitive inferences require vestibular- compartment data.
Further shifts of the Ω = 1 boundary under Ksp perturbations (±10–20%) and temperature (35–39 °C; with pK1(T), pK2(T), and Ksp(T) updated) are provided in Supplementary Figures S3, S4. These analyses preserved the qualitative topology of the boundary while moving its position modestly. A consolidated sensitivity analysis (Supplementary Figure S2) showed that broadening the sampling windows to [Ca^2+^] 180–380 μM or pH 7.4–7.9 did not move Ω = 1 (only the plotted axes enlarged). Varying σ_C_ between 40 and 120 μM steepened or flattened the logistic mapping from threshold distance to R without shifting Ω = 1, consistent with the boundary's independence from the risk transform.
Reproducibility. All analyses were performed in silico; the synthetic cohort (N = 10, 000) and plotting code are provided as Supplementary Data S2 and exactly regenerate Figure 2 using the baseline parameter set described in Section 2.3.
4 Discussion
Human otoconia are calcite-based CaCO_3_ biominerals, so the carbonate saturation-index framework used in crystallization and biomineralization applies naturally to the vestibular system. We therefore adopted the physically interpretable saturation index Ω (Equation 1) as the core descriptor of the precipitation–dissolution balance and evaluated it under endolymphatic conditions (16, 17). Although many marine skeletons (e.g., corals, mollusks) precipitate aragonite rather than calcite, the Ω formalism remains applicable; differences arise through Ksp and kinetic pathways. In this study, Ksp was treated as an effective constant at ~37 °C and physiological ionic strength, and—in the minimal model—activities were approximated by concentrations. Potential departures from ideality (e.g., organic-matrix mediation, non-ideal solution effects) are addressed in Section 4.6 (Limitations and Future Directions).
Building on this formulation, we constructed a minimal ion–chemistry model that integrates endolymphatic [Ca^2+^] and pH into a quantitative assessment of BPPV risk. Expressing otoconia stability through Ωcaptures the dynamic balance between CaCO_3_ precipitation and dissolution, the principal mineral process governing otoconia. Because carbonate speciation links Ω non-linearly to [Ca^2+^] and pH, even modest perturbations can drive the system toward net dissolution (Ω < 1) or net precipitation (Ω>1; Figure 3). Although endolymph is generally maintained in a supersaturated state (Ω>1), its buffering capacity is lower than that of cerebrospinal fluid, rendering it more vulnerable to systemic or local disturbances in acid–base balance or calcium homeostasis (38). Consistent with this susceptibility, our simulations produced a right-skewed distribution of risk scores R (Figure 2) and clear, interpretable Ω = 1 boundaries in the pH[Ca^2+^] plane (Figure 1), linking biochemical shifts directly to predicted mechanical stability of otoconia.
Relationship between the saturation index (Ω) and risk score (R). At the equilibrium point (Ω = 1), the risk is R = 0.5. When Ω falls below 1, R rises steeply toward 1, indicating a dissolution-prone state for otoconia and a higher likelihood of recurrent or persistent BPPV; when Ω exceeds 1, R declines toward 0, reflecting a stable condition in which otoconia are preserved. This monotonic relationship shows that even a modest reduction of Ω below 1 sharply increases the predicted risk.
4.1 Mechanistic implications
At Ω = 1, precipitation and dissolution of CaCO_3_ are in dynamic equilibrium, so otoconial mass is preserved on average while ion exchange continues. The system is highly sensitive: small changes in pH or [Ca^2+^] can push it toward net precipitation (Ω>1) or net dissolution (Ω < 1), with disproportionate effects on otoconia integrity and detachment risk (Figure 3). The equilibrium calcium threshold Ccrit(pH) defined in Equation 10 makes this dependence explicit: modest acidification (which lowers α_2_) or reduced calcium availability readily depresses Ω below unity, predisposing to instability. In our risk mapping (Equation 8), this appears as : ΔC>0 yields R near 1 (undersaturation), whereas ΔC < 0 yields R near 0 (supersaturation).
The inherent chemical vulnerability of otoconia arises from the weaker ionic bonding of calcite CaCO_3_ compared with hydroxyapatite Ca10(PO4)6(OH)2. Hydroxyapatite has a highly stable lattice and very low solubility product, rendering it resistant to physiological pHfluctuations. By contrast, CaCO_3_ dissolves more readily as pH decreases because carbonate equilibria shift toward and dissolved CO2. Utricular endolymph is typically ~ pH 7.6–7.7 (7, 34–37), so even mild acidotic shifts can depress Ω below unity and raise CaCO_3_ solubility.
4.2 Convergence of etiologies and clinical alignment
A wide range of systemic and local conditions ultimately converge on a single chemical pathway that governs otoconia stability: modulation of the carbonate saturation index, Ω. Disturbances of calcium homeostasis—whether from hyper- or hypocalcemia, vitamin D deficiency or excess, or hyper-/hypoparathyroidism—shift free endolymphatic [Ca^2+^] and thereby alter Ω (19–22). Acid–base disorders operate through carbonate speciation α_2_(pH): metabolic or respiratory acidosis lowers pH, reduces α_2_, and depresses Ω, whereas alkalosis has the opposite effect (23, 24). Disease processes such as vestibular migraine, Ménière's disease, autoimmune inner-ear disease, and viral inner-ear disorders can change endolymph composition and destabilize otoconia (25, 26). Genetic factors encompass channelopathies and transporter defects affecting endolymph homeostasis and migraine susceptibility. Variants in CACNA1A (P/Q-type voltage-gated Ca^2+^ channel α1A subunit) and ATP1A2 (Na^+^/K^+^-ATPase α2 subunit) primarily alter Ca^2+^ handling in vestibular epithelia, leading to changes in free endolymphatic [Ca^2+^] and otoconia stability. Mutations in SLC26A4 (pendrin, exchanger) and SLC4A11 (H^+^/OH^−^ transporter) predominantly affect pH and bicarbonate balance, shifting carbonate equilibrium (α_2_) and thereby indirectly influencing CaCO_3_ saturation (Ω) (27–31). Pharmacological agents also steer Ω in predictable directions: thiazides tend toward metabolic alkalosis and raise Ω; acetazolamide induces metabolic acidosis and lowers Ω; loop diuretics reduce [Ca^2+^] and, with volume depletion, further depress Ω (32, 33).
Viewed through Equation 1, these influences either decrease the activity product (for example by lowering [Ca^2+^] or α_2_(pH), or by increasing ionic strength and thereby lowering activity coefficients during labyrinthitis, autoimmunity, or barrier breakdown), or else increase it and move the system away from the dissolution threshold (22, 39–47). Repetitive neurogenic inflammation in vestibular migraine or Ménière's disease may additionally disrupt Ca^2+^ transporters in dark cells, transiently elevating free [Ca^2+^]; concurrent degradation of otoconial matrix proteins then impairs lattice incorporation and weakens stability (26, 48). Combinations of insults are particularly consequential: acidosis coupled with labyrinthitis can drive Ω well below 1 and accelerate CaCO_3_ dissolution, whereas alkalosis with elevated [Ca^2+^] can push Ω well above 1, stabilizing otoconia but potentially favoring pathological crystal aggregation. Taken together, these convergent mechanisms destabilize the protein–mineral composite of otoconia and yield fragments more prone to detachment during head movements, aligning the chemical predictions with clinical vulnerability patterns (Figure 1).
4.3 Alignment with clinical and epidemiological data
The model's predictions are consistent with a broad spectrum of clinical and epidemiological findings. Higher recurrence rates in post-menopausal women and in patients with osteoporosis reflect hormonally mediated reductions in calcium homeostasis, which also explain the greater prevalence observed in older adults and in women. Seasonal variation in BPPV incidence, particularly during winter months when vitamin D deficiency is more prevalent, further supports the predicted vulnerability of otoconia under conditions of impaired calcium absorption (49).
Using the blood-derived proxy for Ω, the model predicts that when laboratory surrogates fall outside customary reference ranges (e.g., pH ~7.30–7.35 or ionized calcium ~0.68–0.96 mM), cases often map to the dissolution-prone domain (Ω ≤ 1) with high risk (frequently R≥0.8). In our baseline mapping, 80% of the cohort had R>0.68. These thresholds emphasize that even modest systemic deviations can destabilize otoconia and are consistent with reports of persistent or recurrent dizziness in patients with metabolic or acid–base disturbances.
For comparative interpretation at fixed Ksp, we introduced a dimensionless relative critical-calcium index (Equation 9), ICcrit__∝1/(α_2_(pH)CT). Within the simulated endolymph pH range (7.5–7.8), the index is on the order of 0.06–0.20: it rises toward ~0.2 with acidosis (lower α_2_) and falls toward ~0.06 with alkalosis, providing a simple surrogate link between systemic acid–base status and inner-ear carbonate equilibrium. Importantly, this framework offers a clinically feasible bridge from standard blood-gas analysis to risk stratification without invasive endolymph sampling. Where relevant, changes in ionic strength should be interpreted as activity effects (via γ) that modify the activity product in Equation 1 rather than Ksp itself (see Supplementary Figure S1 for Davies-based illustrations).
Beyond chronic factors, the onset of BPPV after systemic illness or prolonged bed rest can be interpreted—among other mechanisms—as a plausible consequence of respiratory acidosis. Reduced ventilatory drive and CO_2_ retention during inactivity may lower blood pH, depress α_2_, and thereby lower Ω, promoting CaCO_3_ dissolution (50, 51). Immobilization-related changes in bone turnover may further perturb systemic calcium balance. Frequent recurrence after apparent resolution is likewise compatible with the notion that many patients reside near the Ω = 1 equilibrium boundary, where modest biochemical perturbations can tip the balance toward instability. Reported ranges of [Ca^2+^] and pH across these conditions align with simulation-derived zones of instability, underscoring the translational relevance of the saturation-index framework in linking mechanistic predictions to observed clinical patterns.
4.4 Clinical translation and preventive potential
Because the model is deterministic and requires only two inputs ([Ca^2+^], pH), it can be operationalized with serum ionized calcium and blood gases as non-invasive surrogates. Crucially, these blood-based variables are not one-to-one estimators of vestibular endolymph; they support relative-risk inference via a blood-derived Ω proxy, reflecting directional trends rather than absolute endolymph states. This stance follows from the practical infeasibility of direct human utricular/saccular sampling and is especially useful for individualized risk stratification in patients with recurrent BPPV or elevated baseline risk (e.g., osteoporosis, chronic kidney disease, migraine, Ménière's disease).
To facilitate translation while respecting this limitation, we pre-specify two indirect validation designs. (i) Cross-sectional case–control: test whether a blood-derived Ω proxy (from ionized calcium and blood-gas–derived , temperature-corrected to 37 °C; arterial preferentially, venous acceptable with caveats) is independently associated with prevalent BPPV after adjustment for confounders (age, sex, BMI, renal function, vitamin D/PTH, diabetes/respiratory disease, diuretic/acetazolamide/PPI use, and hydration). (ii) Repeated-measures cohort: evaluate temporal concordance between within-person fluctuations in the proxy and symptom phases (exacerbation, resolution, and post-repositioning residual dizziness) using mixed-effects models and lagged windows (e.g., ±24–48 h). In both designs, inference is limited to association and timing, not absolute endolymph chemistry.
We also commit to robustness checks around key physicochemical assumptions: activity-coefficient/ionic-strength variation (e.g., extended Debye–Hückel within Pitzer-bounded ranges), Ksp within physiologic intervals, CT estimated from rather than fixed, and temperature within clinical limits. These analyses quantify boundary shifts around Ω = 1 and test the stability of relative-risk mapping under reasonable biochemical uncertainty.
The framework provides a mechanistic lens on post-repositioning dizziness and the clinical heterogeneity of BPPV. After apparently successful canalith repositioning, endolymph may transiently reside near the metastable Ω≈1 regime, in which precipitation and dissolution are finely balanced; otoconia are neither fully stabilized nor fully dissolved, yielding residual vestibular instability perceived clinically as lingering disequilibrium rather than true vertigo (52). More broadly, BPPV spans a continuum of otoconial states, from intact crystalline otoliths to partially dissolved or fragmented particles, which helps explain absent nystagmus, atypical directions/durations of positional responses, and subtle imbalance without overt vertigo (53–55).
Bone homeostasis reflects slow hydroxyapatite turnover over months to years; by contrast, human otoconia are calcite-based, and their stability depends on the carbonate saturation index in the minimal model). Because α_2_(pH) and [Ca^2+^] modulate Ω on short timescales, transient shifts in ionized calcium and acid–base status can promptly tilt otoconia toward dissolution. Thus, while low BMD and hypocalcemic states index chronic susceptibility, ionized calcium is expected to track near-term BPPV vulnerability more closely than BMD or total calcium (22, 56)—a hypothesis pending the prospective validation outlined above.
Preventive and therapeutic implications suggested by the model include maintaining acid–base neutrality during acute illness or the post-operative period, correcting vitamin D or calcium deficiency while avoiding excessive supersaturation, and judicious pharmacologic modulation of vestibular ion transport in channelopathy settings—e.g., thiazides (alkalosis → Ω↑) or acetazolamide (acidosis → Ω↓) vs. loop diuretics ([Ca^2+^]↓ → Ω↓) (57–60). A practical advantage is that the Ω proxy is computable entirely from clinically accessible data, obviating hazardous endolymph sampling, while the pre-specified validation and robustness testing provide the necessary bridge toward eventual clinical use.
4.5 Pharmacological modulation and hydration balance
Clinically, excessive dehydration can raise ionic strength and thereby alter ion activities (via the activity coefficients, γ), which lowers the activity product in Equation 1 and—particularly under acidic conditions—can accelerate CaCO_3_ dissolution.
Within this framework, drug classes have predictable directions of effect. Thiazide diuretics tend to promote calcium retention and metabolic alkalosis, thereby increasing α_2_(pH) and raising Ω (57, 58). By contrast, acetazolamide (carbonic-anhydrase inhibition) typically induces metabolic acidosis, lowering α_2_(pH) and thus decreasing Ω–useful for fluid dynamics in Ménière's but destabilizing for carbonate chemistry in our model (59). Loop diuretics increase calciuresis and, when coupled with dehydration, reduce free [Ca^2+^] and can depress Ω (58, 60); however, their contraction alkalosis tends to increase α_2_(and Ω), so the net effect depends on fluid and electrolyte management.
Clinical application aims to maintain eubicarbonatemia and normocalcemia so that endolymph remains mildly supersaturated (Ω>1). Preferential use of alkalinizing or calcium-sparing strategies (e.g., thiazides where appropriate), adequate hydration, and correction of vitamin D and calcium deficits can mitigate dissolution risk. If loop diuretics are unavoidable, they should be paired with careful fluid/electrolyte monitoring (including ionized calcium and acid–base status). Framed this way, endolymphatic pressure control aligns with preservation of the carbonate equilibrium that supports otoconia integrity.
4.6 Limitations and future directions
This minimal model reduces inner-ear chemistry to a saturation index Ω computed from ionized calcium [Ca^2+^] and pH and maps it to a probability-like risk score via a logistic transform. Its usability relies on deliberate simplifications: Ksp is treated as an effective constant at 37 °C and physiological ionic strength; activities are approximated by concentrations; calcite is assumed as the operative polymorph (polymorph-dependent kinetics not modeled); baseline total inorganic carbon CT is held fixed; and spatial heterogeneity, time-dependent transport, and chemo–mechanical coupling (detachment forces, otolithic-membrane pathology, and aggregation) are omitted. Organic-matrix effects and explicit protein–ion interactions—known to influence nucleation, stabilization, and dissolution—are likewise not yet incorporated. These choices were intentional to provide a transparent, reproducible, and falsifiable starting point.
A central limitation is the practical infeasibility of direct human utricular/saccular endolymph sampling, precluding empirical measurement of absolute vestibular chemistry. Consequently, our framework is hypothesis-generating, and all clinical inferences are restricted to what can be supported by blood-based surrogates. Specifically, we construct a blood-derived Ω proxy—from serum ionized calcium and blood-gas–derived /pCO_2_–to test directionality (association with BPPV) and temporal concordance (within-person fluctuation vs. symptom phases), not to estimate absolute endolymph states. In the absence of direct human data, our parameter ranges were therefore anchored to vertebrate vestibular studies, which consistently report utricular [Ca^2+^] around 250–280 μM and pH ~7.6–7.7 (34–37). By contrast, Bächinger et al. (61) reported endolymph calcium concentrations of 0.017–0.133 mmol·L^−1^ (17–133 μM), values largely derived from animal endolymphatic sac and cochlear measurements, and inferred that similar mechanisms may operate in humans (61). These values cannot be directly compared with vestibular (utricle/saccule) endolymph concentrations, underscoring the importance of clearly specifying the anatomical compartment when modeling BPPV.
4.6.1 Sensitivity analyses
We implemented four sensitivity sweeps—(i) activity/ionic-strength assumptions (Davies activity coefficients with Pitzer-bounded ranges; note that ionic strength changes Ω by altering ion activities via γ, i.e., the activity product in Equation 1, rather than Ksp itself); (ii) total inorganic carbon CT (±10%); (iii) the solubility product Ksp (±10%−20%); and (iv) temperature (35–39 °C), updating pK1(T), pK2(T), Ksp(T), and CO_2_ solubility accordingly. Across all four, the Ω = 1 boundary shifted only modestly, and the qualitative dissolution-prone topology was preserved. Boundary displacements were most pronounced under activity/ionic-strength (Supplementary Figure S1) and Ksp variations (Supplementary Figure S3), intermediate for CT (Supplementary Figure S2), and smallest for temperature within the clinical range examined (Supplementary Figure S4). As expected, the Ω = 1 boundary is independent of logistic-mapping parameters (e.g., dispersion σ_C_). These findings support the qualitative robustness of the framework while highlighting where quantitative calibration will matter most for clinical translation.
Future work will: (i) implement activity-aware Ω using bounded Pitzer ranges for ion–ion interactions (62, 63); (ii) estimate CT from measured bicarbonate and pCO2 rather than assuming it fixed; (iii) add carbonate–protein interactions and matrix effects to refine nucleation/dissolution kinetics; and (iv) couple the chemical module to biomechanical simulations of otoconia dynamics to build a multiphysics description of BPPV. Ultimately, prospective, surrogate-based calibration in humans (using serum ionized calcium and blood-gas–derived /pCO_2_, together with clinical outcomes) will be required before any clinical deployment.
Although the framework omits organic-matrix effects, explicit protein–ion interactions, and mechanical forces, its qualitative predictions are expected to hold for human otoconia: because human otoconia are composed of calcium carbonate, the sharp rise in dissolution risk once Ω < 1 is a basic physicochemical property, not a model artifact (see Figure 3). Accordingly, specific numerical values (e.g., Ccrit) and contour shapes should be viewed as illustrative, whereas near-boundary behavior around Ω≈1 is likely to be valid. The model should therefore be regarded as hypothesis-generating, with prospective calibration and incorporation of additional mechanisms required for quantitative validation. Our sensitivity analyses further support this view, showing only modest shifts of the Ω = 1 boundary under physiologically plausible parameter variations while preserving the qualitative dissolution-prone topology.
5 Conclusion
Canalith repositioning maneuvers remain the cornerstone of care for benign paroxysmal positional vertigo (BPPV). Yet the calcium-carbonate composition of human otoconia—unlike the phosphate-based mineral of bone—renders them chemically fragile and sensitive to relatively small ionic and pH perturbations (43). Beyond the established mechanical framework of BPPV— comprising the canalithiasis and cupulolithiasis mechanisms—we introduce an Ω-based biochemical model (pH, [Ca^2+^]) that delineates the otoconia stability–dissolution boundary (Ω≈1) and complements—rather than replaces—this framework.
A minimal, deterministic ion–chemistry framework formalizes this fragility through the carbonate saturation index Ω, which integrates ionized calcium and pH-dependent carbonate speciation. When Ω falls below unity, dissolution predominates, predisposing otoconia to fragmentation and detachment. Given its deliberate parsimony and reliance on synthetic data, the framework is intended as a hypothesis-generating construct rather than a validated clinical tool.
This perspective links systemic influences—such as vitamin D deficiency, estrogen decline, acidosis, and endolymphatic inflammation—to otoconia vulnerability and recurrent BPPV risk. In this translational pathway, serum ionized calcium together with blood-gas–derived /pCO_2_ are positioned only as non-invasive surrogates for relative-risk inference (a blood-based Ω proxy), rather than one-to-one estimators of absolute endolymph chemistry. Prospective, surrogate-based validation and calibration in humans—along with robustness testing to variations in CT, ionic strength/activity coefficients, Ksp, and temperature within physiological bounds—are required before any clinical deployment.
Finally, coupling this chemical module to biomechanical simulations of otoconia dynamics offers a route toward a multiphysics description of BPPV pathophysiology. A prospectively validated, activity-aware Ω (with bounded Pitzer ranges) integrated into such a framework may ultimately provide a quantitative bridge between vestibular biochemistry and clinical management; at present, the model is best viewed as a clear, testable basis for future studies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1von Brevern M Radtke A Lezius F Feldmann M Ziese T Lempert T. Epidemiology of benign paroxysmal positional vertigo: a population-based study. J Neurol Neurosurg Psychiatry. (2007) 78:710–5. 10.1136/jnnp.2006.10042017135456 PMC 2117684 · doi ↗ · pubmed ↗
- 2Epley JM. New dimensions of benign paroxysmal positional vertigo. Otolaryngol Head Neck Surg. (1980) 88:599–605. 10.1177/0194599880088005147443266 · doi ↗ · pubmed ↗
- 3Schuknecht HF. Cupulolithiasis. Arch Otolaryngol. (1969) 90:765–78. 10.1001/archotol.1969.007700307670205353084 · doi ↗ · pubmed ↗
- 4Ross MD Peacor D Johnsson LG Allard LF. Observations on normal and degenerating human otoconia. Ann Otol Rhinol Laryngol. (1976) 85:310–26. 10.1177/000348947608500302937958 · doi ↗ · pubmed ↗
- 5Walther LE. Otokonien: aktuelle Aspekte aus der Forschung [Otoconia: current aspects of research] [German]. HNO. (2016) 64:767–76. 10.1007/s 00106-016-0234-727590488 · doi ↗ · pubmed ↗
- 6Tohse H Mugiya Y. Effects of acidity and a metabolic inhibitor on incorporation of calcium and inorganic carbon into endolymph and otoliths in salmon Oncorhynchus masou. Fish Sci. (2004) 70:595–600. 10.1111/j.1444-2906.2004.00846.x · doi ↗
- 7Payan P De Pontual H Boeuf G Mayer-Gostan N. Endolymph chemistry and otolith growth in fish. C R Palevol. (2004) 3:535–47. 10.1016/j.crpv.2004.07.013 · doi ↗
- 8Byun H Chung JH Lee SH Park CW Kim EM Kim I. Increased risk of benign paroxysmal positional vertigo in osteoporosis: a nationwide population-based cohort study. Sci Rep. (2019) 9:3469. 10.1038/s 41598-019-39830-x 30837524 PMC 6401187 · doi ↗ · pubmed ↗