Arabidopsis thaliana Cyclic Nucleotide‐Gated Channel 19 is involved in root extracellular ATP and Pep1 signalling
Youzheng Ning, Bryony C. I. C. Jacobs, Clementine Langlet, Limin Wang, Zhizhong Song, Adeeba M. Dark, Elsa Matthus, Sebastian Eves‐van den Akker, Taufiq Rahman, Julia M. Davies

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
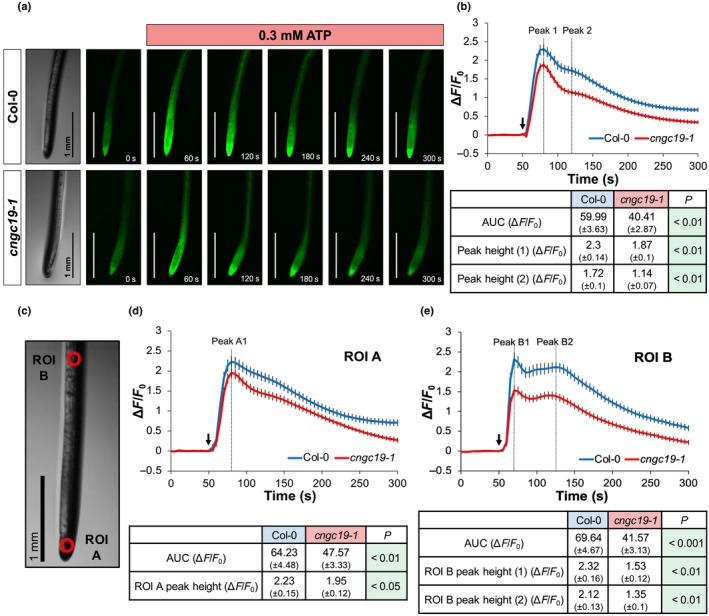 Fig. 1
Fig. 1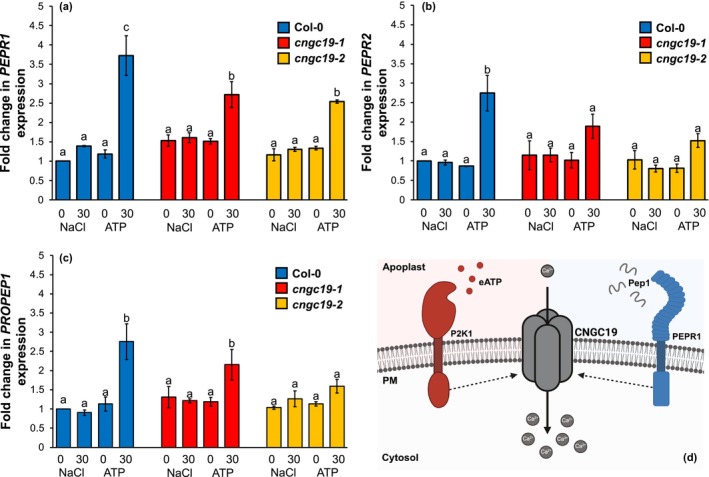 Fig. 2
Fig. 2- —Biotechnology and Biological Sciences Research Council10.13039/501100000268
- —University of Cambridge10.13039/501100000735
- —Henry Lester Trust10.13039/501100017646
- —Ludong University10.13039/501100008279
- —China Scholarship Council10.13039/501100004543
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhotosynthetic Processes and Mechanisms · ATP Synthase and ATPases Research · Mechanisms of cancer metastasis
Disclaimer
The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.
Introduction
Extracellular ATP (eATP) can function as a damage‐associated molecular pattern (DAMP) that signals wounding incurred by microbial ingress and herbivore attacks (Tanaka & Heil, 2021; Yuan et al., 2025). Arabidopsis thaliana root and leaf wounding trigger rapid accumulation of eATP (Dark et al., 2011; Myers et al., 2022). eATP is then sensed by the plasma membrane (PM) co‐receptors Does Not Respond to Nucleotide1/P2‐type purinoceptor Kinase 1 (DORN1/P2K1) and P2K2 (Choi et al., 2014; Pham et al., 2020). The perception of eATP leads to downstream signalling cascades involving elevation of cytosolic free calcium ([Ca^2+^]cyt), reactive oxygen species and nitric oxide plus MAP (Mitogen‐Activated Protein) kinase activation (Demidchik et al., 2003; Foresi et al., 2007; Chen et al., 2017; Kim et al., 2023). Analyses of eATP‐induced transcriptomes point to DORN1/P2K1‐dependent regulation of multiple defence‐related hormone signalling pathways involving jasmonic acid (JA), ethylene and salicylic acid (Tripathi et al., 2017; Jewell et al., 2019). Accordingly, loss‐of‐function DORN1/P2K1 and P2K2 mutants show increased susceptibility to a variety of plant pathogens, including fungi (Rhizoctonia solani and Sclerotinia sclerotiorum), oomycetes (Plasmodiophora brassicae, Phytophthora infestans and Phytophthora capsici) and bacteria (Pseudomonas syringae) plus insect herbivores (Pieris rapae, Salix exigua and Spodoptera litura), whilst receptor overexpression lines reduce plant susceptibility (Bouwmeester et al., 2014; Balagué et al., 2017; Chen et al., 2017; Jewell et al., 2022; Kumar et al., 2020; Kundu et al., 2025; Yuan et al., 2025).
[Ca^2+^]cyt serves as an important second messenger in plant stress and immunity signalling (Dong et al., 2022; Köster et al., 2022). eATP induces a specific biphasic [Ca^2+^]cyt ‘signature’ in roots through DORN1/P2K1, with the possibility of a relatively minor involvement of P2K2 or as yet unknown receptors (Demidchik et al., 2003; Zhu et al., 2018, 2020; Matthus et al., 2019a,b, 2022; Smith et al., 2021). How eATP receptors initiate this [Ca^2+^]cyt increase in roots is not fully understood (Sun et al., 2021), but the PM Cyclic Nucleotide‐Gated Channel2 (CNGC2) is involved, with the possibility of CNGC4 and CNGC6 also (Duong et al., 2022; Wang et al., 2022). The Arabidopsis CNGC family consists of 20 channel‐subunit members, with important functions in signalling (Jarratt‐Barnham et al., 2021). CNGC2 has recently been shown to be phosphorylated by DORN1/P2K1 in leaves (Sun et al., 2025) and was reported to mediate the Arabidopsis root's [Ca^2+^]cyt response to eATP (Wang et al., 2022). However, the significant residual [Ca^2+^]cyt increase in the loss‐of‐function cngc2 mutant indicates that other channels also support the eATP‐induced [Ca^2+^]cyt signal in roots (Wang et al., 2022). Studies have revealed Cyclic Nucleotide‐Gated Channel 19 (CNGC19) involvement as a PM Ca^2+^‐permeable channel in generating the [Ca^2+^]cyt signal and defence signalling pathways induced by herbivore attack on leaves and fungal colonisation of roots (Meena et al., 2019; Jogawat et al., 2020). Both of these challenges can trigger eATP accumulation through wounding (Nizam et al., 2019; Myers et al., 2022). Moreover, CNGC19 and DORN1/P2K1 are co‐expressed whilst eATP upregulates DORN1/P2K1‐dependent CNGC19 expression specifically in roots (Tripathi et al., 2017; Jewell et al., 2019; Sowders & Tanaka, 2023). Such findings suggest that CNGC19 may be part of the eATP response pathway. Indeed, a recent study by Kundu et al. (2025) has shown that CNGC19 is part of the whole seedling [Ca^2+^]cyt response to eATP (measured using cytosolic aequorin in the cngc19‐2 loss‐of‐function mutant) and that CNGC19 is a possible interacting partner of DORN1/P2K1 (Kundu et al., 2025). This begs the question of whether CNGC19 operates in the root's response to eATP.
In this study, CNGC19 was found to make a significant contribution to the Arabidopsis root's eATP‐induced [Ca^2+^]cyt increase (determined using the intensiometric GCaMP3 [Ca^2+^]cyt reporter (Vincent et al., 2017) in the cngc19‐1 T‐DNA insertion loss‐of‐function mutant). The root transcriptional response to eATP revealed amplification of CNGC19 expression through DORN1/P2K1, but only partially through CNGC2. Transcription of specific eATP‐induced defence‐related genes was found to require CNGC19, indicative of discrete signalling pathways. These genes included those for the DAMP PM Plant Endogenous Peptide Receptors PEPR1 and PEPR2. As CNGC19 is also needed for Pep1‐induced [Ca^2+^]cyt increase (determined with cngc19‐2 mutant seedlings by Meena et al., 2019 and also here with cngc19‐1 roots), together the results point to CNGC19 as a common component of these two DAMP pathways.
Results and Discussion
CNGC19 contributes to eATP‐induced [Ca2+]cyt increase in roots
Columbia‐0 (Col‐0) and cngc19‐1 (both expressing cytosolic 35S::GCaMP3; Supporting Information Fig. S1a) were used to test for CNGC19's involvement in the root's biphasic [Ca^2+^]cyt response to 0.3 mM eATP (the same concentration used in previous work on CNGC2; Wang et al., 2022) (Methods S1–S3; Fig. 1a; Videos S1, S2). Analysis of the eATP‐induced [Ca^2+^]cyt increase (Fig. 1b) showed that the cngc19‐1 mutant was significantly impaired in the magnitude of the biphasic maximal responses (‘peaks’) and total [Ca^2+^]cyt mobilised (estimated as area under the curve (AUC)). Previous research using GCaMP3 and aequorin has shown that the root apex generates the first increment (Peak 1) of the [Ca^2+^]cyt increase and the semi‐autonomous Peak 2 then occurs sub‐apically (Matthus et al., 2019a,b, 2022). Here, a region of interest (ROI) was placed at the apex (ROI A) and sub‐apically (ROI B; Fig. 1c). Rapid and distinctive fluorescence increases occurred at both ROI A and ROI B in response to 0.3 mM eATP, with cngc19‐1 showing significantly lower maximum responses and AUC in both ROIs compared with Col‐0 (Fig. 1d,e). There were no significant differences between genotypes in response to 0.6 mM NaCl control solution (Fig. S1b–f; Videos S3, S4) whilst the eATP‐induced [Ca^2+^]cyt response was significantly greater than the control in both genotypes (Fig. S2). With normal expression of the DORN1/P2K1 and P2K2 eATP receptors evident in cngc19‐1 roots (Fig. S3a,b, measured by quantitative reverse transcription polymerase chain reaction (qRT‐PCR); Methods S3, S4; Table S1), the lesions in the mutant's [Ca^2+^]cyt response to eATP are unlikely to be the result of impaired ability to sense eATP and instead point to CNGC19's acting as a PM Ca^2+^ channel in eATP signalling.
Root extracellular ATP (eATP)‐induced cytosolic free calcium ([Ca2+]cyt) response is significantly reduced in Arabidopsis thaliana cyclic nucleotide‐gated channel19‐1 (cngc19‐1). eATP‐induced fluorescence (F) was measured for 250 s in Columbia‐0 (Col‐0) and cngc19‐1 roots (expressing cytosolic GCaMP3) following a 0.3 mM eATP treatment at 50 s. (a) Intensiometric images (including a brightfield) taken from a single representative Col‐0 or cngc19‐1 root across a specified time series. The 0.3 mM eATP treatment is indicated by a box. Bars, 1 mm. (b) Mean ± SE normalised GCaMP3 fluorescence (ΔF/F 0) in the full root tip (c. 2.5 mm) in response to 0.3 mM eATP in Col‐0 and cngc19‐1. The black arrow indicates application of a 1 μl 0.3 mM eATP treatment (50 s). Two ΔF/F 0 peaks (Peak 1, 80 s; Peak 2, 120 s) are shown by dashed lines. eATP‐dependent area under the curve (AUC) and peak height analyses are also shown (±SE), with significant differences (P) between Col‐0 and cngc19‐1 determined by Student's t‐test. (c) Brightfield image of a root tip annotated with the two regions of interest (ROI) investigated, ‘A’ and ‘B’. ROI A represents a 150 μm2 region found within the first 1 mm of the root apex and ROI B represents a 150 μm2 region c. 2.5 mm away from the root apex. Mean ± SE normalised GCaMP3 fluorescence (ΔF/F 0) measured in response to 0.3 mM eATP in (d) ROI A and (e) ROI B in Col‐0 and cngc19‐1. eATP‐induced AUC and ΔF/F 0 peak height analyses are also included; (d) ROI A (Peak A1, 80 s) and (e) ROI B (Peak B1, 70 s; Peak B2, 125 s). The significance value (P) for differences between Col‐0 and cngc19‐1 were determined by a Student's t‐test. Data were obtained across six biological replicates with n = 39 (Col‐0) and n = 35 (cngc19‐1).
CNGC19 expression is DORN1/P2K1‐dependent but does not affect receptor expression
eATP's inability to increase eATP receptor expression agrees with results on roots from a DORN1/P2K1 GUS reporter line (Sowders & Tanaka, 2023). By contrast, eATP (0.3 mM) significantly increased CNGC19 expression in Col‐0 roots in a DORN1/P2K1‐dependent manner; induction was significantly lower in the dorn1‐3 loss‐of‐function mutant (Fig. S3c,d). Significant induction in Col‐0 was only evident after 30 min, tallying with a previous finding of a slow response to eATP using CNGC19 promoter‐driven GUS expression in roots (Sowders & Tanaka, 2023). Moreover, eATP‐induced CNGC19 expression was dependent on CNGC2 (which lies downstream of DORN1/P2K1 in root [Ca^2+^] signalling; Wang et al., 2022). Significant CNGC19 induction in Col‐0 and the complemented cngc2‐3 mutant was evident at 30 min but was significantly impaired in the cngc2‐3 loss‐of‐function mutant (Fig. S3e). However, a significant level of CNGC19 upregulation remained in the cngc2‐3 mutant, suggesting the presence of another pathway downstream of DORN1/P2K1 (Fig. S3e). In contrast to CNGC19 (but in common with the eATP receptors), CNGC2 expression was not significantly increased by eATP and was unaffected by loss of CNGC19 function (Fig. S3f). Overall, these results highlight positive feedback in roots between eATP and CNGC19 expression via the DORN1/P2K1 signalling pathway that is not wholly reliant on CNGC2. They also extend the observation that DORN1/P2K1 expression in roots is not amplified by eATP (Sowders & Tanaka, 2023) to include P2K2 and CNGC2. Thus, there appear to be modules of eATP signalling components that at the transcriptional level vary in their responsiveness to eATP.
CNGC19 contributes to eATP‐induced transcriptional responses and may link to immunopeptide DAMP signalling
eATP regulates transcription of immunity‐related genes through DORN1/P2K1 (Tripathi et al., 2017; Jewell et al., 2019; Yuan et al., 2025). Whether this is mediated by CNGC19 has yet to be tested directly in any tissue. Upregulation of Jasmonic Acid ZIM‐domain 5 (JAZ5) by eATP or in R. solani‐infected roots is DORN1/P2K1‐dependent (Tripathi et al., 2017; Kumar et al., 2020). Here, JAZ5 in roots was significantly upregulated by eATP in Col‐0 but not cngc19‐1 (Fig. S4a), suggesting that CNGC19 is needed to regulate JA‐mediated responses (Tripathi et al., 2017; Jewell et al., 2019). Defence‐related genes WRKY DNA‐Binding Protein 40 and Mitogen‐Activated Protein Kinase 3, which are both dependent on DORN1/P2K1 and CNGC2 for eATP upregulation in roots (Wang et al., 2022), also required CNGC19 on longer (30 min) eATP exposure (Fig. S4b,c). By contrast, the response of Calcium‐dependent Protein Kinase 28 (also DORN1/P2K1‐ and CNGC2‐dependent for transcription) encoding a regulatory DORN1/P2K1‐interacting protein (Wang et al., 2022; Sowders et al., 2024) showed no significant difference between genotypes (Fig. S4d). This suggests a CNGC19‐specific defence‐related transcriptome induced by eATP.
Damaged root cells not only release ATP but also immunopeptides, such as Pep1 (Hander et al., 2019). Damage‐induced [Ca^2+^]cyt increase causes activation of the MetaCaspase4 zymogen; this cysteine protease cleaves the inactive precursor PROPEP1 to the active form Pep1, which then exits the cell to activate defence responses by binding to PEPR1 or PEPR2 (Hander et al., 2019). eATP upregulates transcription of the PEPR1/PEPR2/PROPEP1 DAMP signalling module through DORN1/P2K1 (Tripathi et al., 2017; Jewell et al., 2019). Here, eATP upregulation of PEPR1/PEPR2 was impaired in both cngc19‐1 and cngc19‐2 roots after 30 min eATP treatment (Fig. 2a,b), although upregulation of PROPEP1 was only significantly impaired in cngc19‐2 (Fig. 2c). That cngc19‐1 and cngc19‐2 do not always appear to be allelic was noted previously by Meena et al. (2019). Nevertheless, the PEPR data are indicative of a pathway for priming and augmenting longer term DAMP signalling. Indeed, Pep1 upregulates CNGC19 expression in root meristems (Dhar et al., 2025).
Cyclic Nucleotide‐Gated Channel 19 (CNGC19) is required for extracellular ATP (eATP)‐induced PEPR1 and PEPR2 expression in Arabidopsis thaliana roots. Whole roots of Col‐0, cyclic nucleotide gated channel19‐1 (cngc19‐1) and cngc19‐2 were treated with 0.3 mM ATP or 0.6 mM NaCl for 0 or 30 min before RNA extraction was carried out, followed by quantitative reverse transcription polymerase chain reaction (qRT‐PCR). UBQ10 (Poly Ubiquitin10) and Tubulin beta chain 4 (TUB4) were used as housekeeping genes for normalisation. (a) Plant Endogenous Peptide Receptor1 (PEPR1), (b) PEPR2 and (c) Pro Peptide1 (PROPEP1) expression in Col‐0, cngc19‐1 and cngc19‐2. Data are means (±SE) of three biological replicates, with each biological replicate, including three technical replicates. Different letters indicate significant differences as determined using two‐way ANOVA and a post hoc Tukey Honestly Significant Difference (HSD) test. (0, 0 min; 30, 30 min; ATP, 0.3 mM ATP treatment; NaCl, 0.6 mM NaCl treatment). (d) Schematic of the suggested involvement of CNGC19 (shown as a homotetramer for simplicity but heterotetramers are also feasible) in the root's [Ca2+]cyt (cytosolic free Ca2+) response to eATP and Peptide 1 (Pep1). eATP and Pep1 are recognised by the plasma membrane‐bound receptors, Does Not Respond to Nucleotide1/P2‐type purinoceptor Kinase 1 (DORN1/P2K1) and PEPR1, respectively. Pep1 binding to PEPR2 has been omitted for simplicity. The binding of these putative ligands to their receptors would promote the opening of CNGC19 to mediate the influx of Ca2+ to the cytosol. Dashed lines indicate proposed interactions. For eATP, this could include physical interaction (Kundu et al., 2025), phosphorylation by the receptor's cytosolic kinase domain or possible production of cyclic guanosine monophosphate (GMP; Sun et al., 2021). For the peptide pathway, there is no physical interaction between CNGC19 and PEPR1 (or PEPR2; Meena et al., 2019) whilst PEPR1 (and PEPR2) may be capable of cyclic GMP synthesis or phosphorylation by their kinase domains (Qi et al., 2010; Ma et al., 2012; Liu et al., 2013). PM, plasma membrane. Figure created in BioRender (https://BioRender.com/44pdker).
That cngc19‐1 is impaired in eATP‐induced root [Ca^2+^]cyt elevation whilst cngc19‐2 seedlings are impaired in both eATP‐ and Pep1‐induced [Ca^2+^]cyt increase (Meena et al., 2019; Kundu et al., 2025) points to CNGC19 as a common component of these two DAMP pathways. Certainly, CNGC19 co‐expresses with DORN1/P2K1, PEPR1, PEPR2 and PROPEP1 (Obayashi et al., 2022). Here, Pep1 induced a [Ca^2+^]cyt increase in roots (measured with GCaMP3) that was significantly impaired in cngc19‐1 (Videos S5–S8; Fig. S5). Similar basal PEPR1 and PEPR2 expression in Col‐0 and cngc19‐1 roots (Fig. 2a,b) suggests that this impairment is due to loss of channel function. Overall, results point to CNGC19's operating in both eATP and Pep1 pathways in roots. CNGC2 has also been found to function in both eATP and Pep signalling (Qi et al., 2010; Wang et al., 2022), and it would be interesting to test a cngc2/cngc19 double mutant. However, such a mutant would be unlikely to yield a complete lack of response to eATP as CNGC4 and CNGC6 are also implicated in that pathway (Duong et al., 2022; Wang et al., 2022). Whether CNGC19 acts in both eATP and Pep signalling at the level of an individual cell now needs to be determined, as does the scope for utilising this protein in improving plant responses to damage (Fig. 2d).
Competing interests
None declared.
Author contributions
YN, BCICJ, LW, ZS, SEA, TR and JMD planned and designed the research. BCICJ, YN, CL, LW, ZS, AMD, EM and JMD performed the experiments and analysed the data. All authors contributed to writing. BCICJ and JMD revised the manuscript. YN and BCICJ contributed equally to this work.
Supporting information
Fig. S1 Col‐0 and cngc19‐1 root [Ca^2+^]cyt response to 0.6 mM NaCl is statistically similar. Fig. S2 The eATP‐induced [Ca^2+^]cyt response is significantly larger than the response to control solution in both Col‐0 and cngc19‐1 roots. Fig. S3 eATP receptor expression is independent of CNGC19, but eATP‐induced CNGC19 expression requires DORN1/P2K1 and CNGC2. Fig. S4 CNGC19 is required for eATP‐induced JAZ5, WRKY40 and MPK3 expression in roots. Fig. S5 The root Pep1‐induced [Ca^2+^]cyt response is significantly reduced in cngc19‐1. Methods S1 Plant materials and growth conditions. Methods S2 GCaMP3 fluorescence microscopy. Methods S3 Statistical analyses. Methods S4 qRT‐PCR gene expression studies. Table S1 The primer sequences used in qRT‐PCR.
Video S1 The eATP‐induced [Ca^2+^]cyt response in a representative Col‐0 root.
Video S2 The eATP‐induced [Ca^2+^]cyt response in a representative cngc19‐1 root.
Video S3 The NaCl‐induced [Ca^2+^]cyt response in a representative Col‐0 root.
Video S4 The NaCl‐induced [Ca^2+^]cyt response in a representative cngc19‐1 root.
Video S5 The Pep1‐induced [Ca^2+^]cyt response in a representative Col‐0 root.
Video S6 The Pep1‐induced [Ca^2+^]cyt response in a representative cngc19‐1 root.
Video S7 The control‐induced [Ca^2+^]cyt response in a representative Col‐0 root.
Video S8 The control‐induced [Ca^2+^]cyt response in a representative cngc19‐1 root.Please note: Wiley is not responsible for the content or functionality of any Supporting Information supplied by the authors. Any queries (other than missing material) should be directed to the New Phytologist Central Office.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Balagué C , Gouget A , Bouchez O , Souriac C , Haget N , Boutet‐Mercey S , Govers F , Roby D , Canut H . 2017. The Arabidopsis thaliana lectin receptor kinase Lec RK‐I.9 is required for full resistance to Pseudomonas syringae and affects jasmonate signalling. Molecular Plant Pathology 18: 937–948.27399963 10.1111/mpp.12457 PMC 6638305 · doi ↗ · pubmed ↗
- 2Bouwmeester K , Han M , Blanco‐Portales R , Song W , Weide R , Guo LY , van der Vossen EA , Govers F . 2014. The Arabidopsis lectin receptor kinase Lec RK‐I. 9 enhances resistance to Phytophthora infestans in Solanaceous plants. Plant Biotechnology Journal 12: 10–16.23980842 10.1111/pbi.12111 · doi ↗ · pubmed ↗
- 3Chen D , Cao Y , Li H , Kim D , Ahsan N , Thelen J , Stacey G . 2017. Extracellular ATP elicits DORN 1‐mediated RBOHD phosphorylation to regulate stomatal aperture. Nature Communications 8: 2265.10.1038/s 41467-017-02340-3PMC 574162129273780 · doi ↗ · pubmed ↗
- 4Choi J , Tanaka K , Cao Y , Qi Y , Qiu J , Liang Y , Lee SY , Stacey G . 2014. Identification of a plant receptor for extracellular ATP. Science 343: 290–294.24436418 10.1126/science.343.6168.290 · doi ↗ · pubmed ↗
- 5Dark A , Demidchik V , Richards SL , Shabala S , Davies JM . 2011. Release of extracellular purines from plant roots and effect on ion fluxes. Plant Signaling & Behavior 6: 1855–1857.22057319 10.4161/psb.6.11.17014 PMC 3329368 · doi ↗ · pubmed ↗
- 6Demidchik V , Nichols C , Oliynyk M , Dark A , Glover BJ , Davies JM . 2003. Is ATP a signaling agent in plants? Plant Physiology 133: 456–461.14555773 10.1104/pp.103.024091 PMC 1540337 · doi ↗ · pubmed ↗
- 7Dhar S , Kim SY , Shin HJ , Park J , Lee JY . 2025. The molecular framework balancing growth and defense in response to plant elicitor peptide‐induced signals in Arabidopsis . Plant Cell 37: koae 327.10.1093/plcell/koae 327PMC 1168407939700410 · doi ↗ · pubmed ↗
- 8Dong Q , Wallrad L , Almutairi BO , Kudla J . 2022. Ca 2+ signaling in plant responses to abiotic stresses. Journal of Integrative Plant Biology 64: 287–300.35048537 10.1111/jipb.13228 · doi ↗ · pubmed ↗