Effects of Ploidy on Relationship Between Outbreeding Response and Fitness in a Plant Selfing Species
Carlos Olmedo‐Castellanos, Ana García‐Muñoz, Camilo Ferrón, Mohammed Bakkali, A. Jesús Muñoz‐Pajares, Mohamed Abdelaziz

TL;DR
The study shows how ploidy levels affect outbreeding response and fitness in a selfing plant species, suggesting it could influence mating system evolution.
Contribution
The paper demonstrates that outbreeding response is ploidy-dependent and functionally connected to fitness in a selfing plant species.
Findings
Tetraploids showed the strongest and most consistent positive outbreeding responses, especially in gamete production.
Increasing heterozygosity was linked to reduced phenotypic integration.
Outbreeding response is functionally tied to fitness and may act as a selective force promoting outcrossing.
Abstract
Outbreeding response, the phenotypic differences observed between selfed parental lines and their outcrossed offspring, can influence the evolution of selfing strategies. However, such effects remain poorly understood in noncrop species. We explored the phenotypic outbreeding response variation across ploidy levels in Erysimum incanum, a predominantly selfing plant complex with diploid, tetraploid, and hexaploid populations distributed across the Iberian Peninsula and Morocco. We performed controlled within‐population crosses to generate offspring with varying heterozygosity levels across ploidy types. We quantified individual, flower, and reproductive traits, and we estimated fitness components, and assessed trait modularity and phenotypic integration to see how heterozygosity affects trait coordination. Tetraploids showed the strongest and most consistently positive outbreeding…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
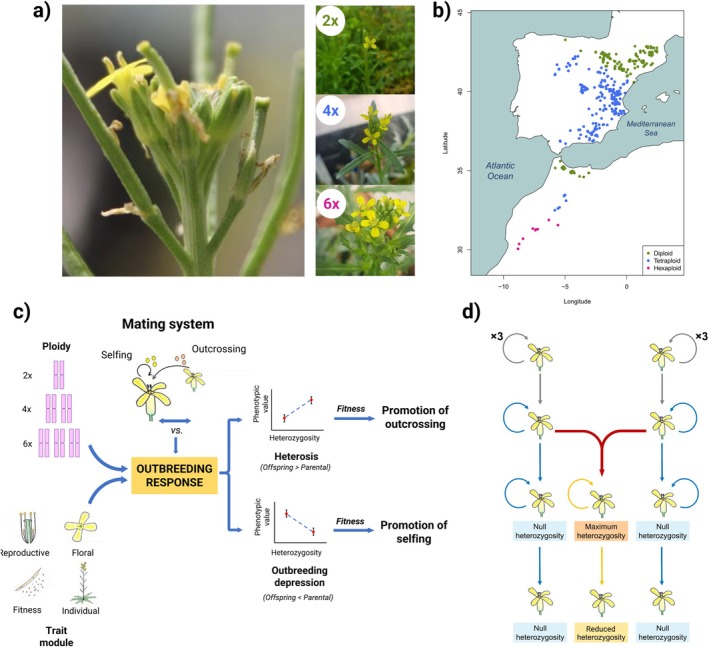 FIGURE 1
FIGURE 1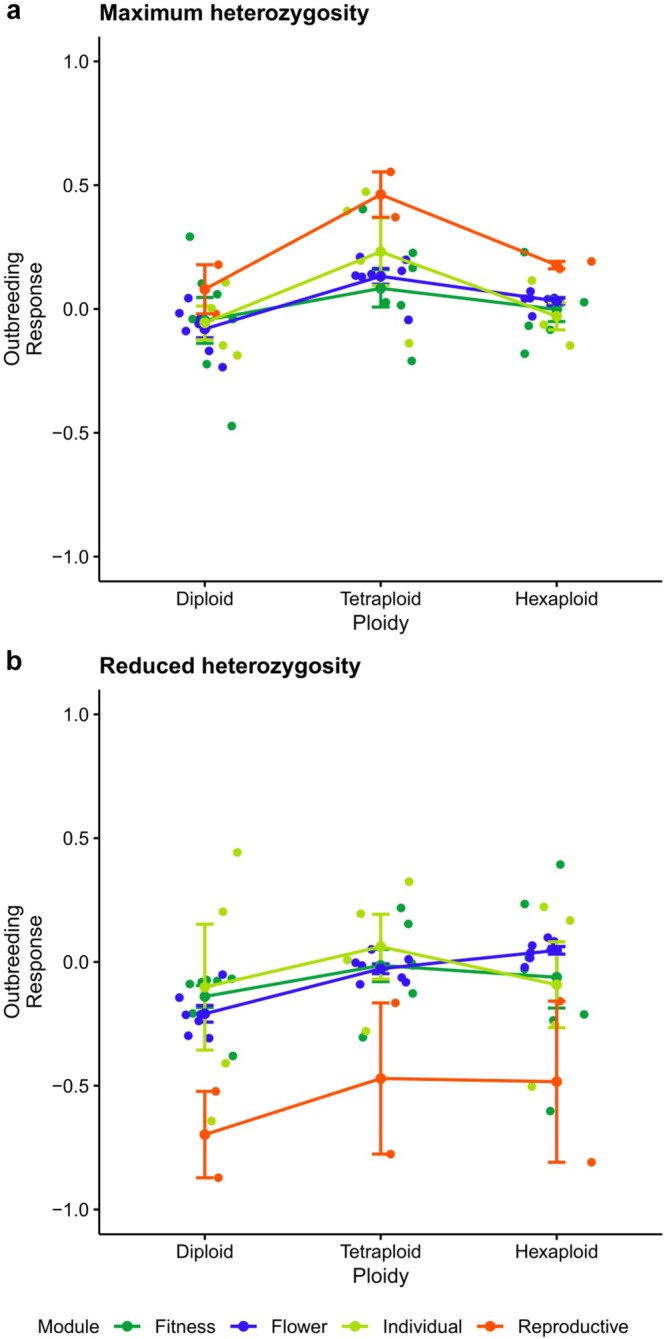 FIGURE 2
FIGURE 2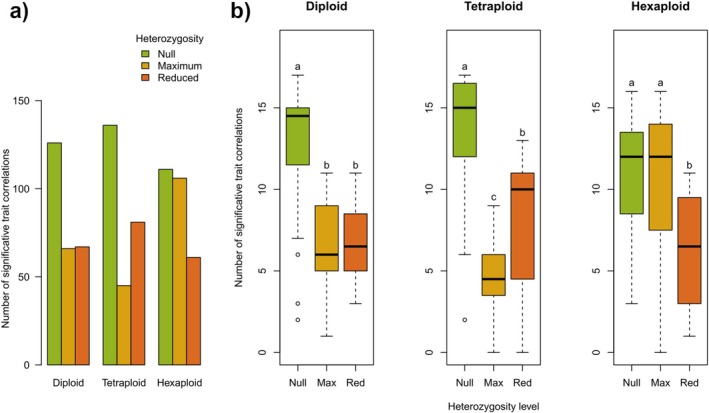 FIGURE 3
FIGURE 3| Ploidy | Population | Location | Coordinates |
|---|---|---|---|
| Diploid |
| Rif (Morocco) |
35° 11′ 8″ N 5° 13′ 19″ W |
| Diploid |
| Pyrenees (Spain) |
41° 58′ 20″ N 0° 46′ 43″ W |
| Tetraploid |
| Middle Atlas (Morocco) |
33° 27′ 06″ N 4° 57′ 38″ W |
| Tetraploid |
| Guadix (Spain) |
37° 20′ 18″ N 3° 02′ 38″ W |
| Hexaploid |
| Anti‐Atlas (Morocco) |
30° 50′ 04″ N 8° 23′ 38″ W |
| Hexaploid |
| Anti‐Atlas (Morocco) |
30° 14′ 16″ N 8° 10′ 37″ W |
| Module | Diploid | 95% CI | Tetraploid | 95% CI | Hexaploid | 95% CI |
|---|---|---|---|---|---|---|
|
| ||||||
| Ovules/Plant | −0.041 | (−0.285, 0.239) | 0.403 | (0.163, 0.562) | 0.230 | (−0.042, 0.425) |
| Pollen/Plant | 0.156 | (−1.325, 0.566) | 0.687 | (0.528, 0.786) | 0.064 | (−0.290, 0.318) |
| Fruitset | 0.103 | (−0.014, 0.189) | 0.227 | (0.181, 0.269) | 0.039 | (−0.015, 0.088) |
| Seedset | 0.059 | (−0.015, 0.112) | 0.027 | (−0.061, −0.068) | −0.068 | (−0.174, 0.032) |
| Survival | 0.292 | (0.227, 0.347) | 0.015 | (−0.119, 0.127) | −0.084 | (−0.169, 0.009) |
| Germination | −0.473 | (−0.559, −0.380) | −0.210 | (−0.298, −0.120) | −0.181 | (−0.322, 0.002) |
| Number of leaves | −0.223 | (−0.326, −0.114) | −0.045 | (−0.132, 0.039) | 0.024 | (−0.172, 0.199) |
|
| ||||||
| Ovules/Flower | −0.019 | (−0.173, 0.147) | 0.371 | (0.291, 0.437) | 0.162 | (0.076, 0.238) |
| Pollen/Flower | 0.179 | (−0.227, 0.421) | 0.554 | (0.445, 0.638) | 0.192 | (0.042, 0.316) |
|
| ||||||
| Plant height | 0.003 | (−0.184, 0.151) | 0.473 | (0.410, 0.530) | 0.115 | (−0.013, 0.224) |
| Number of stalks | −0.187 | (−0.438, 0.121) | −0.138 | (−0.326, 0.073) | −0.148 | (−0.328, 0.078) |
| Stalk diameter | 0.107 | (−0.106, 0.271) | 0.396 | (0.304, 0.474) | −0.021 | (−0.148, 0.121) |
| Number of flowers | −0.147 | (−0.363, 0.099) | 0.196 | (−0.006, 0.348) | −0.063 | (−0.259, 0.175) |
|
| ||||||
| Petal length | −0.089 | (−0.163, −0.003) | 0.135 | (0.089, 0.178) | 0.015 | (−0.044, 0.068) |
| Flower diameter | −0.060 | (−0.122, 0.006) | 0.141 | (0.098, 0.182) | −0.030 | (−0.072, 0.014) |
| Corolla tube length | −0.017 | (−0.083, 0.062) | 0.130 | (0.068, 0.192) | 0.045 | (−0.028, 0.112) |
| Long filament length | 0.044 | (−0.018, 0.101) | 0.199 | (0.172, 0.225) | 0.042 | (0.008, 0.074) |
| Short filament length | −0.035 | (−0.115, 0.050) | 0.154 | (0.115, 0.190) | 0.043 | (−0.008, 0.090) |
| Style length | −0.169 | (−0.424, 0.081) | 0.210 | (0.175, 0.244) | 0.071 | (0.026, 0.115) |
| Herkogamy | −0.235 | (−0.480, 0.012) | −0.044 | (−0.087,0.000) | 0.040 | (−0.016, 0.091) |
| Module | Diploid | 95% CI | Tetraploid | 95% CI | Hexaploid | 95% CI |
|---|---|---|---|---|---|---|
|
| ||||||
| Ovules/Plant | −0.380 | (−0.529, −0.198) | −0.165 | (−0.304, 0.007) | −0.029 | (−0.398, 0.457) |
| Pollen/Plant | −0.208 | (−0.259, −0.156) | −0.127 | (−0.166, −0.086) | −0.212 | (−0.29, −0.135) |
| Fruitset | −0.082 | (−0.157, −0.005) | −0.014 | (−0.073, 0.047) | −0.236 | (−0.352, −0.123) |
| Seedset | −0.077 | (−0.139, −0.019) | −0.020 | (−0.054, 0.014) | 0.017 | (−0.038, 0.070) |
| Germination | −0.068 | (−0.174, 0.048) | −0.006 | (−0.076, 0.068) | 0.234 | (0.064, 0.379) |
| Survival | −0.089 | (−0.186, 0.013) | 0.218 | (0.157, 0.274) | −0.602 | (−0.649, −0.552) |
| Number of leaves | −0.073 | (−0.156, 0.019) | 0.153 | (0.121, 0.185) | 0.393 | (0.257, 0.510) |
|
| ||||||
| Ovules/Flower | −0.523 | (−0.578, −0.462) | −0.165 | (−0.224, −0.102) | −0.158 | (−0.234, −0.075) |
| Pollen/Flower | −0.872 | (−0.921, −0.808) | −0.776 | (−0.826, −0.717) | −0.809 | (−0.863, −0.745) |
|
| ||||||
| Plant height | −0.642 | (−0.682, −0.597) | −0.280 | (−0.367, −0.173) | −0.504 | (−0.563, −0.436) |
| Number of stalks | 0.442 | (0.320, 0.540) | 0.324 | (0.240, 0.398) | 0.167 | (−0.122, 0.367) |
| Stalk diameter | −0.410 | (−0.481, −0.327) | 0.008 | (−0.462, 0.387) | −0.255 | (−0.395, −0.096) |
| Number of flowers | 0.203 | (0.024, 0.342) | 0.194 | (0.095, 0.284) | 0.222 | (−0.069, 0.418) |
|
| ||||||
| Petal length | −0.214 | (−0.260, −0.165) | −0.003 | (−0.040, 0.035) | 0.049 | (−0.012, 0.104) |
| Flower diameter | −0.239 | (−0.283, −0.192) | 0.050 | (0.019, 0.081) | 0.066 | (0.023, 0.106) |
| Corolla tube length | −0.144 | (−0.186, −0.101) | −0.015 | (−0.046, 0.018) | 0.083 | (0.036, 0.127) |
| Long filament length | −0.298 | (−0.332, −0.262) | −0.082 | (−0.109, −0.055) | −0.021 | (−0.061, 0.019) |
| Short filament length | −0.211 | (−0.258, −0.162) | −0.063 | (−0.094, −0.031) | 0.016 | (−0.035, 0.063) |
| Style length | −0.308 | (−0.516, −0.107) | −0.090 | (−0.122, −0.058) | 0.037 | (−0.033, 0.103) |
| Herkogamy | −0.051 | (−0.356, 0.254) | 0.011 | (−0.015, 0.036) | 0.098 | (0.035, 0.158) |
| Ploidy | Diploid | Tetraploid | Hexaploid | |||
|---|---|---|---|---|---|---|
| Heterozygosity | Maximum | Reduced | Maximum | Reduced | Maximum | Reduced |
| Ovules/Plant | 0.156**** | 0.119**** | 0.262**** | 0.086**** | 0.203**** | 0.203**** |
| Pollen/Plant | 0.141 | 0.054 | 0.304** | 0.033 | 0.342**** | 0.339* |
| Fruit set | 0.087* | 0.076** | 0.192 | 0.071**** | 0.095 | 0.128* |
| Seed set | 0.115 | 0.082* | 0.15 | 0.067**** | 0.128** | 0.181 |
| Survival | 0.065 | −0.046** | −0.041 | 0.03** | −0.002 | −0.121 |
| Germination | 0.034 | −0.047*** | 0.028 | 0.016 | 0.056* | −0.047* |
| Number of leaves | 0.062* | −0.029 | 0.016 | 0.015 | 0.046 | −0.039* |
| Ovules/Flower | 0.064 | 0.172**** | 0.193 | 0.094**** | 0.271**** | 0.166** |
| Pollen/Flower | −0.08 | −0.024 | −0.175* | −0.025** | 0.156 | 0.018 |
| Plant height | 0.143**** | 0.189**** | 0.307* | 0.084**** | 0.115*** | 0.316* |
| Number of stalks | 0.126**** | 0.039** | 0.147**** | 0.007 | 0.149**** | 0.128** |
| Stalk diameter | 0.148**** | 0.025 | 0.204** | 0.065**** | 0.163**** | 0.111 |
| Number of flowers | 0.138**** | 0.06**** | 0.191**** | 0.071**** | 0.155**** | 0.141*** |
| Petal length | 0.014 | 0.021 | −0.04 | 0.039* | 0.117** | 0.009 |
| Corolla tube diameter | 0.022 | 0.055* | −0.121 | 0.057*** | 0.159** | 0.014 |
| Corolla length | 0.11 | 0.023 | −0.106 | 0.032 | 0.138*** | 0.041 |
| Long filament length | 0.186** | 0.016 | 0.323 | 0.042* | 0.245*** | 0.09 |
| Short filament length | 0.116* | −0.035 | 0.161 | 0.043** | 0.135** | 0.072 |
| Style length | 0.15* | 0.041 | 0.163 | 0.022 | 0.199** | 0.104* |
| Herkogamy | −0.01 | 0.027 | −0.057 | −0.034 | −0.04 | 0.134* |
| Ploidy | Heterozygosity | Minimum | Maximum | Reduced |
|---|---|---|---|---|
| Diploid | PINT | 2.207 | 2.171 | 1.66 |
| PINTc | 2.056 | 1.468 | 1.197 | |
| 95% CI | (1.923, 3.197) | (2.157, 3.425) | (1.683, 2.583) | |
| RelPINT | 11.617 | 11.429 | 8.736 | |
| Tetraploid | PINT | 2.618 | 1.781 | 1.903 |
| PINTc | 2.534 | 1.253 | 1.767 | |
| 95% CI | (2.349, 3.084) | (1.802, 2.658) | (1.708, 2.663) | |
| RelPINT | 13.781 | 9.372 | 10.017 | |
| Hexaploid | PINT | 2.187 | 2.567 | 2.925 |
| PINTc | 1.89 | 2.114 | 2.165 | |
| 95% CI | (1.921, 3.054) | (2.157, 3.711) | (2.705, 4.52) | |
| RelPINT | 11.511 | 13.51 | 15.396 |
- —H2020 Marie Skłodowska‐Curie Actions10.13039/100010665
- —Vicerrectorado de Investigación y Transferencia, Universidad de Granada10.13039/501100018947
- —Ministerio de Ciencia, Innovación y Universidades10.13039/100014440
- —Organismo Autónomo de Parques Nacionales10.13039/100015865
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic diversity and population structure · Plant and animal studies · Chromosomal and Genetic Variations
Introduction
1
Mating systems in flowering plants range from self‐pollination (selfing) to cross‐pollination (outcrossing), with these two extremes considered the most evolutionarily stable strategies (Laenen et al. 2018). However, in many predominantly selfing species, individuals can still receive pollen from unrelated conspecifics, allowing for occasional outcrossing. A shift in the mating system may occur if outcrossed offspring exhibit higher fitness than those produced by selfing—a scenario studied by simulations in contact zones between formerly isolated lineages (Harkness et al. 2019). Hence, following the terminology used by Whitlock et al. (2013), we define outbreeding response as the net phenotypic or fitness change observed in the offspring produced by crossing two genetically distinct selfing individuals. Since selfing individuals are typically highly inbred and exhibit low heterozygosity, any outbreeding response is largely driven by heterozygosity increases (Kumar et al. 2020). This response may lead to either heterosis or outbreeding depression, depending on whether the fitness effects are positive or negative (Oakley et al. 2015; Soto et al. 2023). Notably, both outcomes can occur across different traits or generations within the same lineage, as the strength and direction of the response depend on complex genetic, epigenetic, phenotypic, and environmental interactions (Govindaraju 2019). Ultimately, the key evolutionary question centers on the net effect of outcrossing on fitness, and the adaptive relevance of outbreeding response remains incompletely understood (Frankham 2015).
While outbreeding effects have been widely examined in agricultural systems (Labroo et al. 2021; Hochholdinger and Baldauf 2018; Liu et al. 2022), their evolutionary implications in natural systems remain unclear. Despite being a topic of interest since Darwin (1876), the evolutionary role of outbreeding response remains elusive due to its dependence on genetic background, ecological context, and especially genetic distance between parents (Van Dooren 2000). In general, outbreeding response tends to increase with parental genetic distance, though excessive divergence can result in hybrid breakdown due to incompatibilities (Rehman et al. 2021). Conversely, crosses between closely related yet inbred individuals may exhibit heterosis due to masked deleterious alleles (Rhode and Cruzan 2005). This gives rise to the concept of an “optimal outcrossing distance,” where fitness benefits are maximized while both inbreeding and outbreeding depression are avoided (Edmands 1999; Chung et al. 2023).
Importantly, not all traits respond to outbreeding in the same way. Traits not directly related to fitness often show stronger heterotic effects, while fitness‐related traits tend to exhibit weaker or even negative responses, likely due to their complex epistatic interactions and reduced heritability (Söderquist et al. 2020; Whitlock et al. 2013). Moreover, differences in how fitness is measured—that is whether via growth, reproduction, or survival—can influence interpretations of heterosis and outbreeding depression. From an evolutionary standpoint, it is essential to focus not merely on phenotypic changes but on their relationship with reproductive success and long‐term adaptability too.
Polyploidy is widespread in plant lineages and adds another layer of complexity to outbreeding responses (Renny‐Byfield and Wendel 2014). Polyploids tend to harbor higher levels of heterozygosity, exhibit stronger heterotic effects (Chen 2010), and display altered gene expression and trait variance compared to diploids (Riddle et al. 2010; Bansal et al. 2012; Washburn and Birchler 2014). These genomic characteristics influence phenotypic integration—the degree to which traits co‐vary—and may contribute to wider phenotypic spaces and greater evolutionary potential (López‐Jurado et al. 2019). Additionally, heterozygosity increases and altered gene interactions following outcrossing can weaken trait covariation patterns, further reshaping phenotypic architecture (Balao et al. 2011). Theoretical models suggest that phenotypic integration is positively associated with inbreeding depression. As well, integration is negatively correlated with the potential for heterosis (Vasseur et al. 2019). This would imply that low integration may facilitate transitions in fitness landscapes.
The Erysimum incanum complex provides a compelling system to investigate how outbreeding responses are modulated by ploidy and trait architecture. This selfing species, native to the Iberian Peninsula and Morocco, exhibits diploid, tetraploid, and hexaploid cytotypes (Nieto‐Feliner et al. 1993). Its short monocarpic life cycle facilitates the measurement of trait variation and fitness across generations. Furthermore, polyploidy within E. incanum allows for the exploration of how genome duplication alters the adaptive consequences of occasional outcrossing in a predominantly selfing background.
If outbreeding response leads to higher fitness in cross‐fertilized offspring, it may act as a catalyst for transitions from selfing to outcrossing. In turn, the genomic flexibility conferred by polyploidy may amplify this potential by enabling more diverse phenotypic outcomes (Soltis and Soltis 2009). Furthermore, reduced phenotypic integration may facilitate movement through new adaptive landscapes. To test these ideas, we (1) quantify outbreeding response, (2) evaluate how ploidy and trait modules affect its magnitude, (3) investigate the relationship between phenotypic integration and outbreeding response, and (4) assess whether outbreeding response acts as a selection target.
To this end, we performed controlled crosses among genetically distinct selfing maternal lines of E. incanum , belonging to the same population and ploidy level, to generate outcrossed offspring with increased heterozygosity. These were compared with offspring produced via self‐fertilization. Additionally, we established second‐generation selfed lines from the outcrossed individuals to evaluate trait reversibility and persistence. Through this framework, we aim to determine whether outbreeding enhances fitness and may thus contribute to evolutionary shifts in mating systems.
Material and Methods
2
Study System
2.1
Erysimum L. is a highly diverse genus within the Brassicaceae family, distributed across Eurasia, North Africa, and parts of North and Central America (Al‐Shehbaz et al. 2006). One of its species, Erysimum incanum (Figure 1), is considered a species complex, comprising annual and monocarpic taxa and subspecies inhabiting the Western Mediterranean Basin (Nieto‐Feliner et al. 1993; Abdelaziz et al. 2019). This complex includes three ploidy levels: diploids (2n = 2x = 16 chromosomes), tetraploids (2n = 4x = 32), and hexaploids (2n = 6x = 48) (Nieto‐Feliner et al. 1993; Abdelaziz, M and Muñoz‐Pajaers, A.J. in prep.). Diploids occur in a vicariant distribution across the Rif and Pyrenees Mountains (Figure 1b), while tetraploids are found in the southwestern Iberian Peninsula and the Middle Atlas Mountains (Nieto‐Feliner et al. 1993; Fennane and Ibn‐Tattou 1999). In contrast, hexaploids are restricted to the southernmost ranges of Morocco (High Atlas and Anti‐Atlas; Abdelaziz et al., in prep.). The E. incanum complex is predominantly autogamous. Its flowers are small, hermaphroditic, self‐compatible, and exhibit an anther‐rubbing mechanism that promotes efficient self‐pollination (Abdelaziz et al. 2019).
Experimental framework to study outbreeding response. (a) Erysimum incanum flowers and representative individuals of each ploidy level. (b) Geographic distribution of E. incanum populations across the Iberian Peninsula and Morocco. (c) Conceptual workflow of the experimental design. Is outbreeding response a mechanism that promotes outcrossing? If a positive outbreeding response increases fitness, then outcrossing may be favored over selfing. This response may be influenced by ploidy level and trait modularity. The effect of cross‐fertilization between selfing parental lines could vary depending on ploidy and trait module, and the associated fitness changes may favor a shift toward either selfing (negative response) or outcrossing (positive response). (d) Experimental design of greenhouse crosses to generate offspring with different heterozygosity levels. We established three experimental groups: (1) selfed offspring with null heterozygosity, derived from self‐pollination; (2) outcrossed offspring with maximum heterozygosity, resulting from controlled cross‐fertilizations between inbred lines; and (3) selfed offspring from the previous outcrossed group, displaying intermediate heterozygosity, produced via controlled self‐pollination.
Experimental Design and Phenotypic Measurements
2.2
We sowed seeds from six wild populations (Table 1) in the greenhouse facilities in the Faculty of Sciences (University of Granada) for three consecutive generations to reduce residual heterozygosity resulting from potential previous outcrossing events. We then conducted controlled crosses among different maternal lines within each population to generate three heterozygosity groups: (i) null heterozygotes, derived from selfing; (ii) maximum heterozygotes, produced by controlled outcrossing between inbred lines; and (iii) reduced heterozygotes, obtained by selfing the maximum heterozygote offspring.
For each ploidy level, we first intercrossed highly inbred maternal lines to generate maximum heterozygotes. Finally, we performed controlled selfing on maximum heterozygotes to obtain reduced heterozygotes (Figure 1). In total, we obtained 405 diploid plants (257 null, 53 maximum, and 95 reduced heterozygotes), 764 tetraploid plants (457 null, 69 maximum, and 238 reduced heterozygotes), and 453 hexaploid plants (252 null, 112 maximum, and 89 reduced heterozygotes).
We measured flower traits during anthesis using one flower per plant and a digital caliper (±0.1 mm accuracy; Dexter, Alcobendas, Madrid). Measured traits included petal length (maximum length from the edge to the curving point), corolla diameter (distance between opposite petals), and corolla tube length (distance from sepal base to the corolla opening). We also measured the lengths of long and short filaments and the height of the style (from its base to the stigma), allowing us to calculate herkogamy (i.e., the vertical distance between long stamens and the style).
To assess male reproductive investment, we collected half of the stamens per flower and preserved them in 100 μL of 70% ethanol. We estimated pollen quantity per flower using a Multisizer Coulter Counter 3 (Beckman Coulter, Pasadena, USA), provided by the morphometric laboratory at the Centro de Investigación, Tecnología e Innovación (CITIUS), University of Sevilla (Spain).
We collected mature fruits and counted viable seeds, aborted seeds, and unfertilized ovules per fruit to estimate female reproductive investment. We also evaluated seed germination rate by sowing 10 seeds per pot (per heterozygosity group) and calculating the proportion that germinated. Additional measured variables included the number of leaves and fruits per plant, the number of seeds per fruit (average from four fruits), the height of the main flowering stalk, the number and diameter of all stalks, and the total number of flowers per plant.
Statistical Analysis
2.3
Outbreeding Response
2.3.1
We grouped traits into four modules: (i) individual traits (height, number of stalks, stalk diameter, and number of flowers), (ii) flower traits (petal length, flower diameter, corolla length, long filament, short filament, and herkogamy), (iii) reproductive traits (mean number of pollen grains and ovules per flower), and (iv) fitness components (total pollen and ovule output per plant, fruit set, seed set, germination, survival, and number of leaves). We conformed the groups considering size traits not directly related to reproduction for the individual module, traits related to flower morphology for the flower module, the reproductive module as the initial stage of reproduction (gamete production) for the reproductive module, and the pre‐dispersive and post‐dispersive fitness measurements for the fitness components. We conducted statistical analysis with R 4.4.3 software (R Core Team 2022).
We calculated outbreeding response (OR) using a modified version of the inbreeding depression index (Johnston and Schoen 1994):
where μ₀ represents a given mean trait value in null heterozygotes and μᵢ the mean of this same trait in heterozygous individuals (either maximum or reduced). This index, ranging from −1 (maximum outbreeding depression) to +1 (maximum heterosis), allows standardized comparisons across traits and ploidy levels. We chose this approach instead of the classical formulae to measure heterosis because we wanted to study by the whole groups and we did not track the genealogy of each line. As well, our interest was in ploidy and heterozygosity levels and not in the family level. To assess the statistical significance of outbreeding responses, we used bootstrap resampling (n = 100,000) to calculate 95% confidence intervals.
To test whether outbreeding response varied by ploidy level or trait modules, we conducted a generalized linear mixed model (GLMM) test for each heterozygosity level. We set ploidy and trait modules as fixed effects and traits as a random effect. We did this type of analysis as we cannot obtain an individual outbreeding response itself, as we do not have data for different heterozygosity levels either for individuals or families. However, this approach allows us to analyze the short data size we finally have for it (20 traits within each ploidy level divided in four trait modules).
Linear Regression of Individual Contribution to Outbreeding Response on Fitness
2.3.2
To evaluate whether outbreeding responses are subject to natural selection, we calculated each individual plant's contribution to outbreeding response by comparing its trait values (under maximum or reduced heterozygosity) with the population mean under null heterozygosity. We used the same formulae (Equations 1 and 2) and then normalized the output using:
This transformation scales values from (−1,1) to (0,1), which allows interpreting contributions in a positive range. Normalization allows to continue studying the sense of outbreeding response in the same range between −1 and 1. As we used the outbreeding response for each group to get the individual measurement, this avoids having both negative values in the ratio between the individual and the group level (see Appendix Table A1 to see a detailed development). We calculated fitness as the absolute product of fruit production and seed production, and we normalized it by the higher value in every ploidy level. For each trait, we performed linear regressions between the individual contribution to outbreeding response and fitness. As this fitness can be read as pre‐dispersive fitness, we run the same analysis using a post‐dispersive fitness obtained from the product of survival rate, germination rate, and leave number.
Hence, individual contribution to outbreeding response gives the opportunity to study its relationship with fitness as we get one value per individual. This newly proposed analysis differs from regular correlations between fitness and trait values as the study element is the measurement of the outbreeding response itself, which is, the magnitude of the variation and not the value of the given trait. A certain trait can be related to fitness, but its outbreeding response may not.
Phenotypic Integration
2.3.3
We assessed phenotypic integration by quantifying the number of significant pairwise correlations among traits. We grouped individuals by ploidy (diploid, tetraploid, hexaploid) and heterozygosity level (null, maximum, reduced). For each group, we calculated the number of significant correlations involving each trait and derived the average number of correlations per trait–group combination.
To assess this global integration for each group, we obtained a phenotypic integration index based on Wagner (1984) using PHENIX 1.3.1 Package in R (Torices and Muñoz‐Pajares 2015). We also calculated the corrected version of this index by extracting the possible amount of integration due to random covariance, as well as a proportion of integration regarding the highest potential level of integration. Significance was calculated using the bootstrapping method at 10,000 replaces provided by this same R package.
Results
3
We evaluated the effect of cross‐fertilization on phenotypic traits in E. incanum lines differing in ploidy (diploid, tetraploid, and hexaploid) and heterozygosity level (maximum, null, and reduced). Table 2 shows the outbreeding response across trait modules and heterozygosity levels, so that:
Outbreeding Response for Maximum Heterozygosity
3.1
Tetraploids exhibited the highest number of traits with significant outbreeding responses (14 out of 20), of which 13 were positive. In contrast, diploids and hexaploids each showed only four traits with significant responses: all four were positive in hexaploids, while three of the four were negative in diploids (Table 2).
The significant responses observed in diploids and hexaploids were not always shared. All four positive traits in hexaploids were also significantly positive in tetraploids, with larger effect sizes in tetraploids. The only significant response shared between diploids and tetraploids was for germination, which was negative in both; two traits (survival and number of leaves) were significant only in diploids—survival positive and leaves negative—while petal length showed opposing significant responses in diploids (negative) and tetraploids (positive).
Considering trait modules, no fitness component showed a significant response in hexaploids, while the mean number of ovules per plant, the mean number of pollen per plant and the seed set were positively significant only in tetraploids. Survival was significant (positive) only in diploids. Germination was significantly negative both in diploids and tetraploids; the number of leaves per plant was significantly negative only in diploids, and the seed set was the only fitness component trait not showing a significant outbreeding response at any ploidy level.
In the reproductive trait module, both pollen and ovule production per flower showed a significant positive response in tetraploids and hexaploids but not significant in diploids.
For individual traits, only stalk diameter and plant height showed a significant positive response and only in tetraploids. The number of stalks and the number of flowers showed no significant response at any ploidy level.
In the flower trait module, long filament and style length had congruent positive responses in tetraploids and hexaploids. Flower diameter, corolla tube length, and short filament length showed positive responses exclusively in tetraploids. Petal length had opposing responses in diploids and tetraploids. Herkogamy showed no significant outbreeding response in any ploidy level.
Notably, no trait exhibited a significant response across all three ploidy levels. Traits such as seed set, number of stalks, number of flowers, and herkogamy did not show any significant response in any group.
Outbreeding Response for Reduced Heterozygosity
3.2
Diploids exhibited the highest number of traits with significant negative outbreeding responses (14 of 16 significant traits; Table 3). Tetraploids and hexaploids each had 12 traits showing significant response, with 7 being negative in both—but affecting different traits in each. Most traits that show a significantly negative response in diploids were either also negative or non‐significant in the other two ploidy levels. However, flower diameter showed a significantly positive response in both tetraploids and hexaploids, despite being close to zero.
For individual traits, stalk diameter and number of flowers showed significantly positive responses in diploids and tetraploids only. Reproductive traits showed consistently significant negative responses across all ploidy levels. In contrast, flower traits showed positive significant responses in hexaploids for three traits. Regarding fitness‐related traits, ovule and pollen production per plant, fruit set, and seed set all had significantly negative responses. In contrast, traits linked to survival showed positive significant responses in one or both polyploid levels (e.g., germination in hexaploids, survival in tetraploids, and number of leaves in both).
Effects of Ploidy and Trait Module on Outbreeding
3.3
A generalized linear mixed model (GLMM) was fitted to assess the effects of ploidy and module on outbreeding response (Figure 2), with trait included as a random effect. We executed this analysis for both heterozygosity levels.
Variation in outbreeding response across trait modules and ploidy levels in Erysimum incanum under maximum and reduced heterozygosity. Each color corresponds to a different trait module. Dots represent outbreeding response values for individual traits, while central dots indicate the mean response for each trait module. Significance was assessed using the Scheirer–Ray–Hare test. (a) Outbreeding responses under maximum heterozygosity. (b) Outbreeding responses under reduced heterozygosity.
For the maximum heterozygosity level, variance among traits was minimal (Variance = 0.016; SD = 0.125), indicating that most of the variation in the index occurred within traits. Among the fixed effects, tetraploidy had a significant positive effect on outbreeding response compared to diploid level (β = 0.232 ± 0.039, t = 5.88, p < 0.001), whereas hexaploidy did not differ significantly from diploids (β = 0.062 ± 0.039, t = 1.57, p > 0.1). Regarding trait modules, none had a statistically significant effect relative to the reference module. The reproductive module showed a moderate positive trend (β = 0.192 ± 0.115, t = 1.66, p≈0.1) but was not significant.
For the reduced heterozygosity level, both tetraploid and hexaploid levels significantly increased outbreeding response compared to diploids (β = 0.163 ± 0.047, t = 3.45; β = 0.140 ± 0.047, t = 2.97, respectively). Module type had generally no significant effect, except for the reproductive module, which showed a significant negative effect (β = −0.479 ± 0.166, t = −2.88). Random variation among traits was moderate (Var = 0.0355; SD = 0.188), indicating some heterogeneity between traits, but the majority of variation occurred within traits.
Fitness and Outbreeding Response
3.4
We estimated the individual contribution to outbreeding response for each trait, heterozygosity, and ploidy level to evaluate its relationship with fitness (Table 4). We found that all significant relationships were positive, except for survival in diploids with reduced heterozygosity, seed germination in diploids and hexaploids with reduced heterozygosity and herkogamy for tetraploids with reduced heterozygosity. The traits with the most significant relationships were the total amount of ovules, the number of flowers, and the total amount of pollen per plant. Our results show that a higher proportion of individual contribution to outbreeding response in these traits means a higher individual fitness. We also included results for post‐dispersive fitness to enrich the viewpoint but the majority of the regressions were not significant (see Appendix Table A2).
Phenotypic Integration
3.5
We estimated trait integration and evaluated the effects of heterozygosity and ploidy levels (Figure 3a, see Appendix Figure A1 for separated traits). Overall, outcrossed offspring showed less integrated phenotypes compared to selfed individuals (Figure 3b). However, our study of phenotypic integration indices (Table 5) shows that, regarding bootstrapping values, there were not significant differences between heterozygosity levels within each ploidy. Notwithstanding, we observed integration levels according to this index were higher for the null heterozygosity group except at the hexaploid level (Table A2).
Patterns of phenotypic integration in Erysimum incanum. (a) Total number of significant pairwise correlations (i.e., integrations) among traits. (b) Mean number of significant correlations per phenotypic trait. Data are grouped by ploidy level and heterozygosity class. Significance was tested for differences between heterozygosity levels within each ploidy group using t‐tests. Levels within a ploidy that do not share a letter are significantly different at p < 0.05.
We observed a generally negative relationship between trait integration and heterozygosity level. Null heterozygosity plants had the highest number of significant trait correlations across all ploidy levels. At higher heterozygosity, diploids and tetraploids showed reduced integration; but in hexaploids, integration remained similar to null heterozygotes. These results are globally in agreement with phenotypic integration index data. Even at the hexaploid level, this average trait correlation was similar for null and maximum heterozygosity levels, confidence intervals were completely overlapping.
Selfed tetraploids exhibited the highest number of significant correlations overall. Interestingly, while increased heterozygosity reduced integration in diploids and tetraploids, hexaploids maintained stable integration levels, suggesting a potential buffering effect of higher ploidy.
Discussion
4
Outbreeding response has been extensively studied in the context of agriculture and breeding (Paril et al. 2024; Schnable and Springer 2013). However, it may also play a key role in evolution, potentially acting as a driver of outcrossing when it results in enhanced offspring fitness. Erysimum incanum, a selfing species within Brassicaceae, is highly inbred and comprises populations with varying ploidy levels—diploid, tetraploid, and hexaploid.
To evaluate the impact of outcrossing on fitness, we performed controlled crosses between distinct selfing lines. We found a significantly positive outbreeding response (i.e., heterosis) in tetraploids after a single outcrossing event. This was followed by a decrease in heterozygosity in subsequent selfed offspring, which was often associated with outbreeding depression. Overall, trait modules and ploidy levels explained a greater proportion of variation than heterozygosity level per se. Notably, we also detected a direct positive relationship between outbreeding response and fitness. In parallel, heterozygosity was inversely correlated with phenotypic integration across all ploidy levels. These findings suggest that outbreeding response may play a role in promoting transitions between mating systems.
A meta‐analysis by Whitlock et al. (2013) found that outbreeding tends to negatively affect fitness‐related traits. However, other studies, especially in polyploids, have shown fitness benefits following outcrossing between genetically distinct populations (Rodger et al. 2024). In our case, within‐population outcrosses between selfing lines also enhanced fitness components. Specifically, tetraploid outcrossed individuals produced a higher total number of pollen grains and ovules and exhibited greater reproductive allocation per flower than their selfed counterparts.
Although fitness components are diverse and often context‐dependent (Vasseur et al. 2019), our findings contrast with predictions that polyploidization promotes selfing and asexuality. Instead, they support growing evidence that polyploidy can expand phenotypic potential (Clo 2022), enhancing heterotic effects (Chen 2010). Still, increased phenotypic variation may not always translate into fitness benefits (Hedrick 2012). For instance, while traits such as plant size or architecture may benefit from greater plasticity (Fort et al. 2016), hexaploids—with even broader phenotypic space—may face greater difficulty achieving optimal fitness peaks. The underlying mechanisms are complex (Mackay et al. 2021), but phenotypic expression remains a valuable proxy for exploring the functional consequences of outcrossing and genome duplication (Lippman and Zamir 2007).
In diploids, outbreeding response at maximum heterozygosity was not statistically significant, although it tended to be negative across most traits. This contrasts with the predominantly positive responses observed in tetraploids and hexaploids. The inverse trend in diploids may reflect fundamental differences in the genetic basis of heterosis and outbreeding depression. Specifically, interactions among alleles and regulatory networks may be more susceptible to disruption in diploids than in polyploids. Increased heterozygosity can destabilize well‐established genetic architectures in inbred backgrounds (Fridman 2015). Furthermore, long‐term divergence among selfing diploid lineages may lead to the accumulation of cryptic genetic incompatibilities, resembling those observed in between‐population crosses (Clo et al. 2021). This effect is expected to be less pronounced in polyploids, where additional chromosome sets may buffer against allelic incompatibilities.
The subsequent reduction in heterozygosity through selfing of previously outcrossed diploids resulted in outbreeding depression, reinforcing a stabilization of the selfing strategy and making reversal toward outcrossing less likely. While in outcrossing offspring deleterious mutations may be hidden from natural selection, in selfing offspring mildly deleterious mutations can be exposed to it. Even if they are different alleles for the same locus, if they coincide, this event would be in a broad sense in line with Bateson‐Dobzhansky‐Muller incompatibilities (Pickup et al. 2019). In this sense, the contrast between the initial positive response of the maximum heterozygosity group and the decline observed when heterozygosity is reduced reflects the interplay between heterosis, the inefficiency of purging, and hybrid breakdown, reinforcing the interpretation that early generation fitness gains often give way to later generation depression (Ågren and Sletvold 2025). In contrast, the positive outbreeding response in polyploids suggests that purging of deleterious alleles has not been fully effective, allowing beneficial heterozygous combinations to persist. This incomplete purging may be particularly relevant for traits contributing to fitness, such as flower productivity and gamete output.
Ploidy emerged as the most influential factor explaining variation in outbreeding response. While polyploidy initially narrows the scope of heritable variation, this effect tends to be transient (Clo 2022). The trait module was also a significant predictor, particularly at the reduced heterozygosity level. Trait modules not only reflect functional groupings but also shared developmental pathways and gene networks (Armbruster et al. 2014), making them more likely to exhibit coordinated responses to genetic perturbations.
For example, flower traits often exhibited more consistent and directional outbreeding responses than individual traits. This is likely due to their significant role in reproductive success, which subjects them to stronger selective constraints and lower additive genetic variance (Whitlock et al. 2013). In contrast, individual traits are typically more plastic and influenced by abiotic conditions (Opedal et al. 2023). These differences in genetic architecture and selective regimes may explain why some modules are more responsive to outbreeding than others.
Fitness was positively associated with outbreeding response for multiple fitness‐related traits, including total ovule and pollen production and the number of leaves (Rhode and Cruzan 2005). This relationship was also evident at the reduced heterozygosity level (Table 2), particularly in diploids, where decreased fitness accompanied reduced heterozygosity, suggesting that outcrossing benefits may persist even after a subsequent selfing event. Individuals exhibiting greater positive outbreeding effects had higher reproductive success, which could favor the spread of outcrossing genotypes. Because the relationship between outbreeding and reproductive performance is positive, outbreeding depression following the loss of heterozygosity may result in reduced fitness, reinforcing the advantage of maintaining or restoring heterozygosity through outcrossing.
Although outbreeding response and heterozygosity are conceptually distinct, both can influence fitness (David 1998). While outbreeding response refers to the net change in trait values following outcrossing, its evolutionary relevance depends on its fitness consequences. In our study, the consistent positive effect of outbreeding response on fitness highlights its potential evolutionary importance. Moreover, even short‐term increases in fitness could benefit populations during colonization or environmental transitions (Barker et al. 2019). To our knowledge, this is the first study to explicitly explore the link between outbreeding response and multiple fitness components across ploidy levels and trait categories.
Phenotypic integration decreased with increasing heterozygosity in diploids and tetraploids, but not in hexaploids. Trait integration is shaped by underlying genetic correlations, which are disrupted by heterozygosity. The relative magnitude of heterozygosity increases after the decrease of outcrossing with the increase of ploidy level—being greater in diploids than in tetraploids, and lowest in hexaploids. Even so, all heterozygosity levels showed reduced integration relative to null heterozygosity groups. In highly inbred populations, recombination is limited, and allelic combinations are preserved over generations. This stability promotes higher trait integration, particularly in selfing species, where flower integration is a hallmark of selfing syndromes (Fornoni et al. 2016). Disruption of long‐standing genetic linkages through heterozygosity may reduce trait integration, potentially allowing novel phenotypic combinations. Such effects have been observed in Arabidopsis, where admixture between divergent genotypes produced transgressive trait variation (Palacio‐Lopez and Molofsky 2021). Interestingly, despite their broader combinatorial potential, hexaploids maintained relatively high trait integration, possibly related to gene redundancy which keeps more stable development processes (Birchler and Veitia 2010; Mable et al. 2018). This functional integration in hexaploids may act as a constraint on phenotypic space compared to tetraploids.
According to simulation studies (Harkness et al. 2019), heterosis alone may be insufficient to drive mating system transitions, as its effects tend to be transient. These models suggest that long‐term shifts from selfing to outcrossing require additional evolutionary forces. Still, heterosis may act in synergy with other selective processes, contributing to transitions under the right ecological and genetic conditions (Draghi and Whitlock 2015). In our study, outcrossing occurred between selfing lines within populations—likely involving limited genetic divergence—so the magnitude of heterosis was expected to be modest. Nevertheless, the observed effects highlight the fine‐scale influence of outbreeding response, even among closely related genotypes.
Our results provide empirical evidence that ploidy variation modulates outbreeding response, with consequences for fitness and evolutionary potential. Differences in reproductive outcomes among cytotypes suggest that both heterosis and outbreeding depression are not fixed properties but are shaped by genomic context, including gene expression patterns, epigenetic regulation, and developmental buffering (Comai 2005; Chen 2007). These findings underscore how polyploidy can reshape genetic architecture, modulating the trade‐off between genetic diversity and reproductive efficiency, and ultimately influencing plant adaptation and diversification (Otto and Whitton 2000; Van de Peer et al. 2017; Soltis et al. 2014).
Overall, our study demonstrates a stronger outbreeding response in tetraploids of Erysimum incanum. The positive relationship between outbreeding response and fitness, combined with lower phenotypic integration, suggests that tetraploids occupy a broader phenotypic space than diploids. This flexibility may enhance their capacity to adapt to new or fluctuating environments. While selfing offers short‐term reproductive assurance, it may limit long‐term evolutionary potential (Igic and Busch 2013). Outcrossing becomes adaptive when it results in higher‐fitness offspring, thereby favoring a shift in mating system. However, for such a shift to have an evolutionary impact, it must be stabilized within populations over time. This work provides a first step toward understanding the evolutionary dynamics of mating system transitions in Erysimum. Future research will aim to explore additional factors that could promote outcrossing and determine whether such transitions can be maintained in natural populations.
Author Contributions
Carlos Olmedo‐Castellanos: data curation (equal), formal analysis (lead), investigation (equal), methodology (equal), writing – original draft (equal), writing – review and editing (equal). Ana García‐Muñoz: data curation (equal), investigation (equal), methodology (equal), writing – original draft (equal). Camilo Ferrón: data curation (equal), methodology (equal), writing – original draft (equal), writing – review and editing (equal). Mohammed Bakkali: investigation (equal), methodology (equal), writing – original draft (equal), writing – review and editing (equal). A. Jesús Muñoz‐Pajares: conceptualization (equal), funding acquisition (equal), investigation (equal), methodology (equal), supervision (equal), writing – original draft (equal), writing – review and editing (equal). Mohamed Abdelaziz: conceptualization (equal), formal analysis (equal), funding acquisition (equal), investigation (equal), methodology (equal), project administration (equal), supervision (equal), writing – original draft (equal), writing – review and editing (equal).
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abdelaziz, M. , M. Bakkali , J. M. Gómez , E. Olivieri , and F. Perfectti . 2019. “Anther Rubbing, a New Mechanism That Actively Promotes Selfing in Plants.” American Naturalist 193, no. 1: 140–147. 10.1086/700875.30624113 · doi ↗ · pubmed ↗
- 2Ågren, J. , and N. Sletvold . 2025. “Plant Mating Systems in Small Populations.” Annual Review of Ecology, Evolution, and Systematics 56: 445–466. 10.1146/annurev-ecolsys-110421-102855. · doi ↗
- 3Al‐Shehbaz, I. A. , M. A. Beilstein , and E. A. Kellogg . 2006. “Systematics and Phylogeny of the Brassicaceae (Cruciferae): An Overview.” Plant Systematics and Evolution 259: 89–120. 10.1007/s 00606-006-0415-z. · doi ↗
- 4Armbruster, W. S. , C. Pélabon , G. H. Bolstad , and T. F. Hansen . 2014. “Integrated Phenotypes: Understanding Trait Covariation in Plants and Animals.” Philosophical Transactions of the Royal Society, B: Biological Sciences 369, no. 1649: 20130245. 10.1098/rstb.2013.0245.PMC 408453325002693 · doi ↗ · pubmed ↗
- 5Balao, F. , J. Herrera , and S. Talavera . 2011. “Phenotypic Consequences of Polyploidy and Genome Size at the Microevolutionary Scale: A Multivariate Morphological Approach.” New Phytologist 192: 256–265. 10.1111/j.1469-8137.2011.03787.x.21651562 · doi ↗ · pubmed ↗
- 6Bansal, P. , S. Banga , and S. S. Banga . 2012. “Heterosis as Investigated in Terms of Polyploidy and Genetic Diversity Using Designed Brassica juncea Amphiploid and Its Progenitor Diploid Species.” P Lo S One 7, no. 2: e 29607. 10.1371/journal.pone.0029607.22363404 PMC 3283606 · doi ↗ · pubmed ↗
- 7Barker, B. S. , J. E. Cocio , S. R. Anderson , et al. 2019. “Potential Limits to the Benefits of Admixture During Biological Invasion.” Molecular Ecology 28, no. 1: 100–113. 10.1111/mec.14958.30485593 PMC 6344275 · doi ↗ · pubmed ↗
- 8Birchler, J. A. , and R. A. Veitia . 2010. “The Gene Balance Hypothesis: Implications for Gene Regulation, Quantitative Traits and Evolution.” New Phytologist 186: 54–62. 10.1111/j.1469-8137.2009.03087.x.19925558 PMC 2858765 · doi ↗ · pubmed ↗