MED27 function is essential for cerebellar development and motor behaviour
Sabrina Maher, Eloise Langlois Bernard, Charlotte Zaouter, Shunmoogum A Patten

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
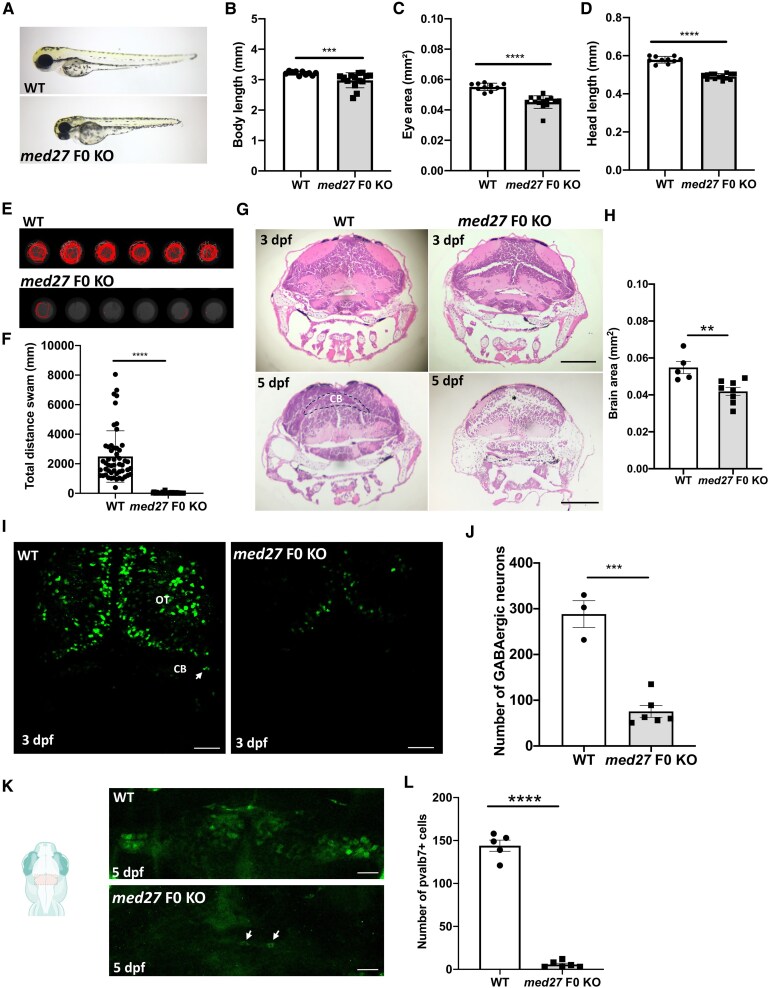 Figure 1
Figure 1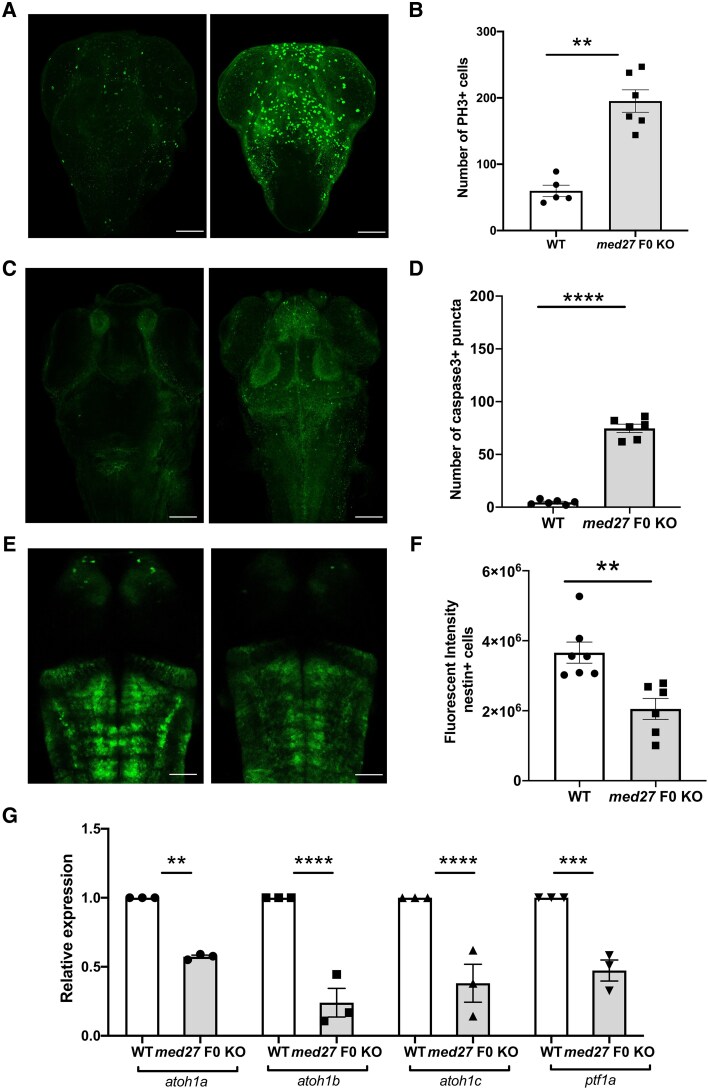 Figure 2
Figure 2- —Canadian Institutes of Health Research10.13039/501100000024
- —FRQS Masters Scholarship
- —Anna Sforza Djoukhadjian Research Chair
- —Fondation Armand-Frappier10.13039/100012618
- —Natural Science and Engineering Research Council10.13039/501100005049
- —FRQS Junior 2 research scholar
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics and Neurodevelopmental Disorders
In the recent publication in Brain by Maroofian et al.,^1^ the authors provide a comprehensive clinical and radiological description of MED27-related disease. They particularly demonstrated that MED27-related disease manifests brain anomalies along with movement abnormalities. Several reports have also identified MED27 variants in a neurodevelopmental disorder characterized by key features such mental retardation, cerebellar atrophy, spasticity, hypotonia and motor deficits.^2-4^ Wu et al.^3^ showed that homozygous missense variants in the gene MED27 resulted in a significant reduction in MED27 protein expression. However, the pathogenicity of loss-of-function of MED27 and the mechanisms underlying neurodevelopmental defects in MED27-related disease currently remain largely unknown.
We investigated the role of MED27 in neurodevelopment using the zebrafish as a vertebrate model system. The zebrafish genome encodes a single med27 orthologue that is highly conserved with human MED27. We generated a med27 biallelic knockout (F0 KO) zebrafish using the CRISPR-Cas9 system.^5^ We found that med27 F0 KO larvae exhibited a smaller body, eye and head size compared to wild-type control fish (Fig. 1A–D). In light of the neurological phenotypes of patients harbouring MED27 variants, we evaluated various aspects of the morphology and function of the CNS in med27 F0 KO larvae. We investigated motor function in the med27 F0 KO fish using the automated Noldus Ethovision XT behaviour monitoring system. We found that med27 F0 KO exhibited significant motor impairments compared to controls (Fig. 1E and F). Haematoxylin and eosin (H&E) staining of transverse brain sections at 3 days post-fertilization (dpf) revealed structural differences and smaller brain size in med27 F0 KO fish compared to wild-type larvae at 3 dpf and 5 dpf (Fig. 1G and H). Importantly, we observed a marked progressive loss of H&E staining in the cerebellum of med27 F0 KO zebrafish brain compared to controls, suggesting a cellular depletion in the region (Fig. 1G). Taken together, these findings are in line with clinical observations in MED27 variant patients.
Given the brain abnormalities in MED27 patients and med27 F0 KO zebrafish, we next sought to examine the brain neuronal networks. We found a significant reduction in the number of GABAergic neurons in the med27 F0 KO zebrafish brain at 5 dpf (Fig. 1I and J), including in the cerebellum. These findings align with the clinical evidence of the loss of GABAergic neurons in several neurodevelopmental disorders including mental retardation,^6^ a key feature in MED27 variant patients. To complement our observations of cerebellar abnormalities and GABAergic neuronal loss in med27 F0 KO zebrafish, we assessed the cerebellar neuronal parvalbumin7 (pvalb7)-positive Purkinje cells which are important inhibitory GABAergic neurons for cerebellar function. We observed a significant reduction in the number of Purkinje cells within the cerebellum of med27 F0 KO zebrafish brain (Fig. 1K and L). Overall, these data suggest that med27 is involved in cerebellum development, particularly in populating GABAergic neuronal subtypes in vertebrates.
The zebrafish CNS proliferative profile is still very high at 2 dpf and is rapidly downregulated up to 5 dpf.^7^ In 5 dpf med27 F0 KO brains, we strikingly found an increase in proliferating cells (Fig. 2A and B) and apoptosis (Fig. 2C and D), suggesting a failure in neurogenesis with defects in progenitor cells differentiating into GABAergic neurons. Interestingly, we also found a significant reduction in the nestin-positive neural stem cells (Fig. 2E and F), reinforcing that MED27 likely plays a role during early neurogenesis, particularly in regulating the development of early neural progenitors. To confirm this hypothesis, we compared the expression level of specific cerebellar neurogenesis markers^8^ (atoh1a, atoh1b, atoh1c and ptf1a, for Purkinje cell neurogenesis^9,10^). We found that the expression of all these markers is significantly decreased at 2 dpf in med27 F0 KO larvae compared to controls by reverse transcription quantitative PCR (RT-qPCR) (Fig. 2G). These findings suggest that these changes caused by the loss of MED27 arise early on as of 2 dpf with important consequences on cerebellar neurogenesis at later developmental stages. Altogether, our data indicate an important role of MED27 in neurogenesis and cerebellar development.
Supplementary Material
awaf237_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Maroofian R, Kaiyrzhanov R, Cali E, et al Biallelic MED 27 variants lead to variable ponto-cerebello-lental degeneration with movement disorders. Brain. 2023;146:5031–5043.37517035 10.1093/brain/awad 257PMC 10690011 · doi ↗ · pubmed ↗
- 2Meng L, Isohanni P, Shao Y, et al MED 27 variants cause developmental delay, dystonia, and cerebellar hypoplasia. Ann Neurol. 2021;89:828–833.33443317 10.1002/ana.26019 · doi ↗ · pubmed ↗
- 3Wu G, Lian R, Li M, et al Novel homozygous missense variants in MED 27 associated with neurodevelopmental disorder: Clinical and pathogenetic research. Heliyon. 2024;10:e 37258.39296199 10.1016/j.heliyon.2024.e 37258 PMC 11409145 · doi ↗ · pubmed ↗
- 4Reid KM, Spaull R, Salian S, et al MED 27, SLC 6A 7, and MPPE 1 variants in a Complex neurodevelopmental disorder with severe dystonia. Mov Disord. 2022;37:2139–2146.35876425 10.1002/mds.29147 PMC 9796674 · doi ↗ · pubmed ↗
- 5De Pace R, Maroofian R, Paimboeuf A, et al Biallelic BORCS 8 variants cause an infantile-onset neurodegenerative disorder with altered lysosome dynamics. Brain. 2024;147:1751–1767.38128568 10.1093/brain/awad 427PMC 11068110 · doi ↗ · pubmed ↗
- 6Zhang A, Sokolova I, Domissy A, et al Maturation delay of human GAB Aergic neurogenesis in fragile X syndrome pluripotent stem cells. Stem Cells Transl Med. 2022;11:613–629.35556144 10.1093/stcltm/szac 022PMC 9216490 · doi ↗ · pubmed ↗
- 7Wullimann MF, Knipp S. Proliferation pattern changes in the zebrafish brain from embryonic through early postembryonic stages. Anat Embryol (Berl). 2000;202:385–400.11089930 10.1007/s 004290000115 · doi ↗ · pubmed ↗
- 8Pose-Mendez S, Schramm P, Valishetti K, Koster RW. Development, circuitry, and function of the zebrafish cerebellum. Cell Mol Life Sci. 2023;80:227.37490159 10.1007/s 00018-023-04879-5PMC 10368569 · doi ↗ · pubmed ↗