Genetic diversity and antimicrobial resistance profiles of Salmonella enterica in the broiler supply chain in Harare, Zimbabwe: tracking transmission from farm to table
Peter Katsande, Alistair R. Davies, Christian Owusu-Nyantakyi, Claudious Gufe, Shuvai Musari, Chenai S. Majuru, Jairus Machakwa

TL;DR
This study tracks Salmonella transmission in Zimbabwe's poultry supply chain, revealing high contamination rates and antimicrobial resistance genes from farms to retail markets.
Contribution
The study provides new genomic insights into Salmonella transmission and antimicrobial resistance in Zimbabwe's broiler supply chain.
Findings
Salmonella contamination rates were highest at slaughter facilities and retail markets compared to farms.
34.5% of isolates harbored resistance genes like fosA3 and qnrB19, with some showing extended-spectrum beta-lactamase production.
Genomic analysis revealed identical core SNPs across supply chain stages, indicating transmission pathways.
Abstract
Salmonella enterica is a significant zoonotic pathogen, posing a serious public health threat through its presence in the food supply chain, particularly in poultry production facilities. This study aimed to investigate the genetic diversity, antimicrobial resistance (AMR) profiles and phylogenetic relationships of S. enterica serovars isolated from various stages of the broiler supply chain in Harare, Zimbabwe. Whole-genome sequencing was employed to analyse 28 Salmonella isolates from broiler farms, slaughter facilities and retail markets. The overall prevalence of Salmonella was 5.1% out of 552 samples tested. Contamination rates were higher at slaughter facilities, where 11% of 100 samples tested positive and at retail markets, where 20% of 20 samples were contaminated. In contrast, farms had a significantly lower prevalence, with only 3.0% of 432 samples showing Salmonella…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
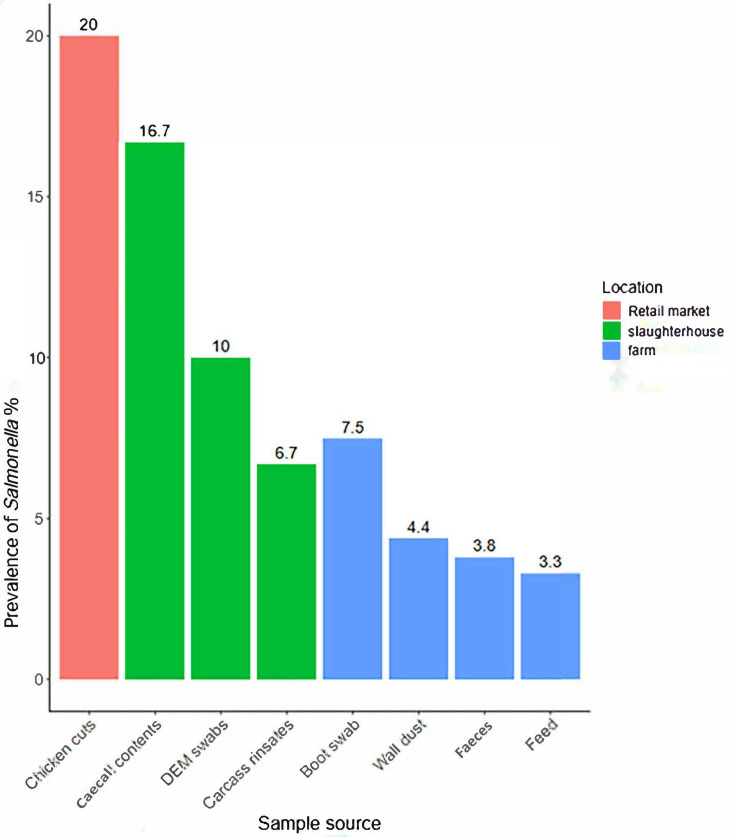 Fig. 1
Fig. 1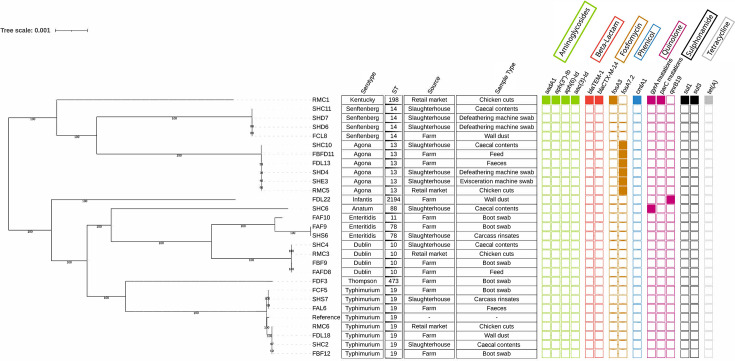 Fig. 2
Fig. 2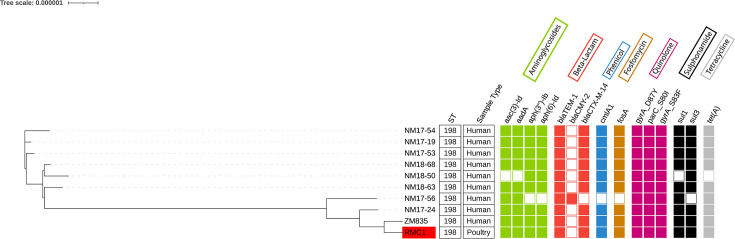 Fig. 3
Fig. 3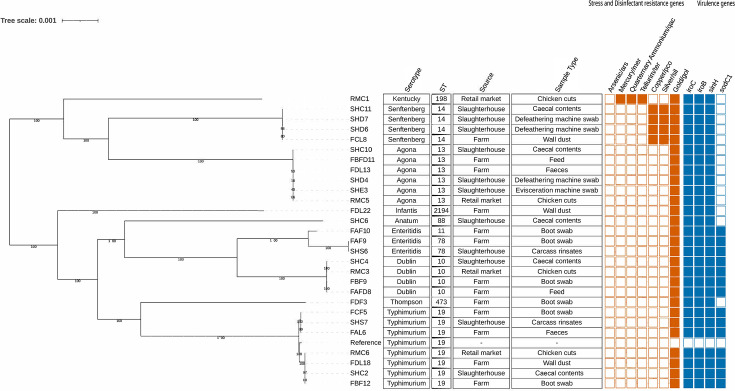 Fig. 4
Fig. 4- —http://dx.doi.org/10.13039/501100000276 Department of Health and Social Care
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSalmonella and Campylobacter epidemiology · Antibiotic Resistance in Bacteria · Listeria monocytogenes in Food Safety
Data Summary
Sequence data reported for the first time in this study have been deposited in the National Center for Biotechnology Information Sequence Read Archive (SRA) under the BioProject PRJNA1154600, and accession numbers are listed in supplementary data (Table S1, available in the online Supplementary Material). All other sequence data used in the analysis are in available databases accessible using accession numbers. One supplementary Excel file is available with the online version of this article.
Introduction
Salmonella is an important zoonotic foodborne pathogen that causes gastrointestinal infections in humans and has significant economic and public health impacts globally [1]. In the USA, Salmonella remains one of the most burdensome foodborne pathogens, with ~1.35 million cases of salmonellosis, 26,500 hospitalizations and 420 deaths reported each year [2]. Low- and middle-income countries experience the highest death rates, with Sub-Saharan Africa recording rates as high as 20–25% [3]. According to a review study by Ramtahal et al. [4], Salmonella was responsible for 19.7% of foodborne illnesses in South Africa. In Zimbabwe, Salmonella continues to be a significant foodborne pathogen and a leading cause of diarrhoeal diseases [56].
Poultry and poultry products are the principal sources of Salmonella and are potentially responsible for most foodborne zoonotic Salmonella transmission [7]. Salmonella can infect flocks in poultry farms and contaminate slaughter facilities through horizontal transmission via contaminated faeces, litter, feed, water, rodents, equipment and infected farm personnel [8]. Carcasses can be contaminated with Salmonella from the digestive tract during the defeathering and evisceration processes [9].
Despite numerous efforts to reduce Salmonella contamination in the food chain, the persistence of this pathogen in poultry products and related environments remains a global concern [10]. Compounding the problem is the worrying emergence of multidrug-resistant (MDR) Salmonella strains, which can spread to humans via the food pathway. Additionally, the pathogenic potential of Salmonella has been linked to the expression of various genes that promote host cell invasion, intracellular survival and stress tolerance in processing environments [11]. Sources and transmission routes of Salmonella along the food chain are not well understood, particularly in resource-limited settings due to the lack of integrated epidemiological surveillance systems. Gaining knowledge of such transmission dynamics is essential for developing effective Salmonella control programmes in the poultry value chain.
Diverse non-typhoidal Salmonella serovars, including those belonging to O-antigen groups O:4 (group B), O:7/O:8 (group C) and O:9 (group D), have been isolated from chicken farms in Zimbabwe [12]. An MDR S. Kentucky ST198 strain linked to both human clinical infections and poultry farms has also been reported [13]. However, Salmonella strains circulating within poultry production facilities have not been comprehensively investigated using a farm-to-fork approach. There is a significant knowledge gap regarding the resistance genes, virulence factors and genetic relatedness of isolates throughout the broiler supply chain. For the effective prevention of Salmonella, understanding the distribution of serovars along the broiler supply chain will provide valuable information regarding contamination routes and the possible persistence of serovars in the chain.
In this study, we examined the genomic diversity and phylogenetic relationships of Salmonella enterica strains circulating in poultry farm environments, slaughterhouses and retail chicken meat in Harare, Zimbabwe. Through whole-genome sequencing (WGS), we assessed the phylogenetic relatedness, antimicrobial resistance (AMR) profiles, virulence factors and stress tolerance of each isolate, aiming to enhance our understanding of transmission dynamics and the potential risks posed by Salmonella along the food supply chain.
Study design and sample collection
A longitudinal surveillance study was conducted over a 28-month period (August 2021–December 2023) to investigate the genetic diversity and AMR profiles of S. enterica circulating in the broiler supply chain in and around Harare, Zimbabwe. Twelve sites were included: four broiler farms (farms A–D), two commercial slaughterhouses (SH1 and SH2) and six retail markets (R1–R6), all located within a 40-km radius of Harare, a major hub for poultry production and distribution. All four farms were integrated into the commercial supply chain and routinely delivered birds to the two participating slaughterhouses.
A total of 552 samples were collected across the broiler supply chain, comprising farms (n=432), slaughterhouses (n=100) and retail markets (n=20). Farm samples included fresh faeces (n=80), feed (n=60), drinking water (n=72), wall dust (n=68), drinker and feeder swabs (n=72) and boot swabs (n=80). Slaughterhouse samples consisted of caecal contents (n=30), swabs from defeathering and evisceration machines (n=40) and post-chill carcass rinsates (n=30). Retail market samples consisted of chilled chicken cuts (n=20) supplied by the two participating slaughterhouses.
Isolation and identification
Bacterial isolation and identification were performed following the World Organisation for Animal Health Terrestrial Manual for isolation of Salmonella from food, feedstuffs, faecal and environmental samples [14]. Briefly, 10–25 g of samples were pre-enriched in 1 : 10 vol/vol Buffered Peptone Water (BPW) (Oxoid, Basingstoke, Hampshire, UK) and incubated for 16–20 h at 36±2 °C. Modified Semi-solid Rappaport-Vassiliadis (MSRV) agar plates (ISO) (Oxoid) and Müller–Kauffmann Tetrathionate broth (Oxoid) were inoculated with 0.1 ml and 1 ml of incubated BPW, respectively. The MSRV plates were incubated at 41.5±1 °C, while Tetrathionate broth was incubated at 36±2 °C. After 24 and 48 h of selective enrichment, 1 µl loop of material from the edge of the turbid growth zone on MSRV and 10 µl of Tetrathionate broth were streaked onto xylose lysine deoxycholate agar (XLD) (Oxoid) and Brilliance™ Salmonella Agar (Oxoid). The plates were incubated at 36±2 °C for 21–27 h. Plates were examined for presumptive Salmonella colonies, which appeared on XLD as opaque/yellow, pink or red colonies with or without black centres and on Brilliance Salmonella Agar as purple colonies. Presumptive Salmonella colonies were purified on nutrient agar (Oxoid), and species identification was confirmed by the API-20E biochemical test (API-20, BioMerieux, UK and Ireland) and slide agglutination with polyvalent ‘O’ (A-S) and poly ‘H’ (phase 1 and phase 2 antisera) (Remel). S. Typhimurium ATCC 14028 and Escherichia coli ATCC 25922 were used as positive and negative control strains, respectively.
WGS and analysis
The Salmonella isolates were sub-cultured on nutrient agar and incubated overnight at 37 °C. Genomic DNA was extracted using QIAamp DNA mini kits (Qiagen) as per the manufacturer’s instructions. DNA libraries were prepared using the Nextera XT (Illumina) sample preparation kit. The isolates were then sequenced on an Illumina NextSeq (San Diego, CA, USA) platform, generating 150 bp paired-end reads. FastQC v0.11.9 [15] and MultiQC v1.11 [16] were utilized to evaluate the quality of reads. PrinSEQ v0.20.4 [17] was employed to clean, filter and trim the raw reads, removing those with a mean quality score below 28 and trimming the first 10 bases from the left end of the reads. Draft genome assemblies were generated using SPAdes v3.14.1 at default settings [18]. The quality of these assemblies was evaluated with QUAST v5.0.2 [19]. The presence of genes and point mutations conferring AMR, virulence and heavy metal stress tolerance was assessed using ResFinder v4.5.0 [20] and AMRFinderPlus v3.12.8 [21]. The sequence type (ST) was determined with MLST v2.19.0 [22], using the pubMLST database [23]. In silico Salmonella serotyping was carried out using the SISTR command line tool [24]. Core genome SNPs were generated using SNIPPY [25], using S. enterica subsp. enterica serovar Typhimurium str. LT2 (accession: NC_003197) as a reference strain. Phylogenetic trees with 200 bootstraps were built using RAX-ML [26] from the core genome SNPs and annotated using iTOL v5 [27]. Sequence data is available in the National Center for Biotechnology Information Sequence Read Archive (SRA) under the BioProject PRJNA1154600.
SNP analysis was performed to compare S. enterica serovar Kentucky ST198 strains isolated from retail chicken cuts with human clinical strains obtained in Zimbabwe, specifically from Harare (n=6), Chitungwiza (n=1), Mutare (n=1) and Kadoma (n=1) [13]. The raw sequencing reads for the human clinical isolates are publicly available in the Sequence Read Archive (SRA) under study accession number PRJNA762287 (https://www.ncbi.nlm.nih.gov/sra).
Results
Prevalence and distribution of Salmonella in the broiler supply chain
The overall prevalence of Salmonella in the broiler supply chain was 5.1%(28/552). Retail markets had the highest contamination rate at 20.0% (4/20), followed by slaughterhouses at 11.0% (11/100) and farms at 3.0% (13/432) (Fig. 1).
Prevalence of Salmonella in broiler farms, slaughterhouses and retail markets.
All Salmonella isolates from the retail market originated exclusively from chicken cuts. In slaughterhouses, the highest Salmonella prevalence was detected in caecal contents at 16.7% [95% confidence interval (CI): 5.6–34.7%), followed by swabs from defeathering and evisceration machines at 10.0% (95% CI: 2.8–23.7%) and carcass rinsates at 6.7% (95% CI: 0.8–22.1%). On farms, Salmonella was detected in boot swabs at 7.5% (95% CI: 3.1–14.9), wall dust at 4.4% (95% CI: 0.9–12.4), faeces at 3.8% (95% CI: 0.8–10.6) and feed at 3.3% (95% CI: 0.4–11.5). However, Salmonella was not isolated from drinking water or swabs from drinkers and feeders.
Characterization and prevalence of S. enterica serovars in the broiler supply chain
All S. enterica isolates were identified as eight distinct serovars. The most prevalent serovar identified in silico was S. Typhimurium (n=7), all of which were ST19, followed by S. Agona (n=6), all ST13; S. Senftenberg (n=4), all ST14; S. Dublin (n=4), all ST10; and S. Enteritidis (n=3), with ST11 (n=1) and ST78 (n=2). S. Infantis (n=1) was identified as ST2194. The single strains of S. Anatum, S. Thompson and S. Kentucky were classified as ST88, ST473 and ST198 STs, respectively. Most STs were identified at multiple different sampling and production stages. Specifically, S. Typhimurium ST19 (isolates: FAL6, SHC2 and RMC6), S. Agona ST13 (isolates: FBFD11, SHC10 and RMC5) and S. Dublin ST10 (isolates: FAFD8, SHC4 and RMC3) were isolated from farms, slaughterhouses and retail markets. S. Senftenberg ST14 (isolates: FCL8 and SHC11) was isolated from both farms and slaughterhouses (Fig. 2).
Phylogenetic tree of Salmonella strains identified with AMR genes. Shaded boxes indicate the presence of the AMR gene, and unshaded boxes indicate the absence of the gene. S. Typhimurium LT2 was used as the reference strain as it was the most prevalent serovar in this study.
Analysis of AMR, virulence, stress tolerance and biocide resistance genes
A total of 15 AMR genes were detected across the broiler production chain, shared among 9 out of 28 Salmonella isolates (Fig. 2). The fosfomycin glutathione transferase gene (fosA7.2) was identified in six Salmonella Agona ST13 strains, isolated from different stages of the broiler production chain (farms and slaughterhouses). fosA7.2 is regarded as a potentially silent resistance gene [28]. While it may not confer detectable resistance under normal conditions, it can become clinically significant under selective pressure or through regulatory mutations and horizontal transfer. This is supported by the elevated fosfomycin resistance MIC values (>512 mg ml^−1^) reported in previous studies [29]. One S. Infantis strain (FDL22) carried the pentapeptide repeat protein gene (qnrB19), conferring resistance to fluoroquinolone antibiotics [13]. The only S. Anatum strain isolated along the production chain (caecal contents) possessed a gyrA mutation. Notably, antibiotic resistance genes qnrB19 and fosA7.2 appeared to be serotype-specific as qnrB19 was found only in S. Infantis and fosA7.2 was found only in S. Agona.
One isolate, the S. Kentucky ST198 strain (RMC1) recovered from a retail market, carried multiple AMR genes conferring resistance to aminoglycosides (aadA1, aph(3'')-Ib, aph(6)-Id and aac(3)-Id), fosfomycin (fosA3), folate pathway antagonists (sul1 and sul3), tetracycline (tetA) and amphenicol (cmlA1). Notably, this S. Kentucky ST198 strain also harboured genes conferring resistance to AmpC beta-lactamase (blaCMY-2), class A β-lactamase (blaTEM-1B) and extended spectrum β-lactamase (blaCTX-M-14), along with chromosomal point mutations in DNA gyrase (gyrA) and DNA topoisomerase (parC). This strain also shared similar antibiotic resistance profiles with S. Kentucky ST198 strains isolated from human clinical cases in Zimbabwe (Fig. 3).
Phylogenetic tree of S. Kentucky ST198 strains from human clinical cases and poultry from this study. Shaded boxes indicate the presence of the AMR gene, and unshaded boxes indicate the absence of the gene.
All isolates, except for one (FCL 16), harboured the virulence genes sinH, iroC and iroB, which enhance the pathogenicity of Salmonella by promoting host cell invasion and facilitating iron uptake [30] (Fig. 4). The sodC1 gene was found in all S. Enteritidis, S. Dublin and S. Typhimurium serovars. Various stress response genes were identified across different serovars. All isolates contained the gold operon (golS and golT). Interestingly, only S. Senftenberg serovars possessed both copper and arsenic stress genes. The S. Kentucky isolate carried additional mercury operon genes (merP, merR, merT, merD, merA, merE and merC), as well as genes conferring resistance to quaternary ammonium compounds (qacC/qacEDelta1) and tellurium stress response (terD, terW and terZ).
Phylogenetic tree of Salmonella strains identified in this study with virulence, stress response and biocide resistance genes. Shaded boxes indicate the presence of the resistance genes, and unshaded boxes indicate the absence of the gene. S. Typhimurium LT2 was used as the reference strain as it was the most prevalent serovar in this study.
Co-occurrence of antimicrobial, heavy metal and biocide resistance genes in Salmonella isolates from the broiler production chain
Of the 15 AMR genes, co-presence with heavy metal and biocide resistance genes was observed in 34.5% (10/29) of Salmonella isolates (Fig. 4). The most common co-presence was for gold and fosfomycin resistance (6/29, 20.7%), followed by gold, quinolone and fluoroquinolone resistance (2/29, 6.9%). Notably, a single S. Kentucky ST198 strain co-harboured genes for resistance to gold, copper, silver, arsenic, mercury and tellurite, along with resistance to aminoglycosides, beta-lactams, phosphonic, phenicol, quinolones, sulphonamides and tetracycline.
SNP analysis among Salmonella isolates from the broiler production chain
Pairwise SNP comparisons among different Salmonella STs revealed low SNP distances within certain strains (Table S2). Specifically, S. Agona ST13 and S. Senftenberg ST14 exhibited mean SNP distances of ~6 and 10, respectively, suggesting a recent common ancestor. Despite the genetic similarity, these isolates originated from different stages of the broiler production chain. For instance, the S. Agona ST13 strain comprised six isolates from various sources: farm (n=3), slaughterhouse (n=2) and retail market chicken (n=1). Within this group, the farm isolate FBFD11 differed by 5 SNPs from the slaughterhouse isolate SHE3 and the retail market isolate RMC5, which were obtained from an evisceration machine swab and chicken cuts, respectively. Similarly, the S. Senftenberg isolate SHD7 from a defeathering machine swab at a slaughterhouse differed by 6 SNPs from the farm isolate FCL8, which was sourced from wall dust. Although S. Typhimurium ST19 displayed a higher mean SNP distance of ~529 SNPs, significant genetic similarity was still observed within this ST. Notably, the farm isolate FBF12 from a boot swab was identical (0 SNPs) to the slaughterhouse isolate SHC2 from caecal contents and differed by only 1 SNP from the retail market isolate RMC6, which was obtained from chicken cuts.
SNP analysis among S. Kentucky ST198 strains from human clinical samples and retail chicken cuts from Zimbabwe
The S. Kentucky ST198 strain (RMC1) isolated from chicken cuts showed a mean distance of ~19 SNPs when compared to other S. Kentucky ST198 strains isolated from human clinical infections in Zimbabwe (Table S3). Notably, the human clinical isolates NM17-24 and ZM835 differed by only 6 and 10 SNPs, respectively, from the chicken isolate RMC1.
Discussion
Salmonella contamination in poultry and poultry products can occur at various stages of the food supply chain. An analysis of Salmonella prevalence across the broiler production chain in Zimbabwe revealed varying contamination rates at different stages. The overall prevalence of 5.1% was lower than the 10% prevalence reported in a previous study conducted in Zimbabwe by Makaya et al. [12]. These discrepancies may be due to differences in geographical locations, sample sizes, farm management practices, sampling procedures and the types of samples analysed [31]. Furthermore, the presence of Salmonella at various stages from farms to retail markets highlights the risk of contamination along the supply chain. Similar studies have shown that Salmonella strains frequently spread throughout the broiler chicken supply chain [32]. As noted by Waghamare et al. [33], this study confirms the higher prevalence of Salmonella spp. from processing facilities compared to production farms. While elevated Salmonella isolation rates in slaughter facilities are likely attributable to contamination within the processing environment, further compounded by persistent cleaning challenges and inadequate disinfection protocols [3435], positive samples were detected across multiple time points, from February 2023 to December 2023. These detections occurred intermittently and included isolates from both chicken cuts and slaughterhouse environments, indicating ongoing contamination rather than a single outbreak event. Notably, the highest frequency of Salmonella occurrence was recorded at the retail stage, which may reflect additional opportunities for cross-contamination during product handling, packaging and distribution [36]. Hygienic practices and cleanliness in these shops are often minimal, and they frequently receive potentially infected live birds from various broiler suppliers [37].
While US regulations allow for the use of antimicrobials such as peracetic acid in immersion chilling [38], this practice is not typical in Zimbabwe, where slaughterhouses rely solely on plain water chilling without chemical decontaminants. This lack of intervention, combined with variable hygienic standards and limited temperature control, likely facilitates Salmonella persistence on carcasses post-processing. At the farm level, biosecurity is also inconsistently applied, particularly in small- to medium-scale operations that dominate local production. Mixed farming systems are common, with poultry raised in proximity to cattle, goats and other livestock, often under minimal biosecurity. Such conditions create opportunities for cross-species transmission, as illustrated by the unusual detection of S. Dublin, a cattle-adapted serovar, in poultry samples. Environmental contamination through shared equipment, water sources or personnel movement is a plausible route for its introduction. Similarly, the occurrence of S. Agona, though not as rare as S. Dublin, is also atypical in poultry and may reflect environmental persistence rather than routine host association. These findings underscore the complex ecology of Salmonella in Zimbabwean poultry systems, where weak biosecurity, limited hygiene and absence of national control programmes contribute to the circulation of both typical and atypical serovars.
The predominance of S. Typhimurium and S. Enteritidis in this study aligns with previous reports from Zimbabwe [123940]. However, the detection of serovars such as S. Agona ST13, S. Senftenberg ST14, S. Dublin ST10, S. Infantis ST2194, S. Anatum ST88 and S. Thompson ST473 represents either a genuine shift in Salmonella diversity or an improvement in detection methods. Regardless, their presence in the poultry chain is noteworthy, as these serovars are linked to human foodborne illness across sub-Saharan Africa [4142].
WGS provided further insight into the relatedness of isolates across the production chain. The low SNP distances (<10 SNPs) among S. Agona ST13 and S. Senftenberg ST14 indicate that these strains belong to the same genetic cluster and likely share a recent common ancestor [43]. For example, the farm isolate FBFD11 showed only 5 SNP differences compared to isolates from the slaughterhouse (SHE3) and retail (RMC5), strongly suggesting clonal dissemination across the chain [44]. Likewise, S. Senftenberg ST14 isolates from farm wall dust (FCL8) and a slaughterhouse defeathering machine (SHD7) differed by only 6 SNPs, indicating persistence within production environments. Importantly, isolates within the same cluster shared identical gene profiles, further supporting common origins. For instance, the farm isolate FBF12, slaughterhouse isolate SHC2 and retail isolate RMC6 were genetically indistinguishable, demonstrating strain transmission from farm through slaughter to retail.
The analysis of virulence and stress response genes among Salmonella isolates from the poultry supply chain highlights significant adaptability and potential public health risks. In Zimbabwe, the prevalence of virulence genes in Salmonella from poultry was previously unknown, making this study the first to investigate Salmonella virulence in poultry isolates. The universal presence of virulence genes such as sinH, iroC and iroB underscores their critical role in Salmonella pathogenicity [45]. The iroB and iroC genes facilitate iron uptake from the host [46], while the sinH gene encodes proteins involved in cell adhesion and invasion [47]. The consistent detection of the sodC1 gene in key serovars highlights a common strategy for evading host defences [48]. These findings suggest that healthy poultry and their environment serve as reservoirs for pathogenic Salmonella strains that could be capable of causing infections in humans.
Stress response genes were widespread, with all isolates carrying the gold operon. The co-occurrence of AMR genes with stress response genes in all isolates is particularly alarming, as it suggests strong selective pressures that favour the emergence of multi-resistant strains. The presence of stress response genes in Salmonella poses significant public health risks by enhancing pathogen resilience, potentially leading to co-resistance with antibiotics, complicating treatment options and increasing environmental persistence and virulence [49].
The presence of an extended-spectrum beta-lactamase- and AmpC-producing S. Kentucky ST198 isolate in chicken cuts is particularly concerning. Though detected only once, this MDR strain has been associated with high-level fluoroquinolone resistance and widespread dissemination in Africa and Europe [5052]. The genetic similarity observed between poultry-derived S. Kentucky ST198 (RMC1) and human clinical isolates, with differences as low as 6–10 SNPs, suggests possible zoonotic transmission and a shared evolutionary lineage. This finding mirrors global reports identifying poultry as a reservoir for S. Kentucky ST198 in human infections [51].
Together, these results reinforce the interconnectedness of farms, processing plants and retail outlets as part of a continuous transmission network. The combination of weak biosecurity, lack of antimicrobial interventions, poor hygiene in slaughterhouses and informal retail practices creates an environment conducive to the persistence and spread of Salmonella, including MDR strains. Given these risks, enhanced biosecurity measures, stricter hygiene protocols in processing facilities and robust surveillance systems are essential to minimizing Salmonella transmission from poultry to humans [53]. While limited sample size and lack of longitudinal data prevent definitive conclusions, the evidence highlights a pressing need for improved biosecurity, enhanced surveillance and targeted interventions across Zimbabwe’s poultry production system. Framing these findings within a One Health perspective emphasizes that controlling Salmonella in poultry is essential not only for food safety but also for mitigating human health risks in Zimbabwe.
The genetic similarity between the isolates highlights the risk of cross-contamination or direct exposure to contaminated poultry products as potential transmission pathways for Salmonella Kentucky ST198. Given these risks, enhanced biosecurity measures, stricter hygiene protocols in processing facilities and robust surveillance systems are essential to minimizing Salmonella transmission from poultry to humans [53].
Conclusion
This study reveals the dynamic nature of Salmonella along the broiler supply chain in Zimbabwe, characterized by varying prevalence rates and diverse genetic profiles among isolates. The findings underscore the coexistence of AMR with heavy metal and biocide resistance, indicating potential co-selection pressures that contribute to the persistence of Salmonella. Understanding these dynamics is essential for developing effective strategies to mitigate Salmonella contamination and in poultry production systems. By addressing these issues through improved surveillance, responsible antimicrobial use and enhanced biosecurity measures, we can strengthen food safety, protect public health and promote sustainable poultry production systems.
Supplementary material
10.1099/mgen.0.001550Uncited Table S1.
10.1099/mgen.0.001550Uncited Table S2.
10.1099/mgen.0.001550Uncited Table S3.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chen H Qiu H Zhong H Cheng F Wu Z et al Non-typhoidal Salmonella infections among children in Fuzhou, Fujian, China: a 10-year retrospective review from 2012 to 2021 Infect Drug Resist 2023162737274910.2147/IDR.S 40815237180635 PMC 10171219 · doi ↗ · pubmed ↗
- 2Centers for Disease Control and Prevention (CDC) Salmonella 2021 https://www.cdc.gov/salmonella/index.htmlaccessed 20-January-2023
- 3Jeon HJ Disease Surveillance and Implications on Public Health Decisions: Example of Invasive Salmonellosis in Sub-Saharan Africa Apollo - University of Cambridge Repository 202310.17863/CAM.101820 · doi ↗
- 4Ramtahal MA Amoako DG Akebe ALK Somboro AM Bester LA et al A public health insight into Salmonella in poultry in Africa: a review of the past decade, 2010–2020 Microb Drug Resist 20222871073310.1089/mdr.2021.038435696336 · doi ↗ · pubmed ↗
- 5Mashe T Gudza-Mugabe M Tarupiwa A Munemo E Mtapuri-Zinyowera S et al Laboratory characterisation of Salmonella enterica serotype Typhi isolates from Zimbabwe, 2009-2017 BMC Infect Dis 20191948710.1186/s 12879-019-4114-031151421 PMC 6544939 · doi ↗ · pubmed ↗
- 6Paglietti B Falchi G Mason P Chitsatso O Nair S et al Diversity among human non-typhoidal Salmonella isolates from Zimbabwe Trans R Soc Trop Med Hyg 201310748749210.1093/trstmh/trt 04623843558 · doi ↗ · pubmed ↗
- 7Di Taranto P Petruzzi F Normanno G Pedarra C Occhiochiuso G et al Prevalence and Antimicrobial Resistance of Salmonella Strains Isolated from Chicken Samples in Southern Italy Microorganisms 20251327010.3390/microorganisms 1302027040005636 PMC 11858494 · doi ↗ · pubmed ↗
- 8Boubendir S Arsenault J Quessy S Thibodeau A Fravalo P et al Salmonella contamination of broiler chicken carcasses at critical steps of the slaughter process and in the environment of two slaughter plants: prevalence, genetic profiles, and association with the final carcass status J Food Prot 20218432133210.4315/JFP-20-25033513257 · doi ↗ · pubmed ↗