ROS-mediated interplay between brassinosteroids and gibberellic acids antagonistically modulates asymmetric periclinal cell division leading to middle cortex formation in Arabidopsis roots
Yoon Kim, Seung Hyun Nam, Soo-Hwan Kim

TL;DR
The study shows how brassinosteroids and gibberellins use reactive oxygen species to control asymmetric cell division in plant roots, shaping the middle cortex.
Contribution
ROS is identified as a central integrator of BR–GA antagonism in regulating asymmetric cell division during root development.
Findings
BRs and GAs antagonistically regulate middle cortex formation through ROS.
ROS-deficient mutants show reduced middle cortex formation and response to BRs or GAs.
BR–GA antagonism is linked to SHR/SCR-mediated asymmetric cell division in root development.
Abstract
Asymmetric cell divisions (ACDs) in the root ground tissue of Arabidopsis thaliana are essential for middle cortex (MC) formation, which contributes to root architecture and environmental adaptability. Here, we demonstrate that brassinosteroids (BRs) and gibberellins (GAs) antagonistically regulate MC formation via reactive oxygen species (ROS). Brassinolide (BL, a BR) or paclobutrazol (PAC, a GA biosynthesis inhibitor) promoted MC formation and sporadic periclinal cell divisions in root endodermal cell files, whereas brassinazole (BRZ, a BR biosynthesis inhibitor) or GA3 suppressed them. Consistently, the BR-signaling gain-of-function mutant bzr1-1D, the GA-biosynthesis-deficient mutant ga1-3, and the GA-insensitive mutant gai-1 exhibited elevated H2O2 levels and increased MC formation. Conversely, the BR-biosynthesis-deficient mutant det2 and the GA-signaling-enhanced rga/gai double…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
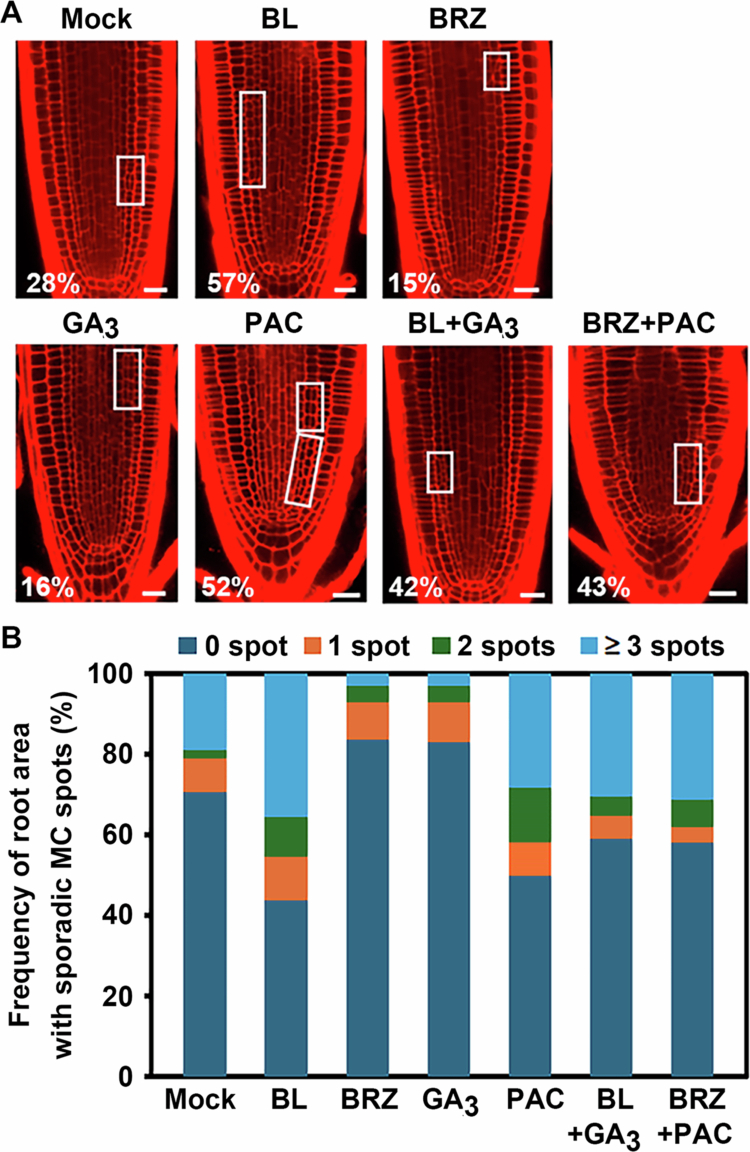 Figure 1
Figure 1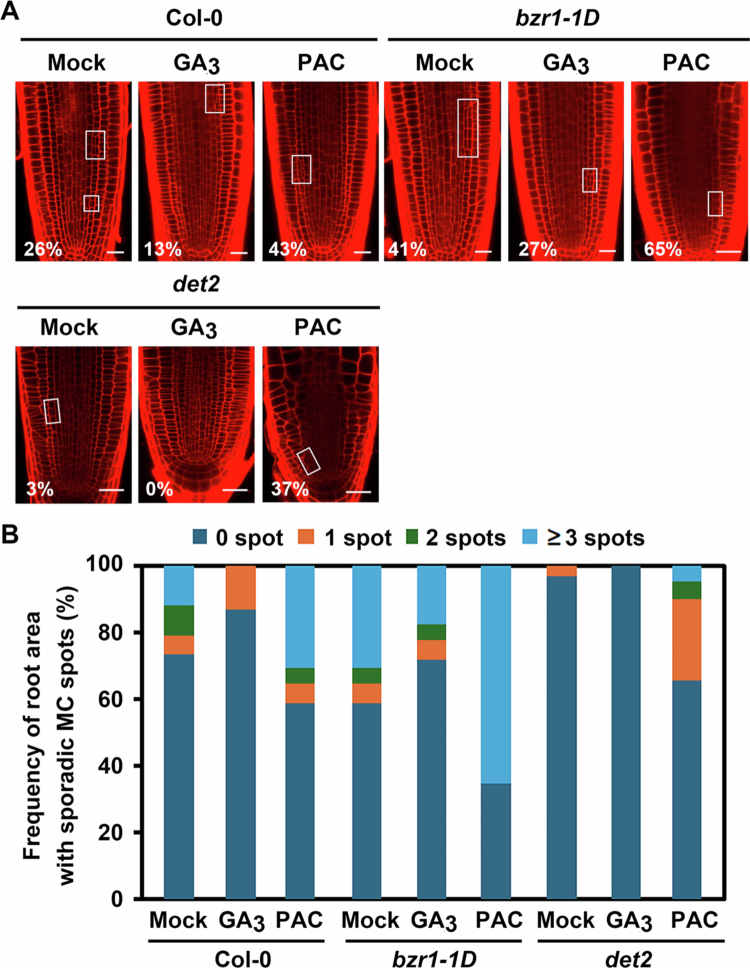 Figure 2
Figure 2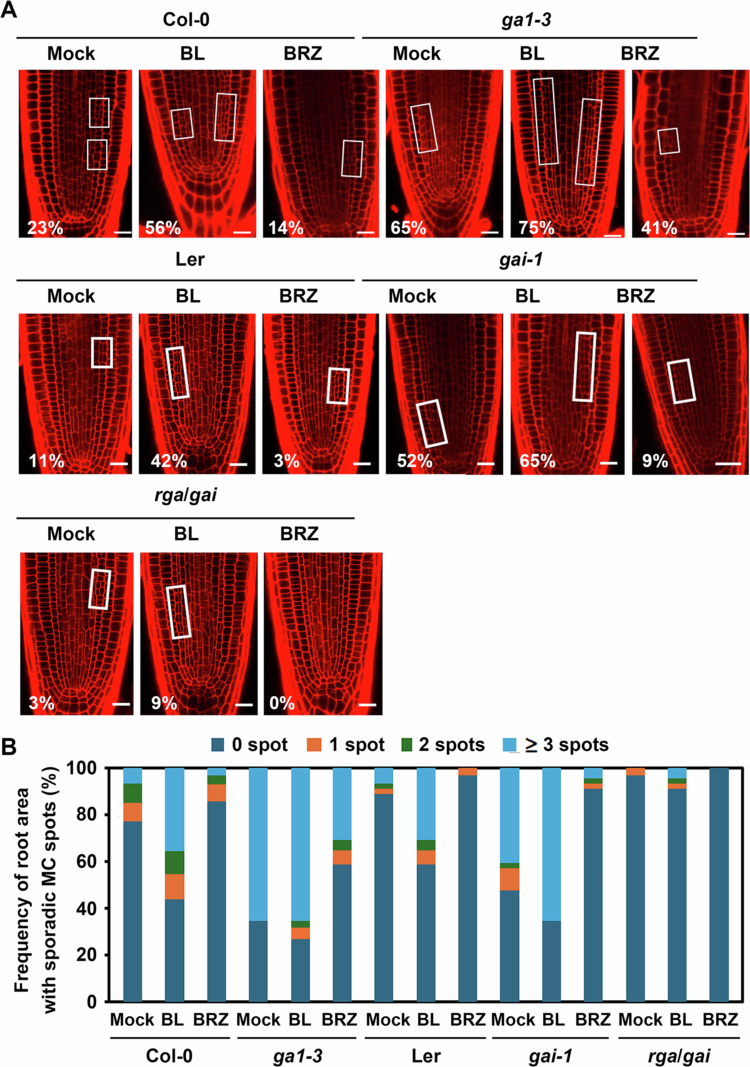 Figure 3
Figure 3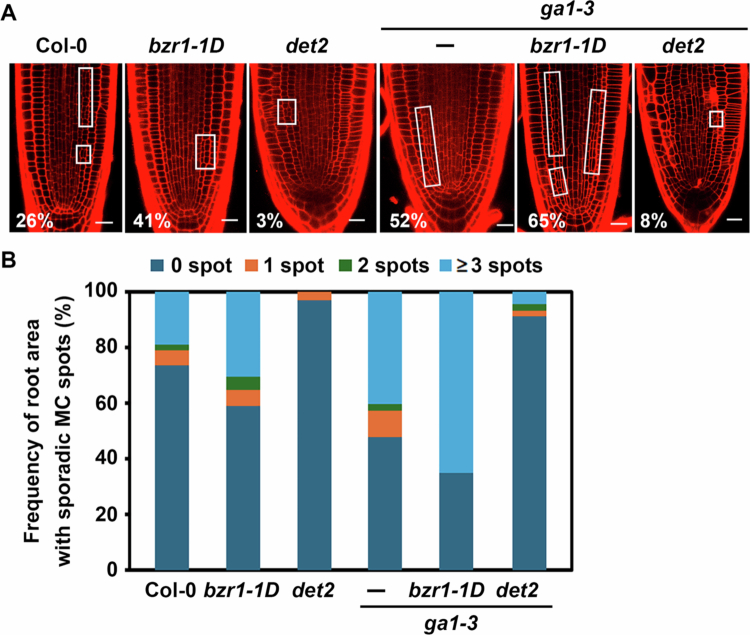 Figure 4
Figure 4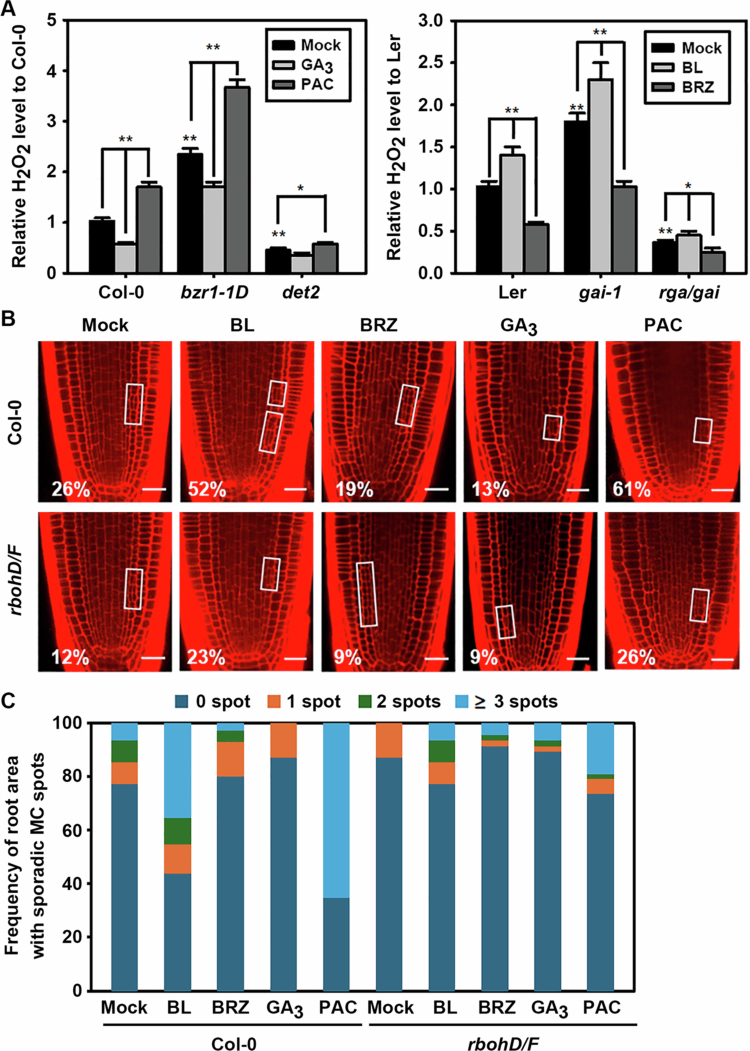 Figure 5
Figure 5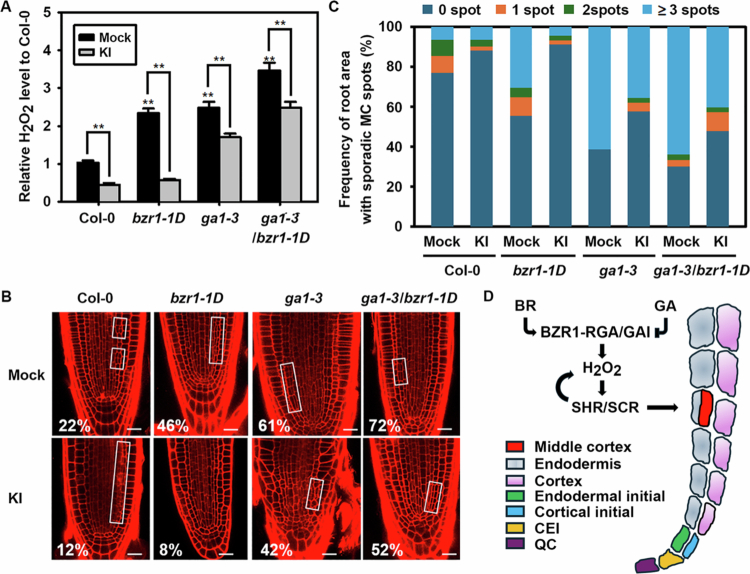 Figure 6
Figure 6- —National Research Foundation of Korea10.13039/501100003725
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · RNA Research and Splicing · Polysaccharides and Plant Cell Walls
Introduction
The root of Arabidopsis thaliana serves as a well–established model to investigate how asymmetric cell division (ACD) drives tissue patterning.1-3 One notable developmental event is the formation of the middle cortex (MC), a cell layer that arises from secondary asymmetric periclinal divisions in the endodermis during post–embryonic root development, typically occurring between 7 to 14 days after germination (DAG).4^,^5 As such, the onset of MC formation is commonly used as an indicator of root ground tissue maturation.
Accumulating evidence indicates that tight spatiotemporal regulation of ACDs during MC formation involves a complex interplay among plant hormones, redox signalling, and transcriptional regulators.4^,^6-13 Among the key regulators are the GRAS family transcription factors SHORT–ROOT (SHR), SCARECROW (SCR), and SCARECROW–LIKE 3 (SCL3), which coordinate ACDs of ground tissues producing cortex, endodermis, and MC.4^,^5^,^9^,^14-16 SHR protein, synthesised in the stele, moves to adjacent cell layers where it forms a nuclear complex with SCR and directly activates CYCLIND6;1 (CYCD6;1), promoting ACDs in the cortical–endodermal initials (CEI) and cortical–endodermal initial daughter (CEID) cells to produce the endodermis and cortex.16-19 SHR acts in a dose–dependent manner: high levels of SHR suppress MC formation, while intermediate levels promote it.16 SCR negatively regulates MC formation by repressing CYCD6;1 expression, as evidenced by precocious MC development in scr and scr3 mutants.5^,^9^,^20 Indeed, low threshold levels of SHR and SCR are sufficient and act early in the cell cycle to determine the orientation of the division plane, resulting in either formative or proliferative cell division in root stem cells.21
In parallel, redox signalling plays a crucial role in ACD regulation.22-25 SHR promotes production of hydrogen peroxide (H_2_O_2_) via transcriptional up–regulation of RESPIRATORY BURST OXIDASE HOMOLOGs (RBOHs), facilitating periclinal cell divisions.26 SCL3 contributes to ROS homoeostasis by modulating PEROXIDASE 34 (PRX34) transcription during tissue maturation.27
Among plant hormones, gibberellic acid (GA) functions as a molecular clock modulating the timing of asymmetric cell divisions in Arabidopsis root ground tissue.10 High GA levels suppress MC formation during early root development in a SHR-dependent process, whereas later reduction of GA biosynthesis permits MC formation.5^,^8^,^9^,^14^,^28 DELLA proteins, such as GIBBERELLIC ACID INSENSITIVE (GAI) and REPRESSOR OF ga1-3 (RGA), are negative regulators of GA signalling and play central roles in this process.29 In this regard, mutants defective in GA biosynthesis (ga1−3) or signalling (gai−1) exhibit increased endodermal ACDs and MC formation, highlighting GA’s inhibitory role.10 SCL3 modulates GA signalling and contributes to MC repression, and SCL3-DELLA interaction, in conjugation with the SHR–SCR module, plays an important role in the GA–mediated spatiotemporal control of MC formation.9^,^30
Brassinosteroids (BRs) are plant steroid hormones that play crucial roles in various aspects of plant growth and development.31 In roots, BRs regulate growth by modulating BRASSINAZOLE RESISTANT1 (BZR1)/BRI1 EMS SUPPRESSOR1 (BES1)-induced ethylene biosynthesis and by promoting peroxidase–dependent production of superoxide anions (O_2_^−^).32 Additionally, the BZR1-/BES1-dependent BR signalling pathway induces ectopic activation of quiescent centre cell division and regulates columella stem cell differentiation in a BR concentration–dependent manner.33-35 With regard to ground meristem differentiation, BR–activated BZR1 directly binds to the promoter of SHR to induce its expression and physically interact with SHR to enhance the transcription of RBOHs, thereby increasing H_2_O_2_ levels.12 This redox activation further enhances the SHR–mediated promotion of MC formation. However, it remains unclear how BRs coordinate with other hormonal pathways, particularly with GAs, in regulating MC formation. In this study, we investigate the regulatory crosstalk between BRs, GAs, and ROS during MC formation. We show that BRs and GAs oppositely modulate ROS production, resulting in antagonistic regulation of asymmetric endodermal cell divisions and MC development in Arabidopsis roots.
Materials and methods
Plant materials and growth conditions
Wild–type Arabidopsis thaliana Columbia−0 (Col−0) and Landsberg erecta (Ler), BRs– and GAs–related mutants (bzr1-1D, det2, ga1−3, ga1−3/det2, and ga1−3/bzr1-1D in the Col−0 background; gai−1 and rga/gai in the Ler background), and ROS–deficient rbohD/F mutant were used for phenotypic analysis of MC formation in this study. The rbohD/F seeds were kindly provided by Dr. Yuree Lee (Seoul National University, Republic of Korea).
For phenotypic analysis MC formation and H_2_O_2_ production, seedlings germinated on half–strength MS agar medium containing 0.8% phytoagar (pH 5.7) were transferred to fresh 1/2 MS medium supplemented with the chemicals indicated in each experiment. Seedlings at eight days after germination (DAG8) were analysed for MC formation and H_2_O_2_ production. Plants were grown in a growth chamber operating under a cycle of 16 h light and 8 h dark at 23−25°C with 70% humidity. All seedlings were grown vertically.
Chemicals and treatments
Brassinolide (BL), paclobutrazol (PAC), and potassium iodide (KI) were purchased from Duchefa Biochemie (Netherlands). Brassinazole (BRZ) was kindly provided by Dr. Yoshida (RIKEN, Japan). All other chemicals used in this study were purchased from Sigma–Aldrich (USA), unless otherwise indicated. For chemical treatments, DAG3 seedlings were transferred to 1/2 MS phytoagar plates supplemented with either BL (10^−12^ M), GA_3_ (10^−6^ M), PAC (10^−6^ M), or BRZ (10^−6^ M). For ROS scavenging assays, KI was added to the medium at a final concentration of 1 mM.
Confocal microscopy and imaging
For phenotypic analysis of MC formation, roots of DAG8 seedlings were stained with propidium iodide (PI, 10 μg/mL) for 1 min and mounted in water. PI signals were obtained using a Zeiss LSM 710 confocal laser–scanning microscope equipped with an argon ion laser (488 nm) and a He/Ne ion laser (543 nm). For observation of PI–stained roots, samples were excited at 543 nm and the PI emission signal was collected at 595−615 nm. MC formation (the frequency of endodermal ACDs) was quantified based on the presence of a clearly distinguishable extra cell layer between the endodermis and cortex. At least 100 roots were analysed for each experiment.
H2O2 assays
ROS levels in root tissues were measured using a commercially available Amplex® Red Hydrogen Peroxide Assay Kit (A22188, Invitrogen), according to the manufacturer’s instructions. In brief, roots of DAG8 seedlings grown on a half–strength MS medium in the presence or absence of the indicated chemicals were pulverised in liquid nitrogen. Following the addition of five–fold volumes of 50 mM sodium phosphate buffer (pH 7.4), the samples were thoroughly mixed and incubated on ice for 10 minutes. Subsequently, the mixtures were centrifuged at 12,000 rpm for 20 minutes at 4 °C. The resulting supernatants were collected and used for determination of H₂O₂ concentrations, as previously described.27^,^36 All experiments were performed at least triplicate, and the data were statistically analysed by Student’s t-test using Microsoft Excel (Microsoft, USA).
Results
GAs and BRs act antagonistically on asymmetric periclinal cell division, and their respective roles in modulating middle cortex formation are interdependent
Integrated regulation of periclinal cell division by the BZR1-SHR transcriptional module leads to MC formation in Arabidopsis roots, while GAs oppositely repress MC formation, suggesting complex regulatory crosstalk between BR and GA in MC development.5^,^8^,^12 Nonetheless, it remains unclear whether and how these hormones interact functionally in the context of MC regulation. To address this gap, we investigated the mutual influence of BRs and GAs on MC formation, focusing on whether these hormones act independently or interdependently through a shared regulatory mechanism.
First, we tested whether two hormones influence each other in MC formation. To do this, Col−0 seedlings were germinated and transferred to medium supplemented with BR– and/or GA–related compounds, such as BL (a BR), GA_3_ (an active GA), BRZ (a BR biosynthetic inhibitor) or PAC (a GA biosynthetic inhibitor). We found that BL or PAC treatment effectively promoted MC formation: 28% of non–treated Col−0 plants showed asymmetric periclinal cell division in endodermis, whereas 57% of BL–treated and 52% of PAC–treated plants exhibited endodermal cell division resulting in MC formation (Figure 1A). In contrast, BRZ or GA_3_ treatment led to an over 10% reduction in the proportion of MC–formed plants compared with mock–treated Col−0. Next, we investigated interactions between BRs and GAs by co–treating Col−0 seedlings with BL + GA_3_ or BRZ + PAC. Interestingly, BL + GA_3_ co–treatment showed an approximately 15% reduction in MC formation compared to BL treatment alone. A similar reduction was observed for BRZ + PAC compared to PAC alone (Figure 1A). Supporting this inhibitory interaction between BRs and GAs in MC formation, the frequency of MC spot areas with sporadic asymmetric cell divisions along the endodermal cell files showed a similar trend: BL and PAC treatments dramatically increased the MC spot area, whereas BRZ and GA_3_ treatments caused a notable decrease (Figure 1B).
Effects of BR– and GA–related chemicals on MC formation. (A) Representative confocal images of chemical–treated Col−0 roots. Open squares indicate root areas with MC–formed endodermal files. Numbers represent the percentage of plants showing MC formation in roots. Scale bar = 20μm. (B) Frequency of sporadic MC–formed root areas along the endodermal cell files for plants shown in (A). DAG8 seedlings were analysed for MC phenotypes. BL (10−12 M), GA3 (10−6 M), PAC (10−6 M), and BRZ (10−6 M). n > 100 seedlings for each experiment.
To further explore the functional interaction of BRs and GAs in MC formation, we examined the effects of GA_3_ or PAC on the BR–signalling gain–of–function mutant bzr1-1D (in which BZR1 is constitutively active) and the BR biosynthesis mutant det2 (which exhibits severe BR deficiency). Consistent with the previous report,12 bzr1-1D exhibited a ~15% increase in MC–formed roots compared to wild–type plants (Col−0), while det2 showed a prominent decrease (Figure 2A). Supplying GA_3_ to bzr1-1D seedlings negated the promotion effect on MC formation: the elevated rate in bzr1-1D (41%) was reduced to 27% by GA_3_ treatment, whereas PAC treatment of bzr1-1D increased the ratio of MC–formed plants to 65%, indicating that GAs can suppress BZR1-mediated, BRs–induced MC formation. Similarly, PAC treatment of det2 mutants increased the rate of MC–formed roots from 3% (mock–treated det2) to 37% (PAC–treated det2), suggesting that GA depletion can partially rescue the BR–deficient MC phenotype. Consistently, sporadic endodermal cell divisions were dramatically increased in plants with enhanced BR signalling (bzr1-1D), and were further increased in PAC–treated bzr1-1D or det2 plants (Figure 2B). In contrast, GA_3_ treatment on bzr1-1D plants efficiently decreased the ectopic MC formation promoted by bzr1-1D.
Antagonistic effects of GA–related chemicals on MC formation in BR–biosynthesis and –signalling mutants. (A) Representative confocal images of bzr1-1D and det2 roots treated with GA3 or PAC. Scale bar = 20μm. (B) Frequency of sporadic MC–formed root areas along the endodermal files for plants shown in (A). Open squares in (A) indicate root areas with MC–formed endodermal files. Numbers represent the percentage of plants showing MC formation in roots. n > 100 seedlings for each experiment.
Next, we examined the effects of BL and BRZ on the GA–deficient mutant ga1−3,37 the GA–signalling insensitive gai−1,38^,^39 and rga−24/gai–t6 (rga/gai in this report),40 which show constitutively active GA signalling due to null mutations both in RGA and GAI. As expected, the ga1-3 mutation in Col−0 plants exhibited an enhanced rate of MC formation compared to Col−0 wild type, and BL treatment of ga1-3 further increased the rate from 65% to 75%, while BRZ treatment partially suppressed it from 65% to 41% (Figure 3A). Similarly, gai−1 plants showed an increased rate of MC formation compared to Ler wild type, and BL treatment further increased the rate from 52% to 65%, while BRZ treatment efficiently suppressed it from 52% to 9%. In contrast, rga/gai plants, which exhibit constitutively active GA signalling, showed a reduced rate of MC formation (3%) compared to Ler wild type (11%). Moreover, BL treatment of these plants partially increased MC formation to 9%, confirming that GA activity negatively regulates the periclinal endodermal division, while BL acts antagonistically to the GA effect. The same trend of antagonistic regulation between BRs and GAs on ectopic MC formation was observed in BL– or BRZ–treated ga1-3, gai-1, and rga/gai: sporadic endodermal cell divisions were increased in GA–deficient ga1-3 or GA–insensitive gai-1, and BL treatment further increased the ectopic cell divisions and BRZ treatment produced the opposite effect (Figure 3B).
Antagonistic effects of BR–related chemicals on MC formation in GA–biosynthesis and –signalling mutants. (A) Representative confocal images of ga1−3, gai-1, and rga/gai roots treated with BL or BRZ. Scale bar = 20μm. (B) Frequency of sporadic MC–formed root areas along the endodermal cell files for plants shown in (A). Open squares in (A) indicate root areas with MC–formed endodermal files. Numbers represent the percentage of plants showing MC formation in roots. n > 100 seedlings for each experiment.
Results presented thus far suggest that BRs and GAs exert opposite effects on the asymmetric endodermal cell division and MC formation. To further assess functional interactions between these two phytohormones, we genetically crossed bzr1-1D or det2 with ga1−3 and examined their MC–related phenotypes. Consistent with observations in Figures 2 and 3, both bzr1-1D and ga1−3 exhibited enhanced MC formation/ectopic periclinal cell divisions along the endodermal files (Figure 4A, B). In contrast, MC formation was dramatically down–regulated in BR–biosynthesis–defective det2. We found that the bzr1-1D/ga1−3 double mutant exhibited an approximate 24% increase in MC–formed roots (65%) compared to bzr1-1D alone (41%) and a 13% increase compared to ga1−3 alone (52%). This indicates an additive or synergistic enhancement of MC formation due to combined BR signalling activation and GA deficiency. In contrast, crossing ga1-3 with BR–deficient det2 dramatically negated the MC–promoting effects of ga1−3, reducing MC formation from 52% (ga1-3) to 8% (ga1-3/det2). These results imply that BRs may share signalling components with GAs in modulating asymmetric endodermal cell divisions, highlighting the functional interplay between BR and GA in MC regulation.
Genetic confirmation of antagonistic regulation of MC formation by BR and GA. (A) Representative confocal images of BR–related, GA–related, and their double mutant roots. Open squares indicate root areas with MC–formed endodermal cell files. Numbers represent the percentage of plants showing MC formation in roots. Scale bar = 20μm. (B) Frequency of sporadic MC–formed root areas along the endodermal files for plants shown in (A). n > 100 seedlings for each experiment.
ROS functions as a key integrator of BR and GA signals in regulating their antagonistic periclinal cell division leading to middle cortex formation during ground tissue maturation
It was well documented that ROS and plant hormones, including BRs and GAs, intricately regulate root growth and development, serving as signalling molecules that govern processes such as cell proliferation and differentiation.23 BRs have been shown to increase H_2_O_2_ levels in roots while GAs decrease them,12^,^27 suggesting interplay between BR– and GA–mediated ROS homoeostasis and the resulting antagonistic regulation of MC development.
To validate this hypothesis, we comparatively analysed ROS accumulation in the roots of various BR– and GA–related mutants. First, ROS accumulation in roots, determined by measuring H_2_O_2_ levels, was markedly increased in BR–signalling–enhanced bzr1-1D and decreased in BR–biosynthesis–defective det2 compared to Col−0 (Figure 5A). Parallel to effects on MC development in bzr1-1D, GA_3_ treatment of Col−0 or bzr1-1D plants significantly down–regulated H_2_O_2_ production, while PAC treatment greatly up–regulated ROS production, implying that modulation of GA homoeostasis in roots alters BZR1- and BR–mediated MC development. On other hands, a semi–dominant GA–insensitive mutant gai-1 showed about a 1.8-fold increase in root H_2_O_2_ levels compared to Ler, while rga/gai plants, which show constitutively active GA signalling, exhibited a reduction of more than 60% in H_2_O_2_ production (Figure 5A). Conversely, supplying BL to Ler, gai-1, and rga/gai plants promoted H_2_O_2_ production, and BRZ treatment reduced ROS production. Collectively, the patterns of ROS accumulation observed in GA_3_- or PAC–treated bzr1-1D and det2, or in BL– or BRZ–treated gai-1 and rga/gai plants, closely mirrored the antagonistic regulation of MC development, suggesting a strong correlation between ROS levels and the BR– and GA–induced MC development.
Antagonistic effects of GA– and BR–related chemicals on H2O2 production and MC formation in BR– or GA–biosynthesis and signalling mutants. (A) Accumulation of H2O2 in roots of bzr1-1D and det2 treated with GA3 or PAC, and in roots of gai-1 and rga/gai treated with BL or BRZ. Bar graphs represent means ± SD relative to mock–treated Col−0 or Ler controls. SD: standard deviation. Statistical differences compared to the mock–treated controls or among bracketed samples are indicated by a single asterisks () for P < 0.05 and double asterisks (**) for P < 0.01. All experiments were performed at least triplicate, and statistical analyses were conducted using Student’s t-test in Microsoft Excel. (B, C) Effects of GA– and BR–related chemicals on MC formation in the ROS–deficient rbohD/F mutant. (B) Representative confocal images of chemical–treated rbohD/F roots. Open squares indicate root areas with MC–formed endodermal cell files. Numbers indicate the percentage of plants showing MC formation. Scale bar = 20 μm. (C) Frequency of sporadic MC–formed root areas along the endodermal files for plants shown in (B). n > 100 seedlings for each experiment.*
SHR induces accumulation of ROS in Arabidopsis roots by increasing the RBOHs, thereby elevating the levels of H_2_O_2.12^,^26 To establish a direct causal link between RBOHs–incurred ROS accumulation and BR– and GA–regulated MC development, we examined the effects of BL, BRZ, GA_3, and PAC on MC formation in the ROS–deficient rbohD/F mutant, which lacks RBOHD and RBOHF, two key NADPH oxidases. As reported, the rbohD/F mutant exhibited a significant reduction in the proportion of MC–formed roots compared to Col−0 wild type and showed a less frequent sporadic endodermal asymmetric cell divisions (Figure 5B, C). Furthermore, while BL or PAC treatment markedly facilitates MC formation in Col−0 plants (26% for BL–treated and 35% for PAC–treated), this stimulatory effect was substantially reduced in rbohD/F, which showed only 9% and 14% increases with BL and PAC treatment, respectively. These results strongly suggests that BRs and GAs antagonistically regulate asymmetric endodermal cell division primarily through their opposing effects on RBOHD– and RBOHF–mediated ROS accumulation in the root.
Next, we accessed the effects of potassium iodide (KI), a ROS scavenger, on ROS– and MC–related phenotypes of Col−0, bzr1-1D, ga1-3 and ga1-3/bzr1-1D double mutants to further validate ROS–mediated interplay of BR– and GA–induced MC formation. As shown in Figure 6, H_2_O_2_ production and the promotion of MC–forming endodermal asymmetric cell divisions were greatly enhanced in bzr1-1D, ga1−3 and ga1−3/bzr1-1D plants compared to Col−0 wild type, while supplying KI in the incubation medium greatly reduced these promotive effects, especially in bzr1-1D plants (Figure 6A−C). In fact, introduction of the ga1-3 mutation into bzr1-1D additively increased H_2_O_2_ production and MC formation in the ga1-3/bzr1-1D double mutant. These results again confirm that BR– or GA–induced regulation of asymmetric endodermal cell division is tightly correlated with H_2_O_2_ levels, and thus ROS may serve as a converging link for BR– and GA–induced antagonistic MC formation in Arabidopsis roots.
ROS production and MC formation in KI–treated mutants. (A) Accumulation of H2O2 in roots of bzr1-1D, ga1-3, and ga1-3/bzr1-1D mutants treated with KI, a ROS scavenger. Bar graphs represent means ± SD relative to mock–treated Col−0 control. SD: standard deviation. Statistical differences compared to mock–treated control or among bracketed samples are indicated by double asterisks (**) for P < 0.01. Experiments were performed in triplicate, and statistical analyses were conducted using Student’s t-test. (B, C) Effects of KI on MC formation in the mutants shown in (A). (B) Representative confocal images of KI–treated mutant roots. Open squares indicate root regions with MC–formed endodermal cell files. Numbers denote the percentage of plants exhibiting MC formation. Scale bar = 20 μm. (C) Frequency of sporadic MC–formed root areas along the endodermal files for plants shown in (B). n > 100 seedlings for each experiment. (D) A schematic model illustrating the ROS–mediated interplay between BR and GA signalling in MC formation. BZR1-activating BR signalling and RGA/GAI–repressing GA signalling oppositely modulate H2O2 production, ultimately leading to SHR/SCR–mediated antagonistic regulation of asymmetric cell division in the root endodermis. H2O2 enhances the interaction between BZR1 and SHR, and the resulting BZR1-SHR complex cooperatively promotes ROS production, forming a positive feedback loop that supports MC formation. Arrows represent promotion, while blunted arrows indicate repression.
Discussion
The apical region of Arabidopsis roots consists of a quiescent centre (QC), which is mitotically inactive, surrounded by various types of stem cell initials, including epidermal–lateral root cap initials, cortical–endodermal initials (CEI), stele initials, and columella initials.2^,^34 Asymmetric divisions of these initials give rise to distinct root tissues, ranging from the outermost lateral root cap and epidermis to inner ground tissue (cortex and endodermis), pericycle, and vascular tissues, resulting in the radial organisation of the root. The CEI undergoes anticlinal divisions to maintain itself and subsequently produces CEI daughter cells, which divide asymmetrically in a periclinal orientation to generate the inner endodermis and the outer cortex, termed MC.7^,^41 It is well established that the timing and extent of ACD–driven MC formation are tightly regulated by a network of plant hormones, redox signals, and transcription factors.3^,^7 In this study, we demonstrated that BRs and GAs exert opposing effects on MC formation in Arabidopsis roots, and that these effects converge on the regulation of ROS levels. Our results support a model in which ROS acts as a central integrative signal, mediating downstream transcriptional events that govern asymmetric cell divisions.
BRs and GAs coordinately regulate a wide range of plant growth and developmental processes, including seed germination, pathogen defence, and etiolation responses.42-46 Their signalling pathways are intricately linked: for example, DELLA proteins such as RGA and GAI inhibited BZR1-DNA binding ability, while GA–induced and HSP–mediated degradation of DELLAs enhanced BR–induced hypocotyl elongation.47^,^48 Consistently, bzr1-1D rescued the short hypocotyl phenotype of the GA–deficient ga1-3 mutant, while ectopic expression of RGA caused destabilization and inactivation of the BZR1.47^,^49 These reports collectively demonstrate that BZR1-mediated BR signalling in plant growth serves as a positive regulator of the GA pathway, and that RGA/GAI function as negative regulators of the BR pathway. Interestingly, our findings contrast this synergism in growth regulation by revealing antagonistic roles in MC formation. The GA–deficient mutant ga1-3 exhibited enhanced MC formation and elevated H_2_O_2_ levels, and an introduction of the bzr1-1D allele into this ga1−3 (ga1-3/bzr1-1D) resulted in an additive increase in ROS accumulation and MC formation, whereas ga1-3/det2 double mutants displayed suppressed MC development, suggesting that both BZR1 and RGA/GAI, in contrast to their roles in plant growth, function positively in BRs– and GAs–involved and ROS–mediated MC formation. These results imply that, during root development, BR and GA signalling components may act interdependently through protein–protein interactions, such as those between BZR1 and DELLA proteins,47^,^49 to regulate ROS homoeostasis and asymmetric division of endodermal cells.
Tissue– and development–specific differential functions of transcription factors (TFs) add multiple layers of regulatory complexity to plant development, highlighting their functional duality in switching between activation and repression modes depending on specific cellular and environmental conditions.50 For instance, in stomatal development, BZR1 inhibits meristemoid cell proliferation in cotyledons, whereas in the hypocotyl, it promotes the same developmental process.51^,^52 The activity of TFs has been reported to be modulated by redox signals, which intricately regulate transcriptional activity by modulating the interaction of redox–responsive TFs with their cis-elements, altering their interaction partners, or controlling their subcellular localisation between the cytoplasm and nucleus.53 Thus, it is plausible that the BZR1-DELLA complex may enhance BZR1’s transcriptional output by recruiting co–factors or altering chromatin accessibility during ROS–enriched MC formation.
Our findings that ROS accumulation correlates with asymmetric endodermal cell division and MC formation in various BR– and GA–related mutants strongly supports the role of ROS as a key integrative signal in this developmental process. This notion is further reinforced by the marked reduction in MC formation upon treatment with the ROS scavenger KI. Indeed, BR signalling–induced H_2_O_2_ production and the subsequent oxidation of BZR1 enhance its transcriptional activity by promoting its interaction with key regulators of the auxin and light signalling pathways, including AUXIN RESPONSE FACTOR 6 (ARF6) and PHYTOCHROME INTERACTING FACTOR 4 (PIF4), thereby modulating diverse BZR1-mediated processes in plant growth and development.54 In addition, H_2_O_2_ significantly increases the interaction between BZR1 and SHR, and the resulting BZR1/SHR complex cooperatively promotes CYCD6;1 expression to induce periclinal cell division in the root endodermis.12 Notably, the BZR1 and SHR/SCL3 module contribute to ROS homoeostasis through the transcriptional regulation of RBOHs and PRX34,12^,^26^,^27 forming a positive feedback loop that supports MC formation during ground tissue maturation in roots. We suggest that ROS functions not merely as a permissive factor but potentially as an instructive cue in activating the transcriptional programmes governed by SHR and SCR.12^,^55
In conclusion, we propose a model in which BR and GA signalling pathways converge on ROS biosynthesis to regulate SHR– and SCR–mediated asymmetric cell division during MC development (Figure 6D). This BR–GA–ROS–SHR/SCR axis provides a mechanistic framework for understanding how hormonal and redox cues are integrated to fine–tune developmental patterning in Arabidopsis roots. Further studies are needed to elucidate the direct molecular targets of ROS within the SHR/SCR network and to understand how the spatial and temporal regulation of BR and GA signalling contributes to developmental plasticity in the root ground tissue. In particular, studies integrating spatial transcriptomics, chromatin accessibility profiling**,** and protein interaction dynamics in a tissue–specific manner will be crucial for disentangle these complex regulatory networks.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Benfey PN, Scheres B. Root development. Curr Biol. 2000;10:R 813–R 815. doi: 10.1016/S 0960-9822(00)00814-9.11102819 · doi ↗ · pubmed ↗
- 2Bennett T, Scheres B. Root development-two meristems for the price of one? Curr Top Dev Biol. 2010;91:67–102. doi: 10.1016/S 0070-2153(10)91003-X.20705179 · doi ↗ · pubmed ↗
- 3Ramachandran P, Ramirez A, Dinneny JR. Rooting for survival: how plants tackle a challenging environment through a diversity of root forms and functions. Plant Physiol. 2025;197:Kiae 586. doi: 10.1093/plphys/kiae 586.PMC 1166357039657006 · doi ↗ · pubmed ↗
- 4Choi JW, Lim J. Control of asymmetric cell divisions during root ground tissuem maturation. Mol Cells. 2016;39:524–529. doi: 10.14348/molcells.2016.0105.27306644 PMC 4959016 · doi ↗ · pubmed ↗
- 5Paquette AJ, Benfey PN. Maturation of the ground tissue of the root is regulated by gibberellin and SCARECROW and requires SHORT-ROOT. Plant Physiol. 2005;138:636–640.15955927 10.1104/pp.104.058362 PMC 1150384 · doi ↗ · pubmed ↗
- 6Chang J, Hu J, Wu L, Chen W, Shen J, Qi X, Li J. Three RL Ks integrate SHR-SCR and gibberellins to regulate root ground tissue patterning in Arabidopsis thaliana. Curr Biol. 2024;34:5295–5306. doi: 10.1016/j.cub.2024.09.074.39476837 · doi ↗ · pubmed ↗
- 7Cui H. Middle cortex formation in the root: an emerging picture of integrated regulatory mechanisms. Mol Plant. 2016;9:771–773.27212386 10.1016/j.molp.2016.05.002 · doi ↗ · pubmed ↗
- 8Cui H, Benfey PN. Interplay between SCARECROW, GA and LIKE HETEROCHROMATIN PROTEIN 1 in ground tissue patterning in the Arabidopsis root. Plant J. 2009;58:1016–1027.19228333 10.1111/j.1365-313X.2009.03839.x PMC 2803106 · doi ↗ · pubmed ↗