Emergence of azithromycin and ciprofloxacin non-susceptible genotype 4.2.2 S. Typhi in Fiji
Ashwini Vinod, Andrew J. Hayes, Yogeshni Chandra, Sonika Kaajal Nair, Tashyam Pillay, Farieya Fayza Khan, Shalini Goundar, Luse Dulaki Kalou, Khushnuma Khan, Maria Borua Anise, Mary Valcanis, Xinwei Ruan, Shalini Singh, Jessica Barnden, Raphaël M. Zellweger, Alumita Vuakanisakea

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
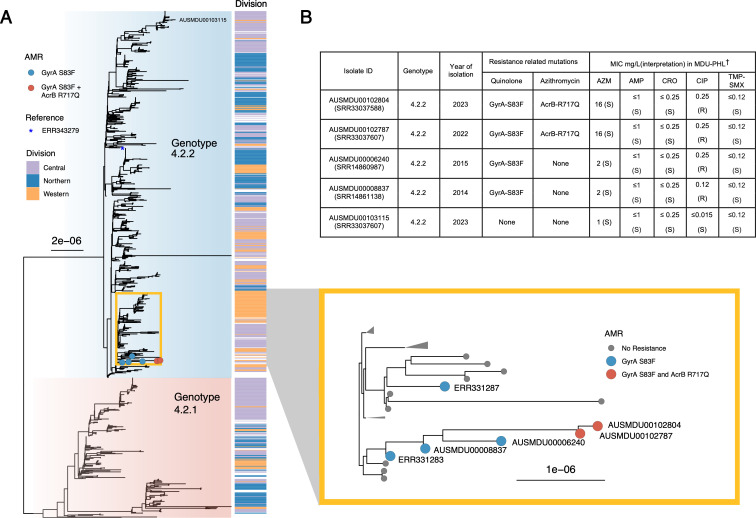 Fig 1
Fig 1- —University of Melbournehttp://dx.doi.org/10.13039/501100001782
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Salmonella and Campylobacter epidemiology · Escherichia coli research studies
LETTER
Typhoid is a systemic infection caused by the bacterium Salmonella enterica subspecies enterica serovar Typhi (S. Typhi) (1, 2). Typhoid is endemic in the island nation of Fiji in the South Pacific and mainly reported in indigenous Fijians and males, with a peak age of incidence between 15 and 29 years (3–5). S. Typhi isolates in Fiji are associated with a single evolutionary-related endemic genotype (4.2) comprised of two lineages—4.2.1 and 4.2.2, and antimicrobial resistance (AMR) is uncommon (5–9). In this study, we conduct a retrospective genomic epidemiology study of S. Typhi in Fiji and describe the presence of extant S. Typhi isolates with multiple chromosomal mutations (GyrA S83F and AcrB efflux pump R717Q) altering susceptibility to ciprofloxacin and azithromycin. These resistance findings have not previously been reported in the genotype 4.2 and were not associated with international strain importation events.
Our primary study site was the Western Division of Fiji with 337,041 inhabitants accounting for 38.1% of the total population of Fiji (10). The Lautoka hospital is the main tertiary hospital in the division and is a referral center for six subdivisional hospitals (Rakiraki, Tavua, Ba, Lautoka/Yasawa, Nadi, and Nadroga/Navosa), 28 health centers and 24 nursing stations (11). The study included S. Typhi isolates cultured between January 2020 and December 2023. All laboratory procedures (culture and antimicrobial susceptibility testing) were performed in Lautoka hospital. Minimum inhibitory concentration (MIC) for ciprofloxacin was not performed prior to June 2023. Whole genome sequencing and additional broth microdilution phenotypic testing was performed at Microbiological Diagnostic Unit Public Health Laboratory in Melbourne, Australia. Chromosomal markers of antimicrobial resistance were predicted using the Typhi module (12) within Mykrobe (13). Details on the bioinformatics methods are provided in the Supplemental material.
We sequenced 197 isolates from 176 patients, representing 38% of the total during the study period. Almost all isolates (99.5%) belonged to the genotype 4.2 with subclade 4.2.2 majority (84.3%). Two genetically related subclade 4.2.2 isolates (1 SNP difference) carried chromosomal point mutations in the quinolone resistance determining region of DNA gyrase at codon 83 (GyrA-S83F), a marker of ciprofloxacin resistance; and the AcrB efflux pump at codon 717 (AcrB-R717Q), a marker associated with resistance to azithromycin (14) (Fig. 1). Review of 781 historical Fijian isolates from Central and Northern divisions between 1981 and 2019 identified four additional isolates with the GyrA-S83F mutation and three likely evolved from an undefined single common ancestor (Fig. 1A). Four of these isolates were tested phenotypically and all exhibited resistance (MIC = 0.25 mg/L) to ciprofloxacin (Fig. 1B). While the two isolates with secondary AcrB-R717Q mutation had an MIC of 16 mg/L, they were higher compared with isolates without AcrB mutation (MIC = 1-2 mg/L). These two isolates in the current study were cultured from blood samples collected in December 2022 and February 2023 from Nadroga/Navosa and Ba subdivisions, respectively. Additional information on epidemiological linkage and treatment outcome is not available.
Population structure and phenotypic antimicrobial resistance profile of S. Typhi isolates in Fiji. (A) Maximum likelihood tree built from 578 parsimony informative SNPs of 978 genotype 4.2 S. Typhi isolates relative to the genotype 4.2.2 reference genome ERL072973 (position indicated by an asterisk). Shading refers to S. Typhi genotype as assigned by genotyphi. The associated health division where the isolate was isolated is displayed by the division column. The inset (yellow box) relates to the position of antimicrobial-resistant (AMR) isolates containing chromosomal point mutations in GyrA S83F (blue dots) and GyrA S83F plus AcrB R717Q (red dots). (B) Phenotypic antimicrobial susceptibility results of clinical S. Typhi isolates with GyrA (S83F) mutation conferring elevated MIC to ciprofloxacin (interpreted using EUCAST breakpoints) and secondary AcrB (R717Q) mutation conferring elevated MIC to azithromycin relative to a representative wild-type isolate (AUSMDU00103115/SRR33037607). †Broth microdilution using the Sensititre system in CMV5AGNF plate. MIC testing was done in December 2024 at the Microbiological Diagnostic Unit Public Health Laboratory. AZM, azithromycin; AMP, ampicillin; CRO, ceftriaxone; TMP-SMX, trimethoprim-sulfamethoxazole; MIC, minimum inhibitory concentration; MDU-PHL, Microbiological Diagnostic Unit Public Health Laboratory.
Azithromycin resistance in S. Typhi is uncommon (15). The first case was identified in Bangladesh among the MDR subclade of 4.3.1 in 2013 (14). More cases were subsequently reported in other countries with AcrB-R717Q/L mutations arising independently with no apparent epidemiological link (16–20). Similarly, the Fijian isolates with point mutations are genomically distinct and occurred in a non-MDR genotype. International importation is unlikely, as the 4.2 genotype is an archetypical S. Typhi genotype of Fiji (6, 7) and associated travelers from Fiji (8).
To conclude, we identified the emergence of MDR S. Typhi strains in the endemic 4.2 genotype. We recommend enhanced phenotypic surveillance with introduction of routine MIC testing for ciprofloxacin (21) and ongoing genomic surveillance to monitor the evolution of the current strains and detection of newly emerging AMR. Further investigation is warranted to demonstrate the correlation between AcrB-R717Q/L mutations, phenotypic azithromycin resistance, including MIC breakpoints and clinical outcomes. The use of typhoid conjugate vaccination (TCV) in the Northern Division has reduced disease burden (22); expansion of TCV will strengthen nationwide typhoid control.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Parry CM, Hien TT, Dougan G, White NJ, Farrar JJ. 2002. Typhoid fever. N Engl J Med 347:1770–1782. doi:10.1056/NEJ Mra 02020112456854 · doi ↗ · pubmed ↗
- 2Crump JA, Sjölund-Karlsson M, Gordon MA, Parry CM. 2015. Epidemiology, clinical presentation, laboratory diagnosis, antimicrobial resistance, and antimicrobial management of invasive Salmonella infections. Clin Microbiol Rev 28:901–937. doi:10.1128/CMR.00002-1526180063 PMC 4503790 · doi ↗ · pubmed ↗
- 3Scobie HM, Nilles E, Kama M, Kool JL, Mintz E, Wannemuehler KA, Hyde TB, Dawainavesi A, Singh S, Korovou S, Jenkins K, Date K. 2010. Impact of a targeted typhoid vaccination campaign following cyclone Tomas, Republic of Fiji, 2010. The American Society of Tropical Medicine and Hygiene 90:1031–1038. doi:10.4269/ajtmh.13-0728 PMC 404772524710618 · doi ↗ · pubmed ↗
- 4Thompson CN, Kama M, Acharya S, Bera U, Clemens J, Crump JA, Dawainavesi A, Dougan G, Edmunds WJ, Fox K, Jenkins K, Khan MI, Koroivueta J, Levine MM, Martin LB, Nilles E, Pitzer VE, Singh S, Raiwalu RV, Baker S, Mulholland K. 2014. Typhoid fever in Fiji: a reversible plague? Trop Med Int Health 19:1284–1292. doi:10.1111/tmi.1236725066005 PMC 4285329 · doi ↗ · pubmed ↗
- 5Getahun Strobel A, Parry CM, Crump JA, Rosa V, Jenney A, Naidu R, Mulholland K, Strugnell RA. 2019. A retrospective study of patients with blood culture-confirmed typhoid fever in Fiji during 2014-2015: epidemiology, clinical features, treatment and outcome. Trans R Soc Trop Med Hyg 113:764–770. doi:10.1093/trstmh/trz 07531638153 PMC 6907003 · doi ↗ · pubmed ↗
- 6Davies MR, Duchene S, Valcanis M, Jenkins AP, Jenney A, Rosa V, Hayes AJ, Strobel AG, Mc Intyre L, Lacey JA, Klemm EJ, Wong VK, Sahukhan A, Thomson H, Page A, Hocking D, Wang N, Tudravu L, Rafai E, Dougan G, Howden BP, Crump JA, Mulholland K, Strugnell RA. 2022. Genomic epidemiology of Salmonella Typhi in Central Division, Fiji, 2012 to 2016. Lancet Reg Health West Pac 24:100488. doi:10.1016/j.lanwpc.2022.10048835769175 PMC 9234096 · doi ↗ · pubmed ↗
- 7Getahun Strobel A, Hayes AJ, Wirth W, Mua M, Saumalua T, Cabenatabua O, Soqo V, Rosa V, Wang N, Lacey JA, Hocking D, Valcanis M, Jenney A, Howden BP, Duchene S, Mulholland K, Strugnell RA, Davies MR. 2024. Genetic heterogeneity in the Salmonella Typhi Vi capsule locus: a population genomic study from Fiji. Microb Genom 10:001288. doi:10.1099/mgen.0.00128839254668 PMC 11385387 · doi ↗ · pubmed ↗
- 8Wong VK, Baker S, Connor TR, Pickard D, Page AJ, Dave J, Murphy N, Holliman R, Sefton A, Millar M, Dyson ZA, Dougan G, Holt KE, International Typhoid Consortium. 2016. An extended genotyping framework for Salmonella enterica serovar Typhi, the cause of human typhoid. Nat Commun 7:12827. doi:10.1038/ncomms 1282727703135 PMC 5059462 · doi ↗ · pubmed ↗