Increase of macrolide resistance among Streptococcus pyogenes pharyngitis driven by a mef(A)-msr(D)/emm2-ST55 lineage in Portugal (2014–2019)
Ana Friães, Rafael Mamede, Beatriz Santos, Gina Marrão, José Melo-Cristino, Mario Ramirez

TL;DR
This study tracks the rise of macrolide-resistant Streptococcus pyogenes in Portugal, showing how a specific lineage became dominant in 2019.
Contribution
Identifies a new mef(A)-msr(D)/emm2-ST55 lineage driving increased macrolide resistance in pharyngitis cases in Portugal.
Findings
Macrolide resistance peaked at 9% in 2019 due to the rapid spread of mef(A)-msr(D)-positive emm2-ST55 isolates.
Resistant isolates showed lower genetic diversity and minimal overlap with susceptible lineages.
Tetracycline resistance was commonly linked to tet(M) and tet(O) genes co-located with erm genes.
Abstract
Increases in macrolide resistance occurred recently among Streptococcus pyogenes (Group A Streptococcus, GAS) in some countries. While the importance of monitoring the clinical and molecular epidemiology of non-invasive GAS is increasingly recognized, most surveillance focuses on invasive infections, since culture is rarely performed in tonsillo-pharyngitis. We determined the antimicrobial susceptibility and characterized the macrolide-resistant lineages of 2,002 pharyngeal isolates recovered in a Portuguese hospital in 2014–2019. There were seasonal variations in the numbers of recovered isolates, with peaks shifting between March–July and October–December, but consistently low numbers in August and September. Macrolide-resistant and macrolide-susceptible GAS presented independent seasonal and clonal dynamics, with resistant isolates showing lower genetic diversity and minimal overlap…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
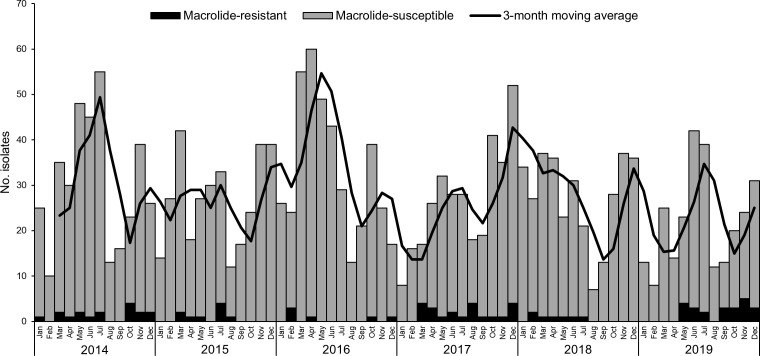 Fig 1
Fig 1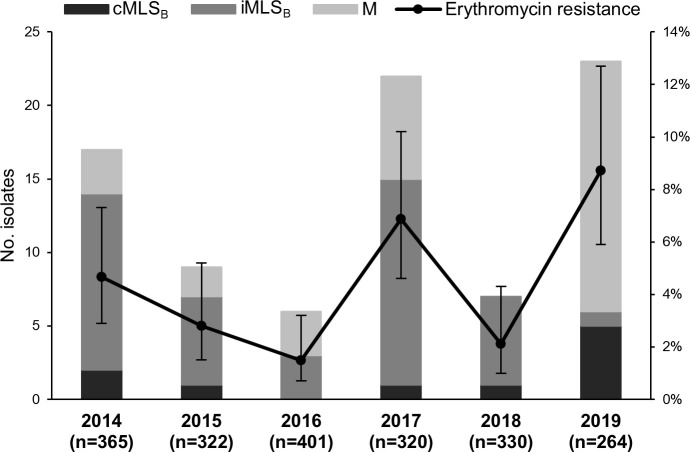 Fig 2
Fig 2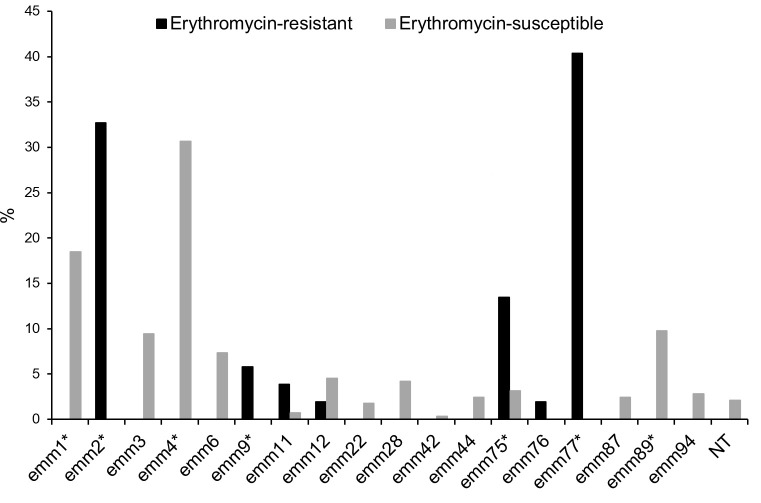 Fig 3
Fig 3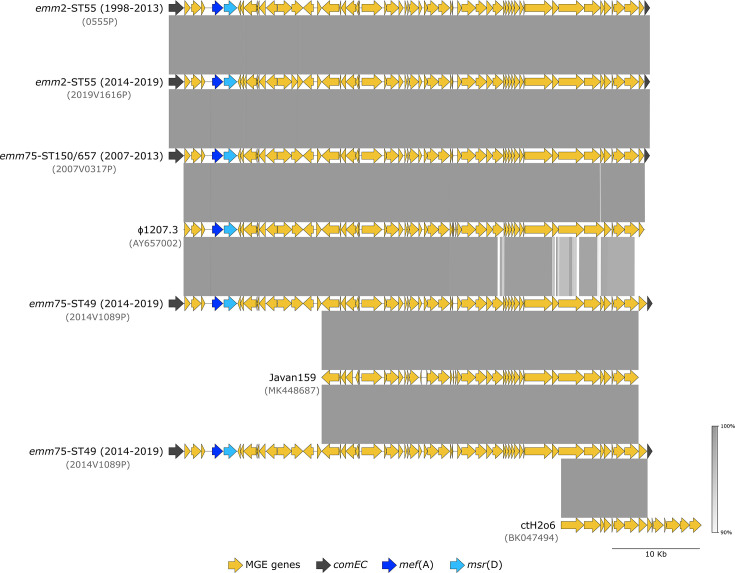 Fig 4
Fig 4| ST | Resistance phenotype | Resistance genes | No. isolates | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | Total | ||||
| 63 | iMLSB, Tet | 12 | 6 | 3 | 14 | 5 | 1 | 41 | ||
| 399 | iMLSB, Tet | 1 | 1 | |||||||
| 55 | M | 17 | 17 | |||||||
| 49 | M | 3 | 2 | 3 | 7 | 15 | ||||
| 150 | cMLSB, Tet | 1 | 1 | 2 | ||||||
| 75 | cMLSB, Tet | 3 | 3 | |||||||
| 403 | cMLSB, Tet | 2 | 2 | |||||||
| cMLSB | 1 | 1 | ||||||||
| 36 | cMLSB, Tet | 1 | 1 | |||||||
| 50 | cMLSB, Tet | 1 | 1 | |||||||
| MGE | Resistance genes | Insertion site | Genetic lineage |
|---|---|---|---|
| ɸ1207.3 |
| ||
| ɸ1207.3-like |
| ||
| ICE |
| ||
| Tn | DQL26_RS02190 | ||
| Tn |
| ||
| Tn |
| ||
| Tn | acetate CoA-transferase α | ||
| Tn |
| ||
| ICE-HKU372-like |
| ||
| pRW35 | - |
- —Fundação para a Ciência e a Tecnologiahttp://dx.doi.org/10.13039/501100001871
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsStreptococcal Infections and Treatments · Otolaryngology and Infectious Diseases · Infective Endocarditis Diagnosis and Management
INTRODUCTION
Streptococcus pyogenes (Lancefield Group A Streptococcus, GAS) remains uniformly susceptible to penicillin and other β-lactams, but macrolides and lincosamides offer alternatives for β-lactam-allergic patients and the association of clindamycin with a β-lactam is recommended for the treatment of severe invasive GAS infections (1). However, macrolide and lincosamide resistance among GAS presents marked geographical and temporal variations that have been associated with both antibiotic consumption and clonal dynamics (1, 2). Recently, an increase in macrolide resistance among invasive GAS was reported in the United States (3), and the World Health Organization included macrolide-resistant GAS in its Bacterial Priority Pathogens List in 2024 (4).
Macrolide resistance in GAS is mostly associated with the presence of the erm family of genes or the mef(A/E)-msr(D) operon. The erm genes confer cross-resistance to macrolides, lincosamides, and streptogramin B, either constitutive (cMLS_B_ phenotype) or induced by a macrolide (iMLS_B_ phenotype), due to rRNA methylation (1, 2). Carriage of mef(A/E)-msr(D) results in resistance to macrolides alone (M phenotype) by a mechanism likely involving ribosomal protection in addition to active efflux (5, 6). The erm and mef(A/E)-msr(D) genes are carried on a variety of mobile genetic elements (MGEs), which allow the horizontal dissemination of macrolide resistance while frequently carrying other resistance determinants (2).
Macrolide resistance among pharyngeal isolates recovered throughout Portugal declined from 28% in 2000 to 1% in 2013 (7). This decrease was not associated with marked changes in macrolide consumption but was instead accompanied by multiple clonal changes (7–9). Since 2013, national guidelines discourage the use of culture for the etiological diagnosis of tonsillopharyngitis in favor of rapid antigen detection tests (10). Several European countries follow similar practices hampering the monitoring of antimicrobial resistance among pharyngeal GAS. Although macrolide resistance among invasive GAS in Portugal remained below 5% throughout the 2010s (11), we had previously found a significantly higher macrolide resistance among pharyngeal isolates than contemporaneous invasive isolates, highlighting the potential of the pharynx as a reservoir of macrolide-resistant GAS (12).
To gain a more comprehensive view of the clonal landscape of macrolide-resistant GAS after 2013, we determined the antimicrobial susceptibility and characterized genomically a collection of pharyngeal GAS isolates prospectively collected in a Portuguese hospital center in 2014–2019.
MATERIALS AND METHODS
Bacterial strains
Between January 2014 and December 2019, pharyngeal swabs were obtained and cultured from suspected cases of streptococcal tonsillo-pharyngitis at Unidade Local de Saúde da Região de Leiria, Leiria, Portugal (ULSRL). ULSRL includes three tertiary hospitals covering an estimated population of 400,000 inhabitants in the center region of Portugal. GAS isolates were identified by colony morphology, β-hemolysis, and detection of the Lancefield group A antigen. The Institutional Review Board of Centro Académico de Medicina de Lisboa approved the study (258/22–AD/24). The study was exempt from obtaining written informed consent from the patients because it used only anonymized demographic data and involved samples collected as part of routine diagnostic procedures, with no additional intervention or identifiable information.
Antimicrobial susceptibility testing
Susceptibility testing was performed by disk diffusion on Mueller-Hinton agar supplemented with 5% defibrinated sheep blood, according to the guidelines and interpretative criteria of the Clinical and Laboratory Standards Institute (CLSI) (13), using the following disks (Oxoid, Basingstoke, UK): penicillin, erythromycin, clindamycin, tetracycline, and levofloxacin. The D-zone test was used to detect inducible clindamycin resistance (13).
Whole genome sequencing
A total of 391 pharyngeal GAS isolates were subjected to whole genome sequencing (WGS) as previously described (14), including all erythromycin-resistant isolates recovered at ULSRL during 2014–2019 (n = 84), one-third of the erythromycin-susceptible isolates recovered at ULSRL during 2017–2019, selected randomly (n = 287), and erythromycin-resistant isolates recovered throughout Portugal during 1998–2013, previously characterized (7–9) and belonging to emm2, emm75, and emm77 (n = 20) (Table S1). De novo assembly and determination of emm types, seven-gene multilocus sequence types (STs), core-genome multilocus sequence typing (cgMLST) profiles, and antimicrobial resistance genes were performed as described elsewhere (14, 15). The genomic analyses also included 215 genetically, temporally, geographically, and clinically diverse isolates of emm2, emm11, emm12, emm75, and emm77 selected from the Davies et al. data set (16) according to previously detailed criteria (14), as well as eight emm77-ST63 isolates from Poland (17) (Table S2). MGE identification relied on BLAST searches and alignments with public sequences of known MGEs carrying macrolide resistance determinants, using ICEfinder (18) to identify putative integrative conjugative elements (ICEs). Detailed methods for WGS and data analysis are available as supplemental material.
Statistical analysis
Isolate diversity was evaluated using the Simpson’s Index of Diversity (SID) with corresponding 95% confidence interval (CI_95%_) (19). Two-tailed Fisher’s exact test and odds ratios were used to identify significant pairwise associations. The Cochran-Armitage test was used to evaluate trends. The P-values for multiple tests were corrected with the false discovery rate (FDR) linear procedure (20). Values of P < 0.05 were considered significant.
RESULTS
Isolate distribution and demographic data
A total of 2,002 non-duplicate GAS isolates were cultured from pharyngeal swabs of patients with tonsillo-pharyngitis at ULSRL in 2014–2019 with an average of 334 isolates per year (range 264–401). Only four patients were adults (≥18 years old), and the median age was 6 years (interquartile range 4–9 years). Fifty-two percent of the patients were female. Age and sex were unknown for one patient.
The isolates were not evenly distributed throughout the year, and differences were seen between years (Fig. 1). The number of isolates was consistently low during August–September, and in most years, January and/or February also presented low isolate counts. In general, there were two peaks each year, one during Spring/early Summer (varying between March and July) and the other in Autumn, between October and December. The distribution of macrolide-resistant isolates was different from that of macrolide-susceptible isolates, with no clear seasonal pattern.
Monthly distribution of 2,002 pharyngeal GAS isolated between 1 January 2014 and 31 December 2019. The black line represents the 3-month moving average of the total isolates.
Antimicrobial resistance
All isolates were susceptible to penicillin and levofloxacin. Erythromycin presented the highest resistance rate (n = 84, 4.2%), followed by tetracycline (n = 77, 3.8%, of which 51 isolates were also erythromycin-resistant), and clindamycin (n = 52, 2.6%).
Erythromycin resistance remained mostly <5%, decreasing between 2014 and 2016 (P = 0.01). However, there were two peaks, one of 6.9% in 2017 and another of 8.7% in 2019 (Fig. 2). Up to 2018, the dominant macrolide resistance phenotype was iMLS_B_. After a complete suppression in 2018, the M phenotype emerged as the dominant phenotype in 2019, accompanied by an increase in cMLS_B_, while iMLS_B_ was found in only one isolate.
Yearly distribution of erythromycin resistance and associated phenotypes among 2,002 pharyngeal GAS isolated between 1 January 2014 and 31 December 2019. Error bars represent 95% CIs. cMLSB, constitutive resistance to macrolides, lincosamides, and streptogramin B; iMLSB, inducible resistance to macrolides, lincosamides, and streptogramin B; M, resistance to macrolides and susceptibility to lincosamides and streptogramin B.
Comparison between macrolide-resistant and macrolide-susceptible isolates
In order to compare the clonal composition of macrolide-resistant and susceptible GAS causing pharyngitis, one-third of the susceptible isolates from each year between 2017 and 2019 were randomly selected for WGS. During this period, resistant isolates (n = 52) presented a significantly lower diversity of emm types (seven emm types, SID = 0.720 [CI_95%_ 0.644–0.797]) and STs (8 STs, SID = 0.735 [CI_95%_ 0.660–0.811]) than susceptible isolates (n = 287; 14 emm types, SID = 0.837 [CI_95%_ 0.810–0.864]); 25 STs, SID = 0.860 [CI_95%_ 0.832–0.887]) (P < 0.01). Furthermore, the emm type distribution differed markedly between the two subgroups, with four emm types overrepresented among macrolide-resistant isolates (emm2, emm9, emm75, and emm77), and three emm types associated with the macrolide-susceptible subset (emm1, emm4, and emm89) (Fig. 3). Only emm11, emm12, and emm75 co-occurred among macrolide-susceptible and macrolide-resistant isolates.
*emm type prevalence among erythromycin-resistant (n = 52) and macrolide-susceptible (n = 287) pharyngeal GAS isolated between 1 January 2017 and 31 December 2019. NT, non-typable due to emm gene deletions or fusions; P < 0.05, significant after FDR correction.
Genetic lineages associated with macrolide resistance
The most common genetic lineage among macrolide-resistant isolates, identified in all study years, was an emm77-ST63 lineage (n = 41, 49%), carrying erm(A) and tet(O) and presenting the iMLS_B_ phenotype and tetracycline resistance (Table 1). The minimum-spanning tree (MST) of the cgMLST profiles of these isolates was constructed together with those of other emm77 isolates, including pharyngeal macrolide-resistant isolates recovered previously throughout Portugal (n = 9) (7), isolates from Poland (n = 8) (17), and genetically diverse isolates from multiple countries (n = 41) (16) (Tables S1 and S2). The MST shows four main genetic lineages and one singleton separated by >1,000 allelic differences (from a total of 1,314 compared loci) (Fig. S1). All emm77-ST63 isolates from Portugal were clustered in the same lineage, together with isolates from multiple continents presenting either ST63 or its single-locus variant (SLV), ST1023. Regardless of isolation date and hospital, isolates from Portugal were closely related, with most MST links ranging between 0 and 11 allelic differences (the exceptions being two isolates from 2007 to 2013 linking at 23 allelic differences). Isolates from the United States (n = 2) and the United Kingdom (n = 2) carrying erm(A), as well as one isolate from Australia with no detected macrolide resistance genes, were also closely grouped to isolates recovered in Portugal during 2007–2013 (MST links of 14–24 allelic differences). The single isolate in our collection harboring erm(T) also belonged to emm77 but presented an unrelated ST (ST399) and grouped into a distinct lineage, close (five allelic differences) to an ST399 isolate from the United States that also carried erm(T).
The emm2-ST55 lineage, exhibiting the M phenotype and carrying the mef(A)-msr(D) gene combination, comprised 17 isolates (20%). An MST was constructed with these isolates and all emm2 isolates in our collection of pharyngeal isolates recovered during 1998–2013 throughout Portugal (n = 5) (7, 9), as well as with the emm2 isolates present in the diverse collection from Davies et al. (16) (Fig. S2). Despite having been recovered in six countries from four continents throughout >20 years, the emm2 isolates presented a very limited genetic diversity, all belonging to ST55 and with MST links ranging from 0 to 59 allelic differences in a total of 1,427 core loci. The isolates recovered in the present study formed a subcluster with distances of 0–3 allelic differences, in agreement with their close geographic and temporal clustering (all recovered in 2019). Remarkably, isolates from Portugal were the only isolates carrying macrolide resistance genes. However, despite sharing the same resistance genes (mef(A)-msr(D)), isolates from 1998 to 2013 were more closely related to macrolide-susceptible isolates from several other countries than to the 2019 macrolide-resistant isolates from ULSRL.
The emm75-ST49 lineage carrying mef(A)-msr(D) accounted for all the M phenotype isolates between 2014 and 2017 (n = 15, 18%) but completely disappeared in 2018 and 2019 after peaking in 2017 (7/22 isolates, 32%). Using cgMLST data, all emm75 isolates from the present study (including 17 macrolide-resistant and nine macrolide-susceptible isolates) were compared with pharyngeal, macrolide-resistant emm75 isolates recovered previously throughout Portugal (n = 6) (7), and with the emm75 assemblies from the Davies et al. data set (n = 40) (16). The respective MST reflects a high genetic diversity, presenting three main genetic lineages and a singleton separated by >1,000 allelic differences (in a total of 1,305 core loci), and multiple links > 100 allelic differences (Fig. S3). All emm75-ST49 isolates carrying mef(A)-msr(D) from our study were closely clustered (MST links of 0–5 allelic differences) and were genetically related (21 allelic differences) to a strain from Canada carrying no macrolide resistance genes. All isolates from the analyzed data set presenting ST49 or its SLV ST861 were grouped in the same sublineage, regardless of carrying the mef(A)-msr(D) genes or not. The remaining isolates from Portugal were all grouped in a distinct sublineage separated by >489 allelic differences, including the pharyngeal resistant isolates from the national study (2007–2013). The latter, despite sharing the same resistance genes (mef(A)-msr(D)), presented distinct STs, namely ST150 (double locus variant [DLV] of ST49) or ST657 (SLV of ST150 and triple locus variant [TLV] of ST49). These isolates formed a cluster linked by 43 allelic differences to a strain from Belgium with no macrolide resistance genes. In 2014 and 2015, there were two cMLS_B_ emm75-ST150 isolates, also tetracycline-resistant, and carrying the erm(B) and the tet(M) genes. These were the only two isolates carrying the erm(B) gene in the MST and were both grouped close to the macrolide-susceptible isolates recovered in our study during 2017–2019, all sharing ST150.
The MST of emm11-ST403 isolates from this study (three macrolide-resistant and two macrolide-susceptible isolates) and the emm11 isolates from the Davies et al. data set (n = 51) (16) grouped all emm11-ST403 isolates into the same sublineage, regardless of geographic origin and of carrying macrolide-resistance genes (Fig. S4).
For emm12, we constructed an MST with the isolates from this study (one macrolide-resistant and 13 macrolide-susceptible isolates) together with diverse emm12 isolates from Davies et al. (n = 69) (Fig. S5). Isolates carrying different macrolide resistance-conferring genes and susceptible isolates were interspersed throughout the MST, regardless of geographic origin.
For the remaining lineages associated with macrolide resistance in our collection (emm9-ST75 and emm76-ST50), no genomic comparisons were performed, since they were each represented by few isolates (n ≤ 3) and were not found among the sequenced macrolide-susceptible isolates.
Mobile genetic elements encoding macrolide resistance
In M phenotype isolates representing emm2-ST55 (from both the 1998–2013 and 2014–2019 periods) and emm75-ST150/657 (from 2007 to 2013), the mef(A)-msr(D) genes were found in elements presenting ≥99.9% nucleotide identity over the entire length of the ɸ1207.3 reference sequence (accession no. AY657002), and inserted into the comEC gene as previously described for ɸ1207.3 (21, 22) (Table 2; Fig. 4). In emm75-ST49 isolates, the mef(A)-msr(D)-carrying element inserted into comEC was different. It presented 99.9% identity with ɸ1207.3 and other related elements, such as ɸ29862, ɸ29961, and ɸ29854 (accession nos. MT303952, MT311968, and MT311967) (23) but only over the 5′ 35.2 kb sequence. It showed lower identity with any of these elements in the 3′ region (93.8%–93.9% pairwise identity over 17.5 kb). Most of this region was 99.99% identical at the nucleotide level, with phage Javan159 (accession no. MK448687) identified in the draft genome of a Streptococcus dysgalactiae subsp. equisimilis strain (accession no. ERR111982) (24). However, this prophage lacks the 5′ 15.6 kb region of ɸ1207.3 carrying mef(A)-msr(D), as well as the 3′-terminal open reading frame (ORF) annotated as encoding a hypothetical protein in the element from our emm75-ST49 isolates. This ORF was found in the partial genome of a phage from an unknown host (Caudoviricetes sp. isolate ctH2o6, accession no. BK047494) identified in a human sample by metagenomics (25). The 3′-terminal 9.8 kb region of the ɸ1207.3-like element in our emm75-ST49 isolates shares 99.97% nucleotide identity with ctH2o6 (Fig. 4). This suggests that the emm75-ST49 ɸ1207.3 variant may have emerged through recombination between ɸ1207.3 (or a related element) and other phages.
Comparison of the elements carrying mef(A)-msr(D) genes in emm2-ST55, emm75-ST150/657, and emm75-ST49 isolates from this study, the reference sequence of ɸ1207.3, phage Javan159 from S. dysgalactiae subsp. equisimilis (24), and the partial sequence of a phage identified by metagenomics in a human sample (ctH2o6) (25). The names of the representative isolates used for each lineage or the GenBank accession numbers of the sequences are indicated in gray. The gray-shaded areas connect regions based on nucleotide identity (90%–100%). Annotated coding sequences are represented as arrows: genes belonging to the MGE in orange, except for mef(A) (dark blue) and msr(D) (light blue), and the interrupted chromosomal comEC gene into which the element is inserted in dark gray.
The erm(A) gene was found exclusively in emm77-ST63 isolates (from this study and from the national study of 2007–2013), together with tet(O) in an element sharing 99.78% nucleotide identity with the ICESp2905 reference sequence (accession no. FR691055) and 99.98% with the ICESp2905 from emm77-ST63 strain 1851/03 from Poland (accession no. NZ_CP155740), inserted into the rumA gene (17, 21) (Fig. S6).
The erm(B) gene was associated with multiple elements. In emm9-ST75, emm12-ST36, emm75-ST150, and emm76-ST50 isolates, erm(B) and tet(M) were found in Tn3872-like elements inserted into larger MGEs identified as putative ICEs in ICEfinder (Table 2; Fig. S7). The element in emm9-ST75 isolates appears to be the same element present in Houston isolate TSPY153 (accession no. CP060639), but with a deletion of 1,221 bp resulting in a chimeric coding sequence (CDS). The element is inserted immediately upstream of a hypothetical protein CDS (locus tag DQL26_RS02190) annotated in the complete genome of an emm9-ST75 isolate lacking the Tn3872 element (NCTC8316, accession no. NZ_LS483521). In emm75-ST150 isolates, the Tn3872-carrying element was inserted into the hsdM gene, similarly to isolates of the same lineage recovered in Spain (21). The emm76-ST50 isolate presents a Tn3872-like element with a transposase inserted downstream of tet(M), carried in a longer element, which is inserted into the hsdM gene. The element found in the emm12-ST36 isolate shares 99.99% nucleotide identity with the element carrying erm(B) and tet(M) in the complete genome of an emm12-ST36 strain from Italy (Isolate 26, accession no. CP167006) (26), although it is inserted into a different gene (acetate CoA-transferase α subunit in our isolate and phosphatidylglycerophosphatase A in CP167006). The Tn3872-like element in the emm12-ST36 isolates shares the lowest nucleotide identity with the reference sequence of Tn3872 of Streptococcus agalactiae (accession no. OP715845) (92.5%) among the Tn3872-like elements identified in this study (Fig. S7).
Besides erm(B) and tet(M), the emm11-ST403 isolates from 2019 (n = 2) carry a truncated aadE gene, as well as the aph(3')-IIIa and sat4 genes encoding aminoglycoside and streptothricin modifying enzymes, suggestive of a Tn6003-like element (27). Although the complete element was not assembled into a single contig in our isolates, read mapping supported the presence of Tn6003-like elements similar to those found in the complete genome of an emm11-ST403 strain from Italy (Isolate 18, accession no. CP167011) (26). In contrast, the emm11-ST403 isolate from 2014 carried only the erm(B) gene in an element similar to ICE-HKU372 (28). As in the emm12 isolate from Hong Kong where this element was originally identified (accession no. ERR060322), the ICE-HKU372-like element in our isolate is inserted into the hsdM gene but has a 1,428 bp sequence encoding two hypothetical proteins inserted into the gene encoding a Cro/CI family transcriptional regulator (locus ERR060322p_11600) (Fig. S8).
Finally, the erm(T) gene was found in a single isolate (emm77-ST399), on a 5,044 bp contig sharing 99.9% nucleotide identity with plasmid pRW35 (accession no. EU192194). As reported in emm77-ST399 isolates from the United States (29), this isolate also carried the tet(M) gene located in a different genetic element.
DISCUSSION
GAS pharyngitis is generally considered to peak during winter and early spring, favored by increased indoor time and crowding (30, 31). Our data support a consistent decrease in the number of cases during mid- and late-summer (August-September), but infection peaks were more variable, frequently occurring in late spring/early summer, while the coldest months of January and February usually presented lower numbers of isolates. This aligns with a study from the Spanish island of Mallorca, which reported a rise in cases starting in March, peaking in June, and followed by a summer slump (32) potentially highlighting different dynamics in temperate climates.
Our results are consistent with macrolide-resistant GAS having their own dynamics, distinct from that of macrolide-susceptible isolates (33). This was reflected in the absence of a seasonal pattern in the distribution of resistant isolates (Fig. 1), as well as in the distinct clonal profiles of the resistant and susceptible subsets during 2017–2019. Resistant isolates presented a lower genetic diversity as measured by ST and emm type, with only three emm types common to both subsets (emm11, emm12, and emm75) and several overrepresented in either susceptible or resistant isolates. Fluctuations in resistance rates were underpinned by specific clones underlining the importance of clonal dynamics in macrolide resistance in streptococci (34). There were also yearly variations in the prevalence of some of the susceptible clones in 2017–2019 (Fig. S7). Notably, although emm12 was always present throughout this period, macrolide-susceptible and -resistant sublineages did not co-exist in the same year.
In periods overlapping with this study, similarly low erythromycin resistance rates (<10%) were reported in Spain (4%–10%, invasive isolates, 2016–2019), Ireland (5%, invasive and non-invasive isolates, 2012–2015), Houston (4%, pharyngeal isolates, 2013–2017), and Canada (8.5%–10%, invasive isolates, 2016–2019) (35–38). However, resistance rates >10% were recorded in Bulgaria (31%, invasive and non-invasive isolates, 2013–2016), Southern Hungary (11%, invasive and non-invasive isolates, 2008–2017), and Central Greece (16%, pharyngeal isolates, 2011–2017) (39–41). These illustrate important geographic variations even within the European region. Notable increases in macrolide resistance were observed in Barcelona, Spain, where resistance increased from 6% in 2013–2015 to 12% in 2016–2018 among invasive and non-invasive infections in adults (21), and in the United States, where resistance among invasive isolates increased from 18% in 2015–2017 to 25% in 2018–2019 mainly due to the expansion of emm92 isolates carrying erm(T) (3).
In line with pharyngeal isolates collected nationwide during 2007–2013 and with invasive isolates throughout the 2010s (7, 11), overall macrolide resistance among the isolates recovered at ULSRL during 2014–2019 was low (4%), presenting a decreasing trend between 2014 and 2016. In 2017, there was a resistance peak of 7% associated with an increase in lineages that were already circulating in previous years, namely emm77-ST63 carrying erm(A) and tet(O) and emm75-ST49 harboring mef(A)-msr(D). These two lineages sharply decreased (emm77-ST63) or completely disappeared (emm75-ST49) in 2018–2019, while an emm2-ST55 clone carrying mef(A)-msr(D) surged in 2019, leading to a macrolide resistance peak of 9% and to an inversion in the dominant resistance phenotypes. Similar changes in resistance phenotypes driven by clonal replacements have been previously observed at the national level in GAS (9). Throughout this study, tetracycline resistance was equally low (4%), and 66% of the tetracycline-resistant isolates were also resistant to erythromycin and clindamycin, reflecting the frequent co-occurrence of tet and erm genes in the same MGEs.
WGS data were used to identify the MGEs carrying macrolide resistance genes, as well as to compare the major genetic lineages found in this study with those previously identified nationwide, and to contextualize them among the international landscape of GAS clones. The dominant emm77-ST63 lineage in this study carried an ICESp2509 element and was already circulating nationally in 2007–2013 (7). Isolates of emm77-ST63 closely related to ours (Fig. S1) or carrying the same MGE have been identified in other countries, such as the United States, United Kingdom, Poland, and Spain (16, 17, 21). Furthermore, emm77 was among the top 10 emm types presenting macrolide resistance worldwide during 2000–2020 (42).
In contrast to emm77, the emm75 isolates carrying mef(A)-msr(D) in this study represent a distinct genetic sublineage from those recovered during 2007–2013, which was reflected by their separate clustering in the MST (Fig. S3), but also on their different STs (ST49 in the current study and ST150/657 in 2007-2013). Isolates of emm75 are known to be genetically diverse and multiple lineages carrying macrolide resistance determinants in different MGEs are geographically spread (16, 21, 42). The emm75-ST49 sublineage had not been previously identified in Portugal but was found in the United States, Canada, and Spain (16, 21, 35). However, unlike the emm75-ST150/657 isolates previously circulating in Portugal and the emm75-ST49 isolates identified in Spain and Houston (21, 35), which carry mef(A)-msr(D) in ɸ1207.3, the emm75-ST49 isolates from this study harbor a new ɸ1207.3 variant that likely emerged through recombination between ɸ1207.3 and other phages (Fig. 3). Further studies are necessary to determine if this variant encodes a functional and transferable phage.
Macrolide-resistant emm2 isolates have been infrequently isolated in Portugal and elsewhere (7, 9, 42). Despite the low genetic diversity of the national and international emm2 isolates, the closer relatedness of the resistant isolates in this study to international macrolide-susceptible isolates than to previous national resistant isolates is consistent with a recent introduction of an emm2-ST55 lineage carrying mef(A)-msr(D) in Portugal, possibly resulting from a novel acquisition of ɸ1207.3 by previously susceptible isolates. The two mef(A)-negative isolates genetically closer to our macrolide-resistant isolates (Fig. S2) were each temporally closer to its linked resistant isolate, despite being geographically distant (a US isolate from 2015 linked to a Portuguese 2019 resistant isolate and an Australian isolate from 1994 linked to a Portuguese 1998 resistant isolate), further supporting the occurrence of two independent phage acquisitions.
The main limitation of this study is that isolates were collected at a single hospital center, covering a limited geographic area. As a result, the findings may not fully represent the national antimicrobial resistance rates or genetic diversity and could be more influenced by any local outbreaks. In this regard, although emm2-ST55 isolates were detected from June to December 2019, continued surveillance across multiple hospitals is needed to determine whether the surge in this previously rare lineage was a regional event or part of a nationwide trend, and whether it persisted after the COVID-19 pandemic.
Our data showed that macrolide resistance is still low among streptococcal pharyngitis in Portugal. This occurs despite clonal shifts in macrolide-resistant lineages, often linked to changes in dominant macrolide resistance phenotypes and to resistance peaks. Moreover, macrolide resistance remains restricted to a few limited genetic lineages with marked clonal and epidemiological differences from macrolide-susceptible isolates.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Silva-Costa C, Friães A, Ramirez M, Melo-Cristino J. 2015. Macrolide-resistant Streptococcus pyogenes: prevalence and treatment strategies. Expert Rev Anti Infect Ther 13:615–628. doi:10.1586/14787210.2015.102329225746210 · doi ↗ · pubmed ↗
- 2Berbel D, González-Díaz A, López de Egea G, Càmara J, Ardanuy C. 2022. An overview of macrolide resistance in streptococci: prevalence, mobile elements and dynamics. Microorganisms 10:2316. doi:10.3390/microorganisms 1012231636557569 PMC 9783990 · doi ↗ · pubmed ↗
- 3Li Y, Rivers J, Mathis S, Li Z, Mc Gee L, Chochua S, Metcalf BJ, Fleming-Dutra KE, Nanduri SA, Beall B. 2023. Continued increase of erythromycin nonsusceptibility and clindamycin nonsusceptibility among invasive group A streptococci driven by genomic clusters, United States, 2018-2019. Clin Infect Dis 76:e 1266–e 1269. doi:10.1093/cid/ciac 46835684991 PMC 11120049 · doi ↗ · pubmed ↗
- 4World Health Organization. 2024. WHO bacterial priority pathogens list, 2024: bacterial pathogens of public health importance to guide research, development and strategies to prevent and control antimicrobial resistance. World Health Organization. Available from: https://www.who.int/publications/i/item/9789240093461. Retrieved 23 Jul 2024.
- 5Iannelli F, Santoro F, Santagati M, Docquier J-D, Lazzeri E, Pastore G, Cassone M, Oggioni MR, Rossolini GM, Stefani S, Pozzi G. 2018. Type M resistance to macrolides is due to a two-gene efflux transport system of the ATP-binding cassette (ABC) superfamily. Front Microbiol 9:1670. doi:10.3389/fmicb.2018.0167030108557 PMC 6079230 · doi ↗ · pubmed ↗
- 6Sharkey LKR, O’Neill AJ. 2018. Antibiotic resistance ABC-F proteins: bringing target protection into the limelight. ACS Infect Dis 4:239–246. doi:10.1021/acsinfecdis.7b 0025129376318 · doi ↗ · pubmed ↗
- 7Silva-Costa C, Ramirez M, Melo-Cristino J, Portuguese Group for Study of Streptococcal Infections. 2015. Declining macrolide resistance in Streptococcus pyogenes in Portugal (2007-13) was accompanied by continuous clonal changes. J Antimicrob Chemother 70:2729–2733. doi:10.1093/jac/dkv 18226142474 · doi ↗ · pubmed ↗
- 8Silva-Costa C, Pinto FR, Ramirez M, Melo-Cristino J, Portuguese Surveillance Group for the Study of Respiratory Pathogens. 2008. Decrease in macrolide resistance and clonal instability among Streptococcus pyogenes in Portugal. Clin Microbiol Infect 14:1152–1159. doi:10.1111/j.1469-0691.2008.02104.x 19046174 · doi ↗ · pubmed ↗