Thienopyrimidine amide analogs target MmpL3 in Mycobacterium tuberculosis
Vanessa Pietrowski Baldin, Christopher L. Harding, Diana Quach, Joseph Sugie, Joe Pogliano, Tanya Parish

TL;DR
This study shows that thienopyrimidine amide compounds likely target MmpL3, a key protein in the tuberculosis bacteria's cell wall, offering a new approach for drug development.
Contribution
The paper provides experimental evidence that TPA analogs target MmpL3 in Mycobacterium tuberculosis.
Findings
TPA analogs lost potency against M. tuberculosis strains with MmpL3 mutations.
TPAs induced cell wall stress and increased ATP production, consistent with MmpL3 inhibition.
Bacterial cytological profiling showed morphological changes matching known MmpL3 inhibitors.
Abstract
The identification of novel agents with mechanisms of action distinct from those currently utilized in tuberculosis treatment remains a significant challenge. The mycobacterial protein MmpL3 has emerged as a promising drug target due to its essential role in the synthesis of the cell wall of Mycobacterium tuberculosis. We previously identified novel thienopyrimidine amides(TPAs) with good anti-tubercular activity. We profiled a subset of TPAs, determining activity against intracellular bacteria and bactericidal activity against replicating bacteria. We ran assays to determine the mode of action by measuring cell wall stress, ATP production, and bacterial cytological profiling. We determined activity against a strain of M. tuberculosis with mutations in MmpL3. We isolated and sequenced resistant mutants. We tested five analogs against a strain of M. tuberculosis with mutations in MmpL3…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
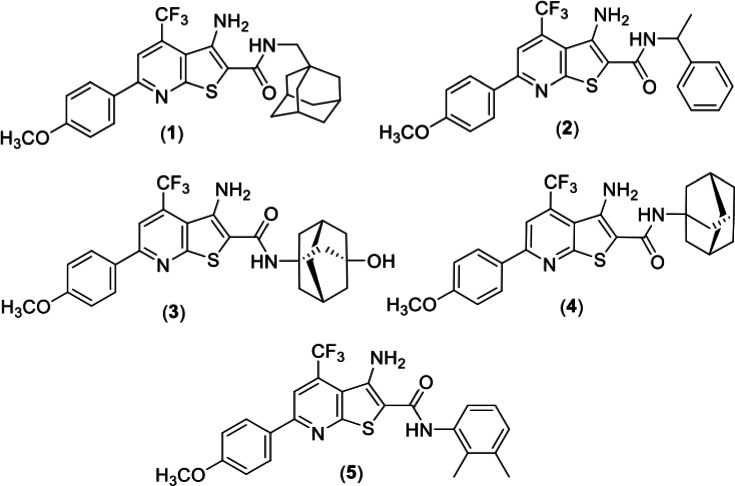 Fig 1
Fig 1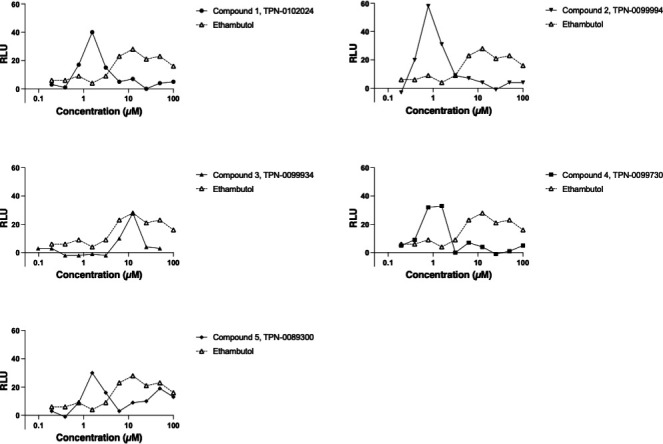 Fig 2
Fig 2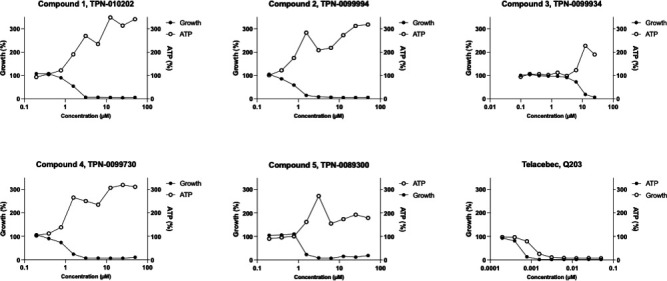 Fig 3
Fig 3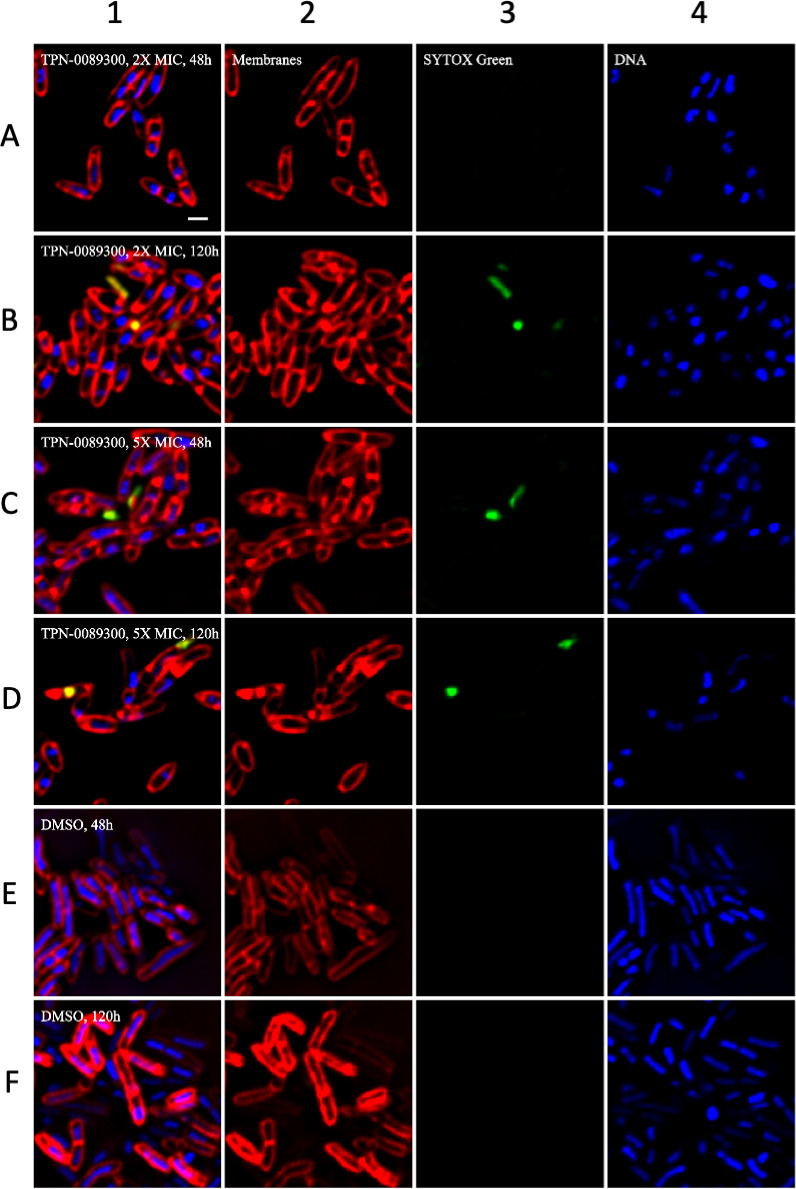 Fig 4
Fig 4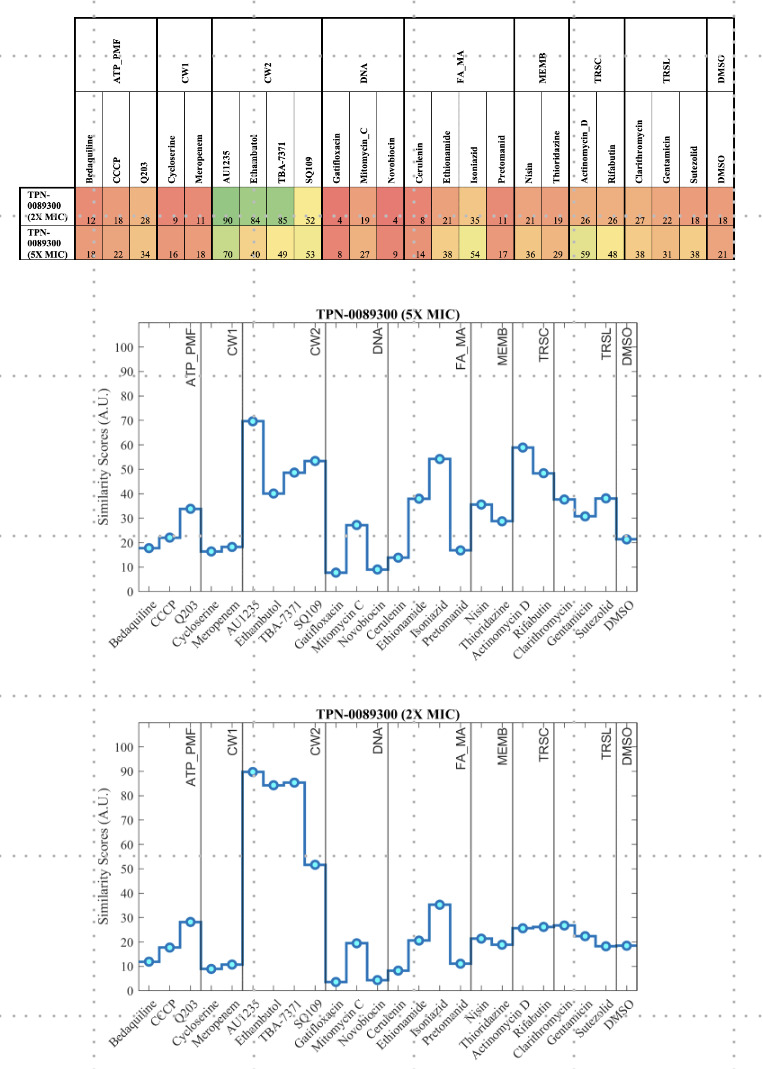 Fig 5
Fig 5| Intracellular | ExtracelluIar IC90 (µM) | MBC (µM) | ||||
|---|---|---|---|---|---|---|
| Molecule ID | IC90 (µM) | Wild-type | RM301 | Fold-change | Replicating | Non-replicating |
| (1) TPN-0102024 | 0.84 ± 0.2 | 1.4 ± 0.9 | >100 | 70 | 4.1 ± 1.8 | >100 |
| (2) TPN-0099994 | 0.18 ± 0.0 | 0.6 ± 0.2 | 3.5 ± 1.7 | 6 | 2.3 ± 1.3 | >100 |
| (3) TPN-0099934 | 1.5 ± 0.9 | 7.5 ± 3.3 | >50 | 7 | 21 ± 7.2 | >100 |
| (4) TPN-0099730 | 0.72 ± 0.2 | 1.1 ± 0.5 | >100 | >90 | 3.6 ± 2.4 | >100 |
| (5) TPN-0089300 | 0.32 ± 0.2 | 1.1 ± 0.5 | 3.9±0.7 | 39 | 3.1 ± 0 | >100 |
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —U.S. Department of Defensehttp://dx.doi.org/10.13039/100000005
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTuberculosis Research and Epidemiology · Mycobacterium research and diagnosis · Cancer therapeutics and mechanisms
INTRODUCTION
Despite the availability of drug treatment regimens, tuberculosis remains a major global health concern, with an estimated 8.2 million new cases reported in 2023, including 400,000 cases of drug-resistant tuberculosis (1). Currently, no vaccine provides effective protection against pulmonary tuberculosis (TB). Additionally, the bacteria can enter a latent phase, where they display no symptoms or cause symptoms that mimic other pathologies (2). Together, these factors highlight the urgent need for intensified efforts in TB control and the development of novel therapies, particularly those targeting drug-resistant bacilli and aiming to shorten the duration of treatment.
Mycobacterium tuberculosis is the causative agent of TB in humans. M. tuberculosis has a thick cell wall, which is composed of a highly hydrophobic bilayer of mycolic acids linked to arabinogalactan, in turn linked to peptidoglycan. This structure forms a unique barrier against drugs and the host immune defenses (3–5). Hence, cell wall biosynthesis represents an attractive target for the development of new inhibitors.
MmpL3 (Rv0206c) is an essential transporter which is highly conserved across the Mycobacterium genus (4, 6, 7) and is the only member of the MmpL family that is essential in M. tuberculosis (2, 4, 7). Its essentiality is attributed to its role in the translocation of trehalose monomycolate to the periplasmic space, where it serves as a precursor for the synthesis of trehalose dimycolate , a critical component of the cell envelope (4, 8). Given the important role of MmpL3 in constructing the mycobacterial cell wall, it has emerged as a high-figure value, druggable target, and at the moment, one of the most studied for anti-TB drug development (6). There are numerous MmpL3 inhibitors with diverse chemical structures, which have potent antibacterial activity (5, 9).
We previously identified and characterized novel thienopyrimidine amide (TPA) analogs that inhibit M. tuberculosis growth and had low cytotoxicity (10). From this series, we identified two subsets. One subset (TPA-L) had increased activity against a LepB (signal peptidase) hypomorph, indicating that the signal peptidase or protein secretion is the target or mode of action (10, 11). The second subset of molecules (TPA-M) had excellent potency but was equipotent against wild-type and hypomorph strains of M. tuberculosis, suggesting that the target was not protein secretion. In this paper, we explored the biological profile of the TPA-M series of molecules and identified MmpL3 as the most likely intracellular target.
MATERIALS AND METHODS
Bacterial strains and culture conditions
M. tuberculosis H37Rv-LP ATCC 25618 (wild-type) (12) and M. tuberculosis LP-0497754-RM301 (MmpL3 F255L, V646M, and F644I) (13) were cultured in Middlebrook 7H9 broth medium supplemented with 10% oleic acid, albumin, dextrose, and catalase (OADC) enrichment (BBL/Beckton, Dickson) and 0.05% w/v Tween 80 (7H9-Tw-OADC). Nutrient-starved cells were generated by resuspending cells in phosphate-buffered saline (PBS) with 0.05% w/v Tyloxapol (PBS-Tyl) at OD 1.0 and incubating for 7 days at 37°C without agitation. Avirulent M. tuberculosis mc^2^6206 (ΔpanCD ΔleuCD) was grown in 7H9-Tw-OADC supplemented with 0.5% wt/vol glycerol, 0.2% wt/vol Casamino Acids, 48 µg/mL pantothenate, and 50 µg/mL leucine at 30°C.
Determination of minimum inhibitory concentration (MIC)
TPA molecules (Fig. 1) were synthesized as previously described (10). Compound stocks were resuspended in DMSO and stored at −20°C. MICs were determined in 96- or 384-well microplates as described (14). Briefly, M. tuberculosis was grown to mid-log phase; cultures were dispensed into plates containing test compounds to a final OD_590_ of 0.02 and incubated at 37°C for 5 days (H37Rv) or 7 days (mc^2^6206). The OD_590_ was measured and IC_90_ determined as the concentration at which 90% of growth was inhibited as compared with controls (DMSO only).
Structure of analogs used in this study (1). TPN-0102024 (2) TPN-0099994 (3) TPN-0099934 (4) TPN-0099730 (5) TPN-0089300.
Determination of activity against intracellular bacilli
THP-1 cells were cultivated in RPMI-1640 medium supplemented with 10% FBS and incubated at 37°C, 5% CO_2_. Cells were treated with 80 nM PMA for 24 h prior to infection, harvested using Accumax solution, and resuspended in fresh cRPMI at a final density of 9 × 10⁵ cells/mL. Cells were infected overnight at a multiplicity of 1:1 with M. tuberculosis expressing LuxABCDE and exposed to compounds in 96-well plates for 72 h at 37°C and 5% CO_2_. Bacterial viability was assessed by luminescence; IC_90_ was determined as the concentration at which 90% of growth was inhibited as compared with controls (DMSO only).
Determination of bactericidal activity
M. tuberculosis H37Rv-LP was cultured in ^7^H9-Tw-OADC or starved for 7 days in PBS-Tyl, adjusted to a theoretical OD_590_ of 0.02 and exposed to compounds in 96-well plates. Cultures were spotted onto 7H10 agar supplemented with v/v 10% OADC7 (H10-OADC) on day 0, 7, and 14. The minmum bactericidal concentration (MBC) was defined as the lowest concentration at which no visible growth was observed. Niclosamide was included as a control.
Induction of cell wall stress
M. tuberculosis strain carrying the P_iniBAC_-lux reporter plasmid (15) was grown to mid-log in GAST/Fe protein-free medium containing 0.3 g/L Bacto Casitone, 0.05 g/L ferric ammonium citrate, 4 g/L dibasic anhydrous potassium phosphate, 2 g/L citric acid, 1 g/L L-alanine, 1.2 g/L magnesium chloride, 0.6 g/L potassium sulfate, 2 g/L ammonium chloride, 1.8 mL of 10 M sodium hydroxide, 10 mL glycerol, 5 mL 10% Tween 80, plus 15 µg/mL kanamycin. Cultures were adjusted to a theoretical OD_590_ of 0.02 and used to inoculate 96-well plates containing compounds. After 72 hours of incubation at 37°C, 1 vol of 10 mg/mL luciferin in 1M HEPES buffer pH 7.8, 1M DTT was added and RLU was read after 25 min of incubation at room temperature (RT). Ethambutol was included as a positive control.
Determination of ATP levels
ATP was measured using the BacTiter-Glo Assay kit (Promega) according to the manufacturer’s instructions. Log-phase M. tuberculosis was exposed to compounds for 24 h, .50 µL BacTiter-Glo was added, incubated at RT for 10 min and RLU read. Growth was measured after 120 h by OD. Q203 was included as a positive control.
Isolation of resistant mutants
Log-phase M. tuberculosis was plated on 7H10-OADC plates with 5× solid MIC and 10× solid MIC. Plates were incubated at 37°C until isolated colonies appeared. Potential resistant mutants were picked and streaked onto 5× MIC. MIC in liquid medium was measured to confirm resistance. Genomic DNA from three isolates was extracted from cultured cells by heat inactivation for 10 min at 100°C followed by 0.22 µm filtration. PCR amplification was performed using 10 µL of extract DNA in a final volume of 50 µL containing 1 µL of Pfu polymerase (Agilent), 5 µL of 10× Pfu amplification buffer (Agilent), 2.5 µL of primers MmpL3_seq_1_fwd (5’-gattcgctacctgagcag-3’) and MmpL3_seq_11_rev (0.5 µM) (5’-catttactgcagccgctg-3’) and 4 µL of 10 mM dNTP mix. PCR amplification was conducted, products were purified using the Qiagen PCR purification kit, and sequencing was performed by Plasmidsaurus using Oxford Nanopore Technology with custom analysis and annotation.
Bacterial cytological profiling
M. tuberculosis mc^2^6206 for bacterial cytological profiling was prepared as described (16). Briefly, cultures were adjusted to an OD_600_ of ~0.06–0.08 and incubated at 30°C for 18–20 hours before exposure to compounds at 1× and 5× MIC for 48h and 120 h. Cells were fixed using a mixture of 100 µL of 16% paraformaldehyde, 3 µL of 8% glutaraldehyde, and 20 µL of 0.4 M phosphate buffer pH 7.5, washed twice with 200 µL of warm medium, concentrated to approximately 30 µL, and stained for 30 min. Full-field fluorescence microscopy images of the samples were preprocessed from their original proprietary microscope file format into a common image format (TIFF). The original image dimensions (3×2048×2048) were cropped by trimming 124 pixels from each edge to avoid optical artifacts, resulting in dimensions of 3×1800×1800. A 600-pixel square sliding window was passed over this image with a step size of 60 pixels to produce 400 sub-images, which were each passed to a trained convolutional neural network. Each full-field image, consisting of 400 sub-images, generated a vector that describes a point in latent space. Similarity scores were calculated by comparing the location of unknown compound-treated cells in this space to control compound-treated cells using a scaled version of the average minimum distance.
RESULTS AND DISCUSSION
TPA analogs are active against intracellular mycobacteria
We previously demonstrated that the TPA series of compounds are active against aerobically-grown M. tuberculosis (10). A subset of analogs had potent antitubercular activity. Since these analogs had equipotent activity against wild-type and LepB hypomorph strains, we hypothesized that they did not target protein secretion. We wanted to determine the mode of action and target of these potent molecules.
We had already noted that compounds were active against replicating bacteria, so we expanded our work to look at activity against intracellular bacteria and non-replicating bacteria. We selected several active analogs (Fig. 1). Molecules were tested for activity against intracellular M. tuberculosis in THP-1 cells. All compounds were active against intracellular M. tuberculosis, with activity similar to the MICs (Table 1).
TPA analogs are bactericidal against replicating bacilli
We assessed the bactericidal activity of the TPA analogs against M. tuberculosis wild type following a 14-day exposure under replicating conditions as well as against non-replicating bacteria generated by nutrient starvation. All analogs were bactericidal against replicating M. tuberculosis with MBC/MIC ratio of <4. No bactericidal activity was seen against non-replicating cells (MBC >100 µM) (Table 1). These data are in contrast to what we would expect for inhibitors of LepB which showed bactericidal activity against non-replicating bacteria further supporting a different target or mode of action (17).
TPA analogs induce cell wall stress in M.
tuberculosis
We were interested in determining the mode of action of our molecules. As a first step, we determined whether molecules induced cell wall stress. We used a reporter strain of M. tuberculosis expressing luciferase under the control of P_iniBAC_, since induction of iniBAC expression is a marker of cell envelope stress (18). We saw induction of PiniBAC for all analogs, which predominantly occurred at concentrations close to the MIC. At higher concentrations, the induction is not seen due to the bactericidal activity of the molecules (dead cells cannot mount a transcriptional response). Of interest, the weakest stress response was seen with compound 3, which also demonstrated the weakest activity against M. tuberculosis with an MIC of 7.5 µM. These data suggest that TPA analogs cause cell wall stress (Fig. 2; Fig. S1).
Exposure to TPA analogs induces cell wall stress in M. tuberculosis. M. tuberculosis PiniBAC-Lux was exposed to compounds for 72 h and luminescence was read. Data are representative of two independent experiments (see Fig. S1).
TPA analogs boost ATP in M. tuberculosis
A link between induction of iniBAC and a burst in ATP production has previously been proposed (19). To determine if our molecules also affect ATP levels, we measured intracellular ATP levels following treatment with TPA analogs. As anticipated, increasing compound concentrations led to a boost in ATP levels (Fig. 3 Fig. S2). Again, we saw the boost occurring at concentrations around the MIC. These data are consistent with the response seen with other MmpL3 inhibitors (3, 20).
TPA analogs boost ATP in M. tuberculosis. M. tuberculosis was exposed to compounds for 24 h and ATP was measured using BacTiter-Glo. Growth was measured by OD after 5 d. Data are representative of two independent experiments (see Fig. S2). Q203 was used as a control. Data were normalized to positive and negative controls.
MmpL3 mutation leads to resistance to TPA analogs
We hypothesized that our molecules could be targeting MmpL3, since this is a highly promiscuous target and cell wall stress and elevated intracellular ATP levels are features associated with MmpL3 inhibition in M. tuberculosis (3). We compared TPA activity against the wild-type M. tuberculosis and a strain with mutations in MmpL3, which is resistant to a wide range of MmpL3 inhibitors (13, 20). The strain LP-0497754-RM301 carries three mutations in MmpL3-F255L,V646M, and F644I. The MmpL3 mutant strain LP-0497754-RM301 demonstrated increased resistance to all analogs, with at least a 4-fold increase in IC_90_ (Table 1). Interestingly, LP-0497754-RM301 was highly resistant to three of the five analogs which lost all activity (IC_90_ >50–100 µM). Thus, our data are consistent with MmpL3 being the drug target for this compound series.
Resistant strains have a non-synonymous mutation in
mmpL3
In order to determine whether there might be other targets, we isolated additional resistant mutants using TPN-0089300. Three strains were selected with an IC_90_ of >100 µM. Since MmpL3 mutations lead to resistance, we sequenced mmpL3 in three resistant isolates (RM3, RM4, and RM6). All isolates had the same non-synonymous mutation in MmpL3 of F644L. This mutation has been associated with resistance to other MmpL3 inhibitors and is similar to one of the mutations in the MmpL3 mutant strain we used to test for resistance (F644I) (4, 8, 13, 21).
Cytological profiling
We determined the phenotypic effect of a representative TPA analog using bacterial cytological profiling (16). We confirmed that TPN-0089300 was active against the avirulent M. tuberculosis strain mc^2^6206 (MIC of 1 µg/ml). Bacterial cells were exposed to TPN-0089300 for 48h and 120 h at 2× and 5× MIC (2 µg/ml and 5 µg/ml respectively). Morphological profiling demonstrated that bacterial morphology changed with cells becoming more rounded and losing membrane integrity after 120 h (Fig. 4). Based on the similarity score, this profile was identified as a strong match to other MmpL3 inhibitors (Fig. 5) (16, 20).
Cytological profiling in response to TPN-0089300. M. tuberculosis mc26206 was inoculated into medium at an OD of 0.06–0.08, treated with TPN-0089300 or DMSO (control) for 48120 h, and fixed with 16% paraformaldehyde, 8% glutaraldehyde, and 0.4 M phosphate buffer at pH 7.5. Cells were washed twice with a warm medium and stained for 30 min with FM 4–64 (Invitrogen), SYTO 40 (Invitrogen), and SYTOX Green (Invitrogen). Panel 1 – combined image; Panel 2 – membrane staining (red); Panel 3 – staining for membrane integrity using Sytox green (green) Panel 4 – DNA staining (blue). (A) TPN-0089300 at 2× MIC for 48; (B) TPN-0089300 at 2X MIC for 120 h; (C) TPN-0089300 at 5× MIC for 48; (D) TPN-0089300 at 5× MIC for 120 h; (E) DMSO for 48; (F) DMSO for 120 h.
Similarity scores for cytological profiling in response to TPN-0089300. Data from Fig. 4 was compared to a set of control compounds. Full-field fluorescence microscopy images were processed to produce 400 sub-images, which were each passed to a trained convolutional neural network. Each full-field image generated a vector that describes a point in latent space. Similarity scores were calculated by comparing the location of unknown compound-treated cells in this space to control compound-treated cells using a scaled version of the average minimum distance.
Conclusions
We provide strong evidence that a subset of TPA analogs, which we now term TPA-M, target MmpL3 in M. tuberculosis. TPA-M compounds were bactericidal for replicating bacilli, induced cell wall stress, and boosted ATP production. Mutations in MmpL3 conferred resistance, and bacterial cytological profiling identified MmpL3 as the most likely mode of action. We demonstrated that the TPA-M analogs do not kill starved, non-replicating bacilli. This is consistent with other MmpL3 inhibitors that have little effect against non-replicating M. tuberculosis (3, 22). Cells treated with MmpL3 inhibitors develop dimples at the dividing pole, where the protein localizes during active cell division and the lack of activity against non-replicating cells has been attributed to the absence of MmpL3 localization at the pole in the non-replicating state (7, 23). Taken together, our data suggest that MmpL3 is the likely target of the TPA-M series.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1World Health Organization. 2024. Global tuberculosis report 2024. Available from: https://www.who.int/teams/global-tuberculosis-programme/tb-reports/global-tuberculosis-report-2024
- 2Diab A, Dickerson H, Al Musaimi O. 2025. Targeting the heart of Mycobacterium: advances in anti-tubercular agents disrupting cell wall biosynthesis. Pharmaceuticals (Basel) 18:70. doi:10.3390/ph 1801007039861133 PMC 11768153 · doi ↗ · pubmed ↗
- 3Ames L, Allen R, Boshoff HI, Cleghorn L, Engelhart C, Schnappinger D, Parish T. 2025. Common biological features of Mycobacterium tuberculosis Mmp L 3 inhibitors. ACS Inf Dis 11:2523–2533. doi:10.1021/acsinfecdis.5c 00394 PMC 1244205740827527 · doi ↗ · pubmed ↗
- 4Kapp E, Calitz H, Streicher EM, Dippenaar A, Egieyeh S, Jordaan A, Warner DF, Joubert J, Malan SF, Sampson SL. 2023. Discovery and biological evaluation of an adamantyl-amide derivative with likely Mmp L 3 inhibitory activity. Tuberculosis (Edinb) 141:102350. doi:10.1016/j.tube.2023.10235037244249 · doi ↗ · pubmed ↗
- 5North EJ, Schwartz CP, Zgurskaya HI, Jackson M. 2023. Recent advances in mycobacterial membrane protein large 3 inhibitor drug design for mycobacterial infections. Expert Opin Drug Discov 18:707–724. doi:10.1080/17460441.2023.221808237226498 PMC 10330604 · doi ↗ · pubmed ↗
- 6Adams O, Deme JC, Parker JL, C Ry PTIC Consortium, Fowler PW, Lea SM, Newstead S. 2021. Cryo-EM structure and resistance landscape of M. tuberculosis Mmp L 3: an emergent therapeutic target. Structure 29:1182–1191. doi:10.1016/j.str.2021.06.01334242558 PMC 8752444 · doi ↗ · pubmed ↗
- 7Williams JT, Abramovitch RB. 2023. Molecular mechanisms of Mmp L 3 function and inhibition. Microb Drug Resist 29:190–212. doi:10.1089/mdr.2021.042436809064 PMC 10171966 · doi ↗ · pubmed ↗
- 8Williams JT, Haiderer ER, Coulson GB, Conner KN, Ellsworth E, Chen C, Alvarez-Cabrera N, Li W, Jackson M, Dick T, Abramovitch RB. 2019. Identification of new Mmp L 3 inhibitors by untargeted and targeted mutant screens defines Mmp L 3 domains with differential resistance. Antimicrob Agents Chemother 63:e 00547-19. doi:10.1128/AAC.00547-1931405862 PMC 6761494 · doi ↗ · pubmed ↗