Ophiostomatoid fungi associated with Hylurgus ligniperda, including six new species from eastern China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
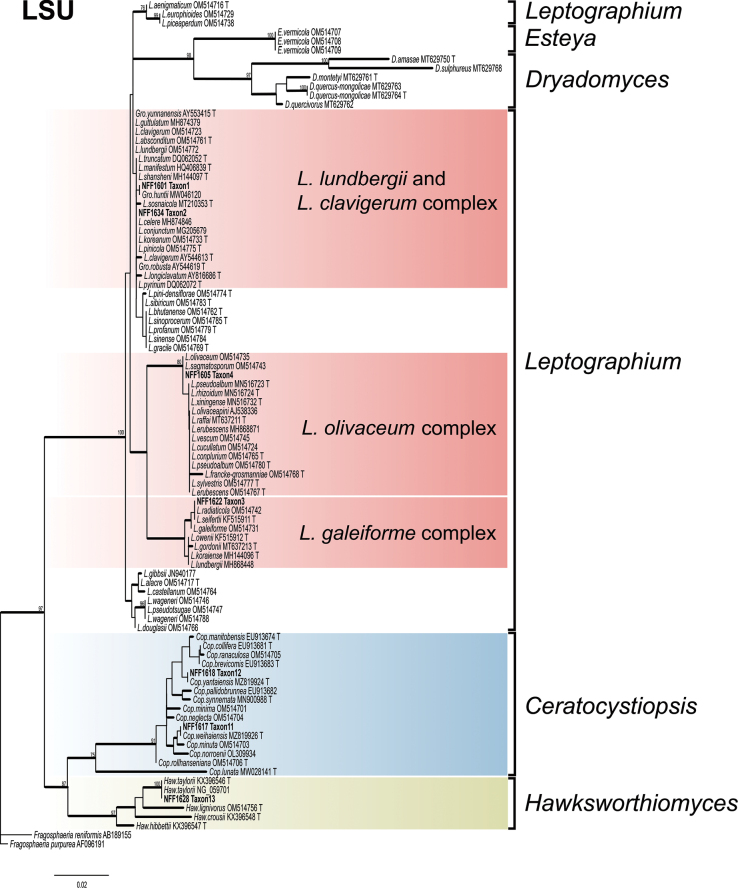 Figure 1
Figure 1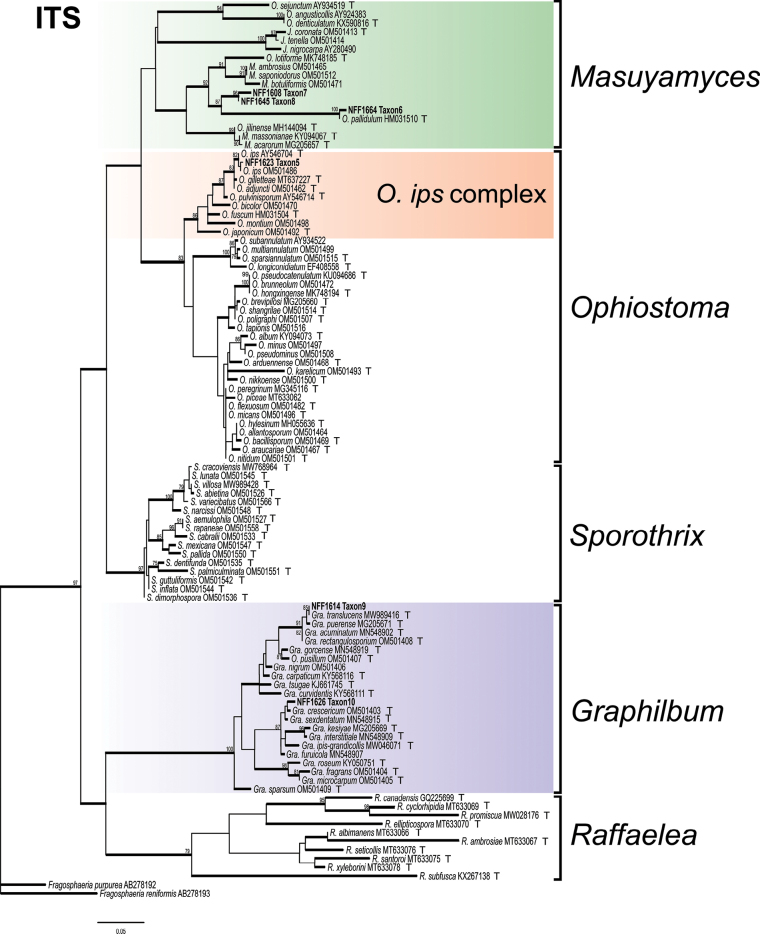 Figure 2
Figure 2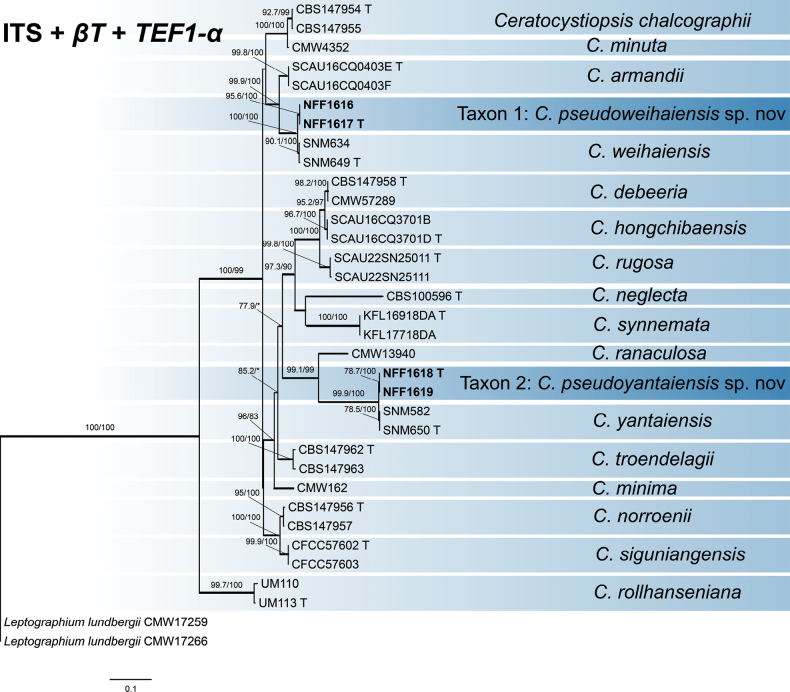 Figure 3
Figure 3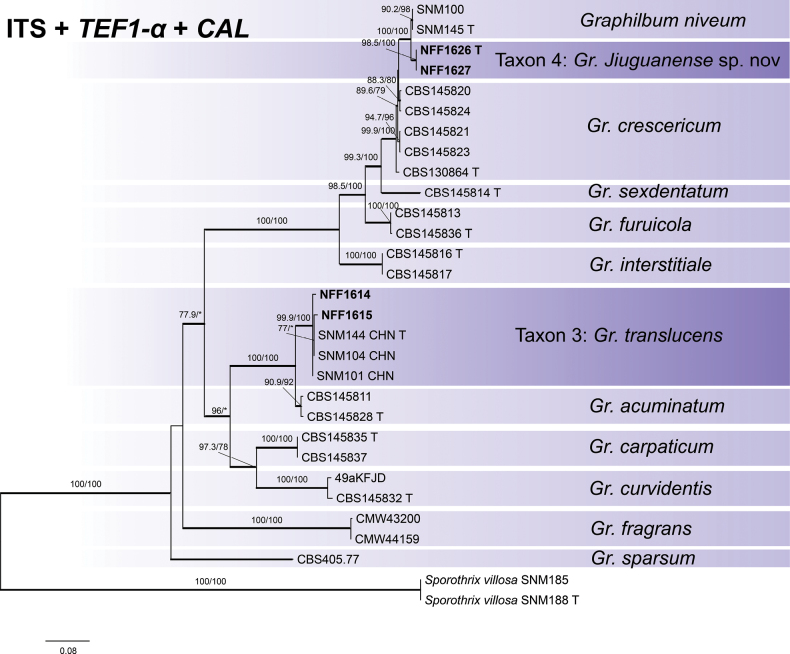 Figure 4
Figure 4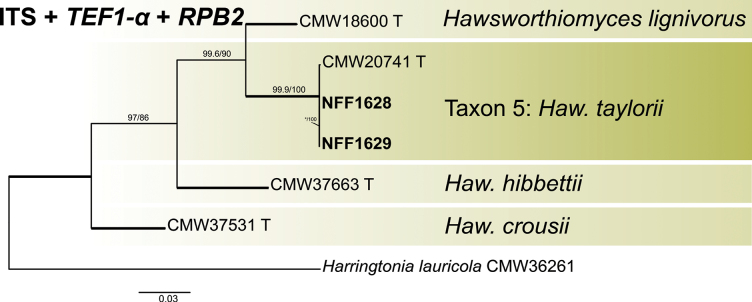 Figure 5
Figure 5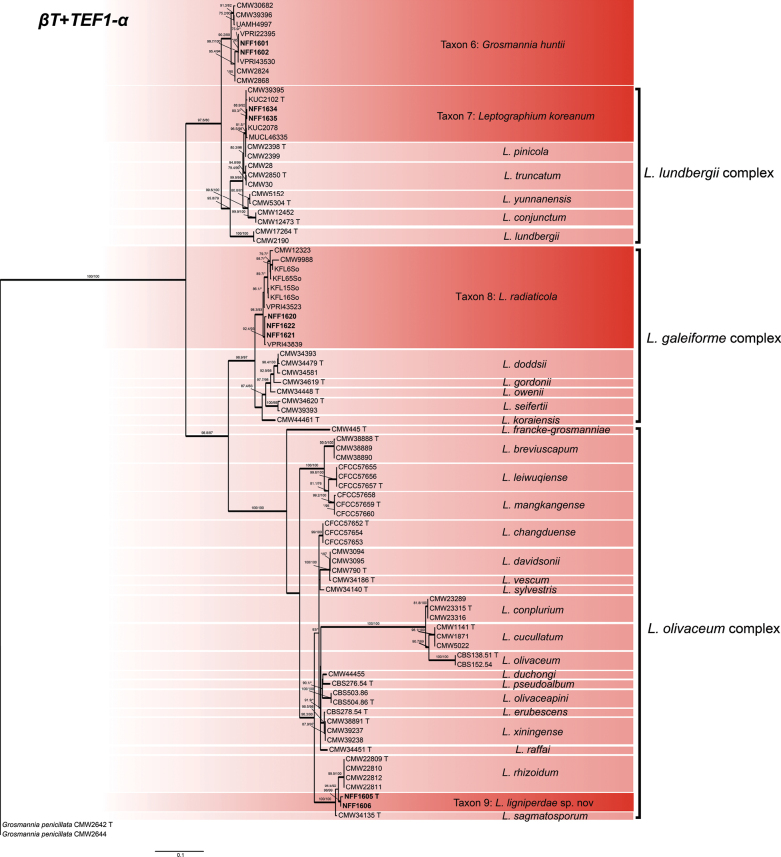 Figure 6
Figure 6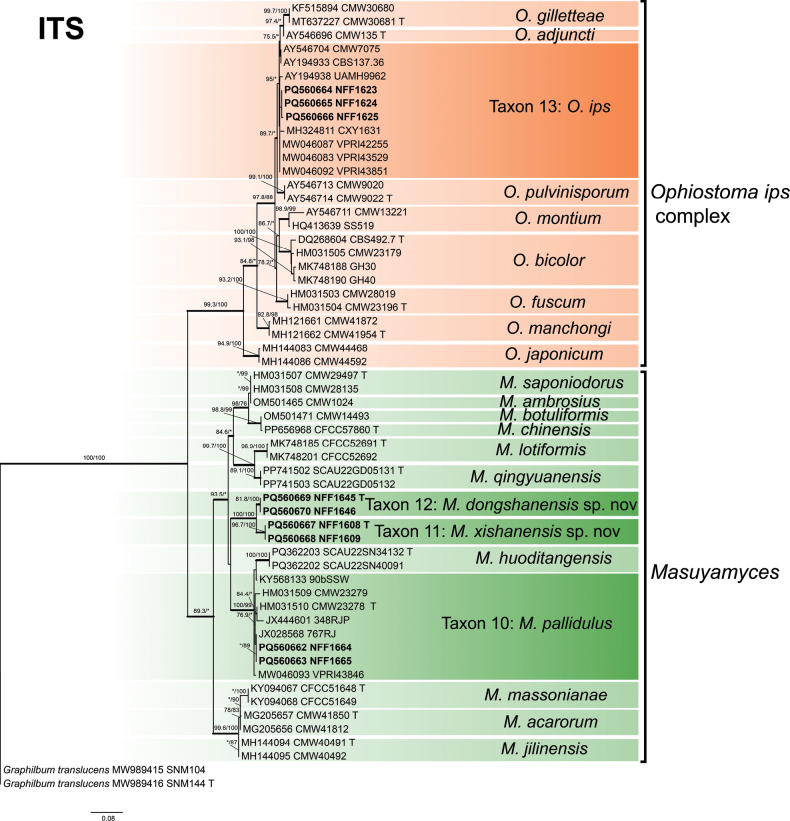 Figure 7
Figure 7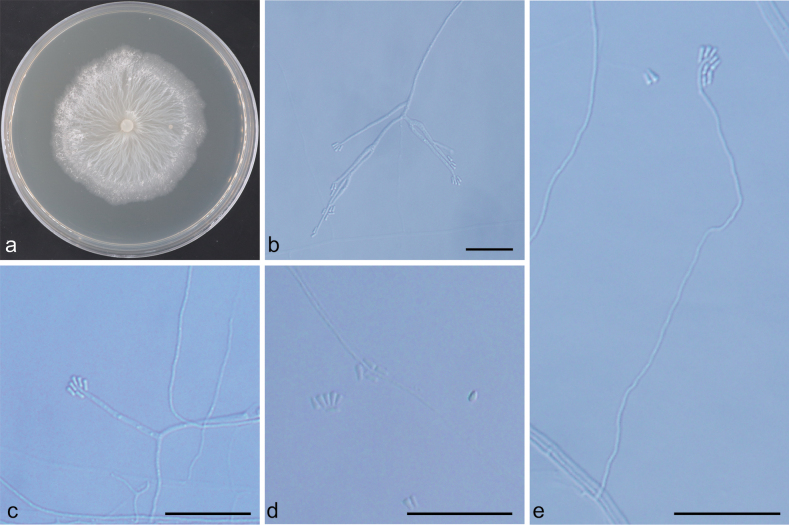 Figure 8
Figure 8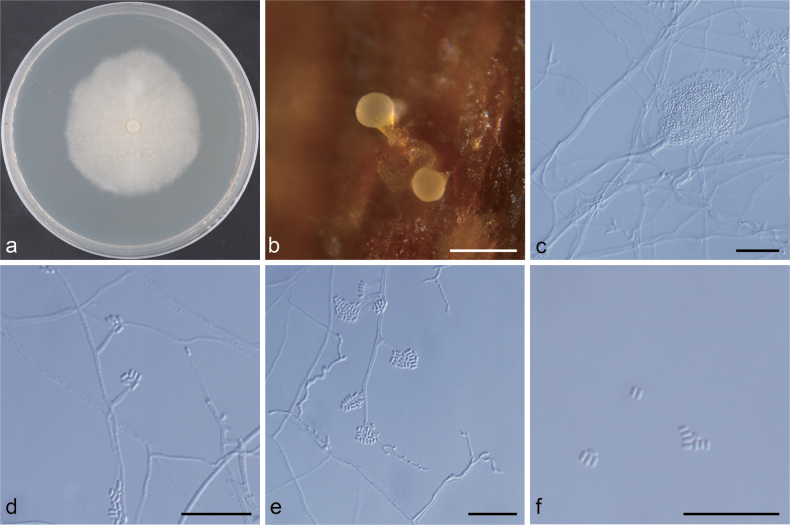 Figure 9
Figure 9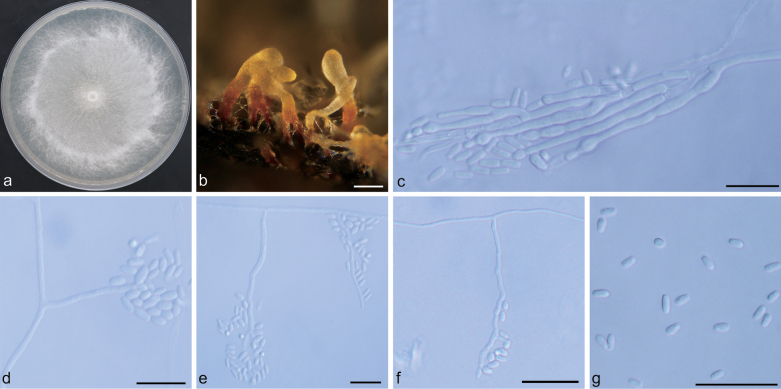 Figure 10
Figure 10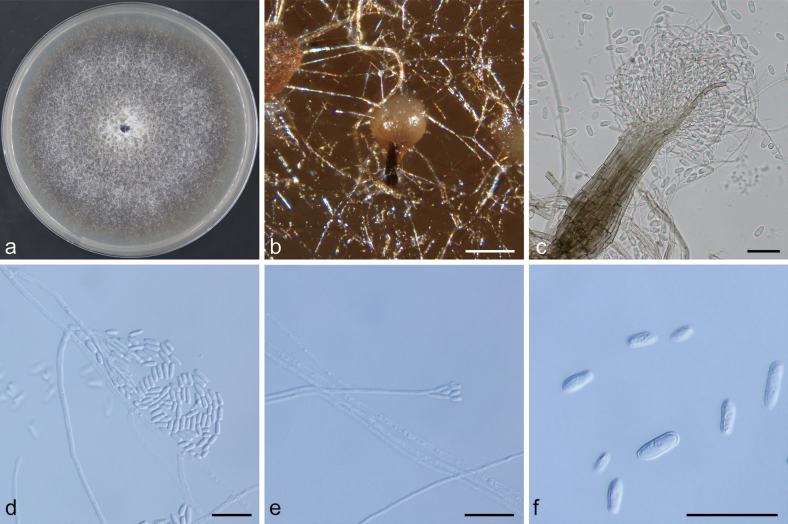 Figure 11
Figure 11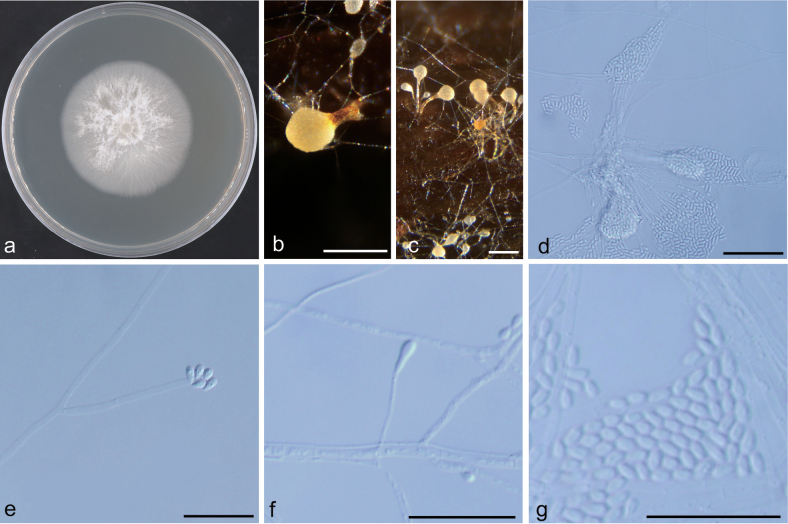 Figure 12
Figure 12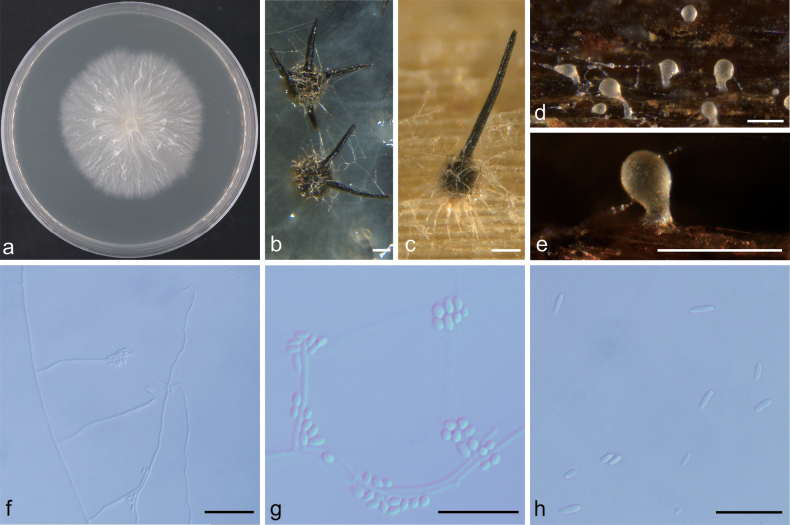 Figure 13
Figure 13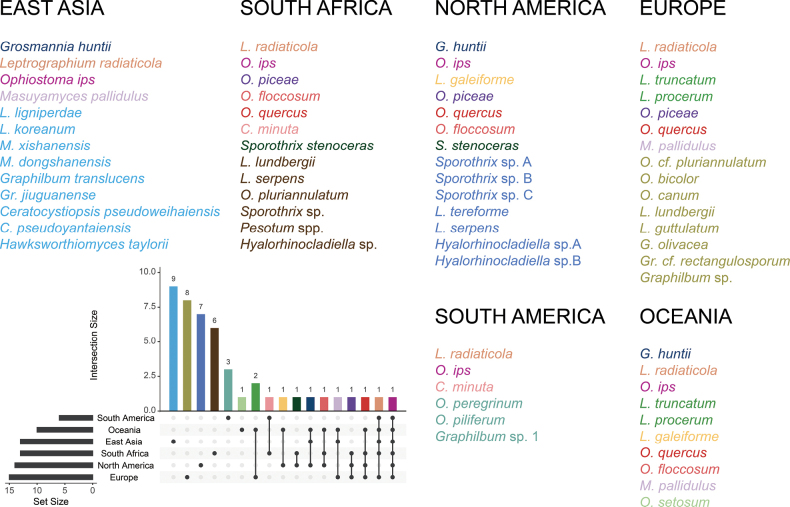 Figure 14
Figure 14| Taxon | Species name1 | Isolate numbers2,3,4 | GenBank5 | |||||
|---|---|---|---|---|---|---|---|---|
| ITS |
|
| βT |
|
| |||
| 1 | NFF1616 |
|
|
|
| - |
| |
| NFF1617(CGMCC3.28600)T |
|
|
|
| - |
| ||
| 2 | NFF1618(CGMCC3.28601)T |
|
|
|
| - |
| |
| NFF1619 |
|
|
|
| - |
| ||
| 3 |
| NFF1614 |
| - |
|
|
|
|
| NFF1615 |
| - |
|
|
|
| ||
| 4 | NFF1626(CGMCC3.28602)T |
| - |
|
|
|
| |
| NFF1627 |
| - |
|
|
|
| ||
| 5 |
| NFF1628 |
|
|
|
|
|
|
| NFF1629 |
|
|
|
|
|
| ||
| 6 |
| NFF1601 |
|
|
|
|
|
|
| NFF1602 |
|
|
|
|
|
| ||
| 7 |
| NFF1634 |
|
|
|
|
|
|
| NFF1635 |
|
|
|
|
|
| ||
| 8 |
| NFF1620 |
|
|
|
|
|
|
| NFF1621 |
|
|
|
|
|
| ||
| NFF1622 |
|
|
|
|
|
| ||
| 9 | NFF1605(CGMCC3.28598)T |
|
|
|
|
|
| |
| NFF1606 |
|
|
|
|
|
| ||
| 10 |
| NFF1664 |
| - |
|
|
|
|
| NFF1665 |
| - |
|
|
|
| ||
| 11 | NFF1608(CGMCC3.28599)T |
| - |
|
| - |
| |
| NFF1609 |
| - |
|
| - |
| ||
| 12 | NFF1645(CGMCC3.28603)T |
| - |
|
| - |
| |
| NFF1646 |
| - |
|
| - |
| ||
| 13 |
| NFF1623 |
| - |
|
|
|
|
| NFF1624 |
| - |
|
|
|
| ||
| NFF1625 |
| - |
|
|
|
| ||
| Genus/Other level | Dataset (Positions) | Nucleotide substitution models | |
|---|---|---|---|
| IQ-TREE | Mrbayes | ||
| ITS complex (438 bp) | GTR + F + I + G4 | GTR + F + I + G4 | |
| GTR + F + I + I + R2 | GTR + F + I + G4 | ||
|
| ITS (627 bp) | GTR + F + I + I + R2 | GTR + F + G4 |
| TIM2 + F + I + G4 | GTR + F + I + G4 | ||
| TN + F + G4 | GTR + F + G4 | ||
| ITS + | ITS: GTR+F+I+I+R2; | ITS: GTR + F + G4; | |
|
| ITS (621 bp) | SYM + G4 | SYM + G4 |
| TN + F + I + G4 | GTR + F + I + G4 | ||
| GTR + F + I + G4 | GTR + F + I + G4 | ||
| ITS + | ITS: TIMe + R2; | ITS: SYM + G4; | |
|
| ITS + | ITS: TIM + F + G4; | ITS: GTR + F + G4; |
|
| |||
| TN + F + R2 | HKY + F + G4 | ||
| TIM3 + F + G4 | GTR + F + G4 | ||
| TIM3 + F + G4 | GTR + F + G4 | ||
| ITS (461 bp) | GTR + F + I + I + R2 | GTR + F + G4 | |
| TIM3 + F + R4 | GTR + F + G4 | ||
| Genus | Taxon | Species1 | Numbers of isolates (n = 326) 2,3 | Total | Total isolation frequency (%) | |
|---|---|---|---|---|---|---|
|
| 1 | 11(5.42) | 0 | 11 | 2.53 | |
| 2 | 11(5.42) | 0 | 11 | 2.53 | ||
|
| 3 |
| 15(7.39) | 0 | 15 | 3.45 |
| 4 | 9(4.43) | 0 | 9 | 2.07 | ||
|
| 5 |
| 11(5.42) | 12(5.17) | 23 | 5.29 |
|
| 6 |
| 9(4.43) | 27(11.64***) | 36 | 8.28 |
| 7 |
| 15(7.39) | 36(15.52***) | 51 | 11.72 | |
| 8 |
| 14(6.90) | 51(21.98***) | 65 | 14.94 | |
| 9 | 0 | 9(3.88) | 9 | 2.07 | ||
|
| 10 |
| 17(8.37) | 14(6.03) | 31 | 7.13 |
| 11 | 7(3.45) | 0 | 7 | 1.61 | ||
| 12 | 8(3.94) | 0 | 8 | 1.84 | ||
|
| 13 |
| 76(37.44) | 83(35.78) | 159 | 36.55 |
| Total | 203(46.67) | 232(53.33) | 435 | 100.00 | ||
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsForest Insect Ecology and Management · Plant and Fungal Interactions Research · Entomopathogenic Microorganisms in Pest Control
Introduction
Global climate change and trade globalization have significantly influenced the proliferation and distribution of forest pests, posing novel challenges for forest management. Bark beetles (Coleoptera, Curculionidae, Scolytinae), a highly diverse group of insects, are prevalent across global forest ecosystems (Morris et al. 2017). These beetles, which primarily target coniferous trees, exhibit strong concealment abilities and can induce extensive tree mortality, thus representing critical forest pests (Ferrenberg et al. 2014). Research indicates a strong correlation between the spread of bark beetles and climate warming (Niemiec et al. 2014). When conditions are favorable, particularly when introduced beyond their native habitats, certain species can transition from minor pests to significant threats to host trees, resulting in substantial economic losses and ecological harm (Ranger et al. 2015; Ranger 2016; Biedermann et al. 2019). In Europe, Ips typographus Linnaeus is regarded as the most significant pest affecting Picea abies (L.) H. Karst. (Schelhaas et al. 2003). Similarly, in China, the invasive alien species Dendroctonus valens LeConte has led to widespread mortality of Pinus tabuliformis Carrière, posing a serious threat to the ecological stability of forestry (Lu et al. 2009; Sun et al. 2013).
In comparison to isolated damage, the combined impact of microbial pathogens and insects can result in severe harm to forest ecosystems (Kenneth F. Raffa and Romme 2008). Consequently, it is imperative to promptly elucidate the novel interactions between forest pests and pathogens (Wingfield et al. 2016; Biedermann and Vega 2020). Ophiostomatoid fungi are primarily associated with bark beetles. These fungi exhibit similar morphological and structural traits, notably the production of adhesive spore masses, which are disseminated by bark beetles to host plants (De Beer 2013). Ophiostomatoid fungi are integral to the successful colonization of bark beetles (Hofstetter et al. 2015), significantly compromising the resistance of host trees and facilitating the invasion by these beetles (Heiniger et al. 2011). Furthermore, these fungi contribute essential nutrients for the growth and development of bark beetles within the nutrient-deficient phloem of coniferous trees (Bentz and Six 2006). Bark beetles associate with a diverse array of ophiostomatoid fungal species, including several highly pathogenic strains (Kgang et al. 2008) and staining fungi (Hýsek et al. 2021), the latter of which can adversely affect wood quality. This group of fungi are regarded as a significant threat to conifers, prompting extensive research of bark beetle fungi globally. In southeastern Australia, fourteen species of Ophiostomatales across six genera, along with one species of Microascales, have been isolated from bark beetles and infected pines (Trollip et al. 2021). In Northeast China, Ips subelongatus Motschulsky is associated with fourteen species of ophiostomatoid fungi spanning four genera (Ceratocystiopsis, Endoconidiophora, Leptographium, and Ophiostoma) (Wang et al. 2020). Notably, Ophiostoma bicolor R.W. Davidson & D.E. Wells, O. brunneo-ciliatum Math.-Käärik, O. ainoae H. Solheim, and Grosmannia piceiperda (Rumbold) Goid are associated with both Ips typographus and Pityogenes chalcographus Linnaeus (Repe et al. 2013).
Hylurgus ligniperda Fabricius (1787), commonly referred to as the red-haired pine bark beetle, is a forest insect that parasitizes the phloem of pine trees (Bedoya et al. 2019). It is presumed to be indigenous to Europe and the Mediterranean Basin, with subsequent introductions into Oceania, North America, and South America. In East Asia, H. ligniperda is considered an invasive species, initially invading Japan and subsequently Korea. Recently, it has successfully invaded and colonized Shandong Province in China (Lin et al. 2021). As a root-dwelling species, H. ligniperda typically enters its host through the soil and feeds on the phloem of sub-healthy pine trees, although it does not directly cause tree mortality. However, in newly invaded regions, the presence of H. ligniperda significantly affects the ecological security of local forestry systems. The invasion of H. ligniperda has resulted in a reduction of the windbreak and sand-fixation capacities of the coastal shelterbelt in Shandong Province, China, and its high dispersal capability poses a substantial threat to the local forest ecosystem.
Numerous studies have documented the isolation of fungi associated with H. ligniperda. In the forests and steppes of eastern Ukraine, H. ligniperda serves as a vector for diverse fungal communities (Davydenko et al. 2014). In Chile, five ophiostomatoid fungi species associated with H. ligniperda and Hylastes ater Paykull have been identified (Zhou et al. 2004c). Eight species of ophiostomatoid fungi have been isolated from H. ligniperda in New Zealand (Ray et al. 2006). In Poland, the ophiostomatoid fungi associated with H. ater, Hylastes opacus Erichson and H. ligniperda on Pinus sylvestris L. was examined, revealing that fungi of the genera Grosmannia and Leptographium were the most prevalent (Jankowiak and Bilański 2013). However, in the newly invaded region of H. ligniperda in Shandong Province, China, research of fungi associated with H. ligniperda remains unexplored.
In this study, we examined the ophiostomatoid fungi community associated with H. ligniperda in the Yanwei region of Shandong Province, China. We employed a combination of morphological observations and multilocus DNA sequence phylogenetic analysis to address existing knowledge gaps in this area. The findings of this research contribute to a deeper understanding of the ophiostomatoid fungi community linked to H. ligniperda in China and establish a foundation for future investigations into the occurrence and management of H. ligniperda.
Materials and methods
Collection of samples and isolation of fungi
Adults of H. ligniperda were collected from coastal shelterbelts in Yantai and Weihai, Shandong Province, between December 2021 and April 2023. The forest stands consisted primarily of ~45-year-old plantations of Pinus thunbergii Parlatore, with a minor presence of similarly aged Pinus densiflora Siebold & Zucc. Using sterile forceps, adults of H. ligniperda collected from traps and galleries were individually transferred into 2 mL sterile centrifuge tubes, transported to the laboratory, and stored at 4 °C until fungal isolation. The adults of H. ligniperda were subjected to disinfection for one minute using a 0.1% sodium hypochlorite (NaClO) solution, followed by three rinses with sterile water. Subsequently, the specimens were fragmented into several parts, with each part placed at a specific distance on 2% malt extract agar (MEA). All cultures were incubated in darkness at 25 °C and monitored daily. To purify the fungal isolates, hyphal tips from the fungal colonies were transferred onto 2% MEA plates. Following an initial analysis of colony and morphological characteristics, representative strains of each morphotype were selected for further studies. All strains were deposited in the Forest Protection Department of Forestry College, Northeast Forestry University, Harbin, Heilongjiang Province, China (NFF) (Table 1). Ex-type cultures of ophiostomatoid fungi described in this study are deposited in the China General Microbiological Culture Collection Center (CGMCC), and the holotype specimens (dry cultures) were deposited in the Herbarium Mycologicum, Academiae Sinicae (HMAS), Beijing, China.
Table 1.: Representative strains of the ophiostomatoid fungi associated with adult Hylurgus ligniperda in Yantai and Weihai, Shandong Province, China.
Morphological and cultural studies
The representative strains were cultured in a 2% MEA medium at 25 °C in a dark environment for a period of 7 to 15 days to facilitate morphological characterization. To examine the morphology of the newly identified species, fungal strains were first inoculated onto sterilized phloem tissues of Pinus thunbergii and cultured in darkness at 25 °C. Colony morphology and growth were observed daily. After 4–5 weeks, the phloem tissues were mounted on microscope slides to examine sexual or asexual structures. An Olympus BX51 (Olympus, Tokyo, Japan) microscope and a Leica Z16 APO (Leica, Leica Microsystems Ltd., Switzerland) stereomicroscope were employed to measure and photograph the microstructure of the ophiostomatoid fungi. The growth structures of each strain, including the length and width of the conidia and conidiophores, were observed and measured on microscope slides prepared with sterile water. A total of 30 measurements were conducted for the reproductive structures of each strain. The results are reported as the average (mean), standard deviation (SD), minimum (min), and maximum (max) values, presented in the format: (min –) (mean – SD) – (mean + SD) (– max).
The growth rate was assessed using the following methodology: a sterile hole punch was employed to extract a 5 mm diameter sample from the periphery of strains exhibiting robust mycelial growth. This sample was then positioned, with the mycelium side facing downward, at the center of a 100 mm Petri dish containing 25 ml of 2% malt extract agar (MEA). For each strain, five replicate plates were prepared and incubated in darkness at temperature intervals of 5 °C, ranging from 5 °C to 35 °C. Starting from the second day of incubation, the orthogonal diameters of the colonies were measured daily until the most rapidly growing mycelium reached the edge of the 2% MEA plate. Colony colors were described based on the color chart of Rayner (1970). All data related to the type specimen are deposited in MycoBank (http://www.MycoBank.org/).
DNA extraction, PCR, and sequencing
DNA extraction was performed using an Invisorb Spin Plant Mini Kit (Tiangen, Beijing, China) from actively growing mycelia, following the manufacturer’s instructions. The internal transcribed spacer regions 1 and 2 of the nuclear ribosomal DNA operon, including the 5.8S region (ITS), the ribosomal large subunit region (LSU), the β-tubulin gene region (βt), the transcription elongation factor 1-α gene region (TEF-1α), the calmodulin gene region (CAL), and the DNA-directed RNA polymerase II second largest subunit gene region (RPB2) were subjected to PCR using the 2 × Taq PCR MasterMix (Tiangen, Beijing, China), following the manufacturer’s instructions. The primer pairs used for amplification included ITS1-F/ITS4 (Gardes and Bruns 1993)(White et al. 1990), LR0R/LR5 (Vilgalys and Hester 1990), Bt2a/Bt2b (Glass and Donaldson 1995), EF2F/EF2R (Marincowitz et al. 2015)(Jacobs et al. 2004), CL2F/CL2R (Duong et al. 2012), and Oph-RPB2F1/Oph-RPB2R1 or Oph-RPB2F2/Oph-RPB2R1 (De Beer et al. 2022). The ITS, LSU, βT, TEF-1α, and CAL gene regions were amplified according to the protocol described by Wang et al. (2020), and the RPB2 gene region was amplified according to the protocol described by De Beer et al. (2022). All PCR products were sequenced by Sangon Biotech, Changchun, Jilin Province, China. All sequences obtained in this study were deposited in GenBank (Table 1) (www.ncbi.nlm.nih.gov/Genbank).
Phylogenetic analyses
The sequences derived from the six DNA fragments were initially identified through a BLAST search in the NCBI GenBank database. Representative sequences exhibiting the highest similarity, along with model strain sequences of closely related species, were subsequently retrieved from the NCBI GenBank database. The GenBank accession numbers for these sequences are indicated on the corresponding phylogenetic trees. Sequence datasets were aligned using MAFFT v7.505 (Katoh and Standley 2013) and trimmed at both termini as necessary. To eliminate ambiguously aligned regions, all datasets were processed using Gblocks 0.91b (Talavera and Castresana 2007) with less stringent parameters. PhyloSuite v1.2.3 (Zhang et al. 2020) was employed to concatenate the datasets of individual gene regions obtained from Gblocks. Subsequently, PhyloSuite v1.2.3 facilitated the construction of multilocus phylogenetic trees using Maximum Likelihood (ML) and Bayesian Inference (BI) methods.
The ModelFinder v2.2.0 (Kalyaanamoorthy et al. 2017) was used to select the best-fit partition model (Edge-linked) using Akaike information criterion (AIC) (Table 2). Maximum likelihood (ML) phylogenies were inferred using IQ-TREE v2.2.0 (Nguyen et al. 2014) under Edge-linked partition model for 1000 standard bootstraps, as well as the Shimodaira-Hasegawa-like approximate likelihood-ratio test. Bayesian Inference (BI) phylogenies were inferred using MrBayes v3.2.7a (Ronquist et al. 2012) under partition model (2 parallel runs, 5000000 generations), until the average standard deviation of split frequencies was < 0.01. Trees were sampled every 100 generations, in which the initial 25% of sampled data were discarded as burn-in. The remaining trees were used to calculate the posterior probabilities. The phylogenetic tree was edited in FigTree v.1.4.3 (http://tree.bio.ed.ac.uk/software/figtree/) and Adobe Illustrator CS6. The final alignment results and the derived topology were archived in TreeBASE (No. 31864).
Statistical analysis
The isolation frequencies of fungal taxa common to adults obtained from galleries and traps were compared. Statistical significance was determined using a chi-square test in SPSS Statistics 20.0 (SPSS Inc.).
Results
Collection of samples and isolation of fungi
In this study, 435 strains of ophiostomatoid fungi were isolated from 326 adult beetles. Preliminary identification was conducted based on growth rate and macroscopic and microscopic morphological characteristics. All strains were preserved and DNA extracted. Standard nucleotide BLAST searches at GenBank were performed using the ITS and LSU sequences of all strains for preliminary classification and affinity assessment. Subsequently, 28 representative strains were selected for comprehensive morphological examination and multilocus phylogenetic analysis.
Phylogenetic analysis
Phylograms generated by ML are presented for all datasets, with branch supports indicated from both ML and BI analyses. The phylogenetic analysis at the genus level was first conducted based on the ITS and LSU regions (Schoch et al. 2012), followed by a more precise species-level delineation utilizing additional gene regions (namely βT, TEF1-α, CAL, and RPB2) and their concatenated dataset (De Beer et al. 2016a; De Beer et al. 2022). The phylogenetic analysis revealed that 28 representative isolates were distributed across 13 terminal clades or phylogenetic groups, spanning six genera within the Ophiostomatales (Figs 1, 2). The results of the phylogenetic analysis are presented as follows: Ceratocystiopsis (Taxa 1–2), Graphilbum (Taxa 3–4), Hawksworthiomyces (Taxon 5), Leptographium (Taxa 6–9), Masuyamyces (Taxa 10–12), and Ophiostoma (Taxon 13).
Maximum Likelihood phylogeny of LSU region for isolates residing in Leptographium, Ceratocystiopsis, and Hawksworthiomyces. Sequences generated in this study are printed in bold. Bold branches indicate posterior probability values ≥ 0.9, while ML bootstrap values of ≥ 75% are recorded at nodes. T = ex-type cultures.
Maximum Likelihood phylogeny of ITS region for isolates residing in Masuyamyces, Ophiostoma, and Graphilbum. Sequences generated in this study are printed in bold. Bold branches indicate posterior probability values ≥ 0.9, while ML bootstrap values of ≥ 75% are recorded at nodes. T = ex-type cultures.
Ceratocystiopsis
Phylogenetic inferences were constructed using ITS (627 positions), βT (407 positions), TEF1-α (572 positions), and the concatenated datasets (ITS + βT + TEF1-α, 1606 positions). Our four representative isolates formed two distinct, well-supported terminal clades based on the ITS + βT + TEF1-α phylogenetic tree (Fig. 3), which are interpreted as two distinct taxa, each as representing an undescribed taxon. The two representative strains of Taxon 1 are most closely related to the C. weihaiensis R.L. Chang & X.Y. Zhang strains of the known species, while the two representative strains of Taxon 2 are closely related to the C. yantaiensis R.L. Chang & X.Y. Zhang strains. Both βT- and TEF1-α-based phylogenies robustly resolved Taxon 1 and Taxon 2 as distinct from related species (Suppl. materials 4, 5).
*Maximum Likelihood tree of Ceratocystiopsis generated from the combined (ITS + βT + TEF1-α) sequence data. Sequences generated in this study are printed in bold. Bold branches indicate posterior probability values ≥ 0.9, while ML SH-aLRT/UFBooT values of ≥ 75% are recorded at nodes, < 75% are indicated by the symbol . T = ex-type cultures.
Graphilbum
Phylogenetic inferences were constructed using ITS (621 positions), TEF1-α (543 positions), CAL (604 positions), and the concatenated datasets (ITS + TEF1-α + CAL, 2102 positions). Individual gene trees and the concatenated phylogeny congruently elucidated interspecific relationships within the genus (Fig. 4, Suppl. materials 6–8). Based on the ITS + TEF1-α + CAL phylogenetic tree, four representative strains were categorized into two distinct branches (Fig. 4). Taxon 3 represents Gr. translucens R.L. Chang & X.Y. Zhang, which were grouped in an independent clade with Gr. acuminatum R. Jankowiak & H. Solheim. Whereas Taxon 5 could not be equated to a known species, which is most closely related to the known species Gr. niveum R.L. Chang & X.Y. Zhang.
*Maximum Likelihood tree of Graphilbum generated from the combined (ITS + TEF1-α + CAL) sequence data. Sequences generated in this study are printed in bold. Bold branches indicate posterior probability values ≥ 0.9, while ML SH-aLRT/UFBooT values of ≥ 75% are recorded at nodes, < 75% are indicated by the symbol . T = ex-type cultures.
Hawksworthiomyces
In Hawksworthiomyces, the concatenated datasets (ITS + TEF1-α + RPB2, 1768 positions) were used to construct the phylogenetic inferences. The two representative strains of Taxon 5 were clustered with the type strains of H.w. taylorii Z.W. de Beer, Marincowitz & M.J. Wingfield and had a high support rate (Fig. 5). Accordingly, that strain of the Taxon 5 was confirmed as H.w. taylorii.
*Maximum Likelihood tree of Hawksworthiomyces generated from the combined (ITS + TEF1-α + RPB2) sequence data. Sequences generated in this study are printed in bold. Bold branches indicate posterior probability values ≥ 0.9, while ML SH-aLRT/UFBooT values of ≥ 75% are recorded at nodes, < 75% are indicated by the symbol . T = ex-type cultures.
Leptographium
The phylogenetic tree constructed using LSU sequences (Fig. 1) indicated that the representative strains within Leptographium were divided into three assemblage species. In Leptographium, the concatenated datasets (βT + TEF1-α, 778 positions) was used to construct phylogenetic inference. The results showed that our nine representative isolates were nested into four robust clades (Fig. 6). Phylogenetic analysis of this combined dataset confirmed that Taxon 6, together with southeastern Australian strains (VPRI22395 and VPRI43530) (Trollip et al. 2021), clustered within the same subclade of Grosmannia huntii (Rob.-Jeffr.) Zipfel, Z.W. de Beer & M.J. Wingf., demonstrating their high similarity with strong statistical support (SH-aLRT/UFBoot = 99.7/100). Taxon 7 was situated within the L. lundbergii complexes, which represents L. koreanum J.J. Kim & G.H. Kim, grouped with L. pinicola (K. Jacobs & M.J. Wingf.) Z.W. de Beer in a distinct lineage. Taxon 8 is positioned within the L. galeiforme complex. The βT + TEF1-α phylogeny resolved the three isolates of Taxon 8 form a monophyletic group with strong support (SH-aLRT/UFBoot = 92.4/93) alongside Australian strains (VPRI43839) representing L. radiaticola (J.J.Kim, Seifert & G.H.Kim) M.Procter & Z.W.de Beer, confirming their high degree of similarity and clustering in southeast Australia strains. The concatenated datasets phylogenetic analysis reveals that Taxon 9 forms a distinct, well-supported clade within the L. olivaceum complex, representing a novel, undescribed species. Taxon 9 formed a clade with L. rhizoidum M.L. Yin, Z.W. de Beer & M.J. Wingf. and L. sagmatosporum (E.F. Wright & Cain) M.L. Yin, Z.W. de Beer & M.J. Wingf. Phylogenetic analysis of the L. olivaceum complex was performed based on the βT (277 positions), TEF1-α (582 positions), and CAL (412 positions) datasets. The βT and CAL tree revealed clearer phylogenetic relationships among the species in the L. olivaceum complex than in the TEF1-α trees (Suppl. materials 9–11).
*Maximum Likelihood tree of Leptographium generated from the combined (βT + TEF1-α) sequence data. Sequences generated in this study are printed in bold. Bold branches indicate posterior probability values ≥ 0.9, while ML SH-aLRT/UFBooT values of ≥ 75% are recorded at nodes, < 75% are indicated by the symbol . T = ex-type cultures.
Masuyamyces and Ophiostoma
ips complex
In Masuyamyces and Ophiostoma ips complex, the ITS (461 positions) and βT (277 positions) datasets were used to construct the phylogenetic inferences. The results showed that our nine representative isolates nested into four robust clades (Fig. 7, Suppl. material 12), well-supported terminal clades, representing O. ips (Rumb.) Nannf. (Taxon 13), M. pallidulus (Linnakoski, Z.W. de Beer & M.J. Wingf.) Z.W. de Beer & M. Procter (Taxon 10) and two undescribed taxa (Taxa 11 and 12). Apart from O. ips, M. pallidulus and two undescribed taxa belonged to Masuyamyces. Our three isolates of O. ips and two isolates of M. pallidulus all formed a poorly supported subclade, somewhat divergent from the other strain. Taxa 11 and 12 were sister species, forming a well-supported clade (Fig. 7, Suppl. material 12).
*Maximum Likelihood tree of Masuyamyces and Ophiostoma ips complex generated from the ITS sequence data. Sequences generated in this study are printed in bold. Bold branches indicate posterior probability values ≥ 0.9, while ML SH-aLRT/UFBooT values of ≥ 75% are recorded at nodes, < 75% are indicated by the symbol . T = ex-type cultures.
Taxonomy
In this study, 13 phylogenetic species were identified, of which six were confirmed to represent different terminal branches, including two phylogenetic species of Ceratocystiopsis, one phylogenetic species of Graphilbum, one phylogenetic species of Leptographium and two phylogenetic species of Masuyamyces. They are interpreted as new species of Ophiostomatales.
Ceratocystiopsis
pseudoweihaiensis
Taxon classificationAnimaliaOphiostomatalesOphiostomataceae
D. Xie, H. W. Chen & D. F. Chi sp. nov.
4AA08C99-586B-5AA3-805E-4771CD3F9623
856956
Etymology.
The epithet pseudoweihaiensis (Latin) refers to the morphological resemblance and phylogenetic affinities with Ceratocystiopsis weihaiensis.
Diagnosis.
Ceratocystiopsis pseudoweihaiensis is phylogenetically distinct from all morphologically similar species, from which it can be readily distinguished using molecular sequence data for the beta-tubulin (βT) and the elongation factor 1-alpha (TEF1-α) regions (Suppl. materials 4, 5).
Type.
China • Shandong Province: Yantai City, from Hylurgus ligniperda, Nov. 2022, Dan Xie (holotypeHMAS 354192, dried culture prepared from NFF1617; ex-holotype culture CGMCC3.28600 = NFF1617).
Description.
Sexual morph not observed. Asexual morph observed mononematous. Mononematous morph: hyalorhinocladiella-like, conidiogenous cells arising directly from mycelium, (3.5–)6–52(–87) × (0.5–)1–1.5(–2) μm; conidia hyaline, smooth, unicellular short oblong, with rounded ends or clavate, (1.5–)2–3(–3.5) × (0.5–)1–1.5(–2) µm.
Culture characteristics.
The colonies are white in color on 2%MEA, hyphae submerged in agar with aerial mycelium. The optimal temperature for growth is 25–30 °C, reaching 63 mm diam in 15 days. Slow growth at 5 °C and 35 °C, growth diameter of 11 mm and 15 mm in 15 days.
Morphological characteristics of Ceratocystiopsis pseudoweihaiensis sp. nov. (NFF1617). a. Fifteen-day-old culture on 2% MEA; b–e. Hyalorhinocladiella-like asexual morph: conidiogenous cells and conidia. Scale bars: 20 μm (b–e).
Ecology.
Isolated from migratory beetles living in Pinus hosts. Insect vector: H. ligniperda.
Distribution.
Currently known only from Shandong Province, China.
Additional specimens examined.
China • Shandong Province: Yantai City, from H. ligniperda, Nov. 2022, D. Xie (living culture NFF1616).
Notes.
Phylogenetic analyses showed that C. pseudoweihaiensis is a close relative of C. weihaiensis. However, several morphological differences separate them. Ceratocystiopsis weihaiensis produces light brown colonies, while those of C. pseudoweihaiensis are white. The key distinguishing feature is the length of the conidiogenous cells, which are significantly longer in C. pseudoweihaiensis (6–52 μm) than in C. weihaiensis (10.9–29.2 μm) (Chang et al. 2021). Based on both phylogenetic and morphological evidence, we propose C. pseudoweihaiensis as a novel species.
Ceratocystiopsis
pseudoyantaiensis
Taxon classificationAnimaliaOphiostomatalesOphiostomataceae
D. Xie, H. W. Chen & D. F. Chi sp. nov.
36A7FFF5-F80D-547E-9F8D-0764AB136E16
856957
Etymology.
The epithet pseudoyantaiensis (Latin) refers to the morphological resemblance and phylogenetic affinities with Ceratocystiopsis yantaiensis.
Diagnosis.
Ceratocystiopsis pseudoyantaiensis is phylogenetically distinct from all morphologically similar species, from which it can be readily distinguished using molecular sequence data for the beta-tubulin (βT) and the elongation factor 1-alpha (TEF1-α) regions (Suppl. materials 4, 5).
Type.
China • Shandong Province: Yantai City, from H. ligniperda, Nov. 2022, Dan Xie (holotypeHMAS 354193, dried culture prepared from NFF1618; ex-holotype culture CGMCC3.28601 = NFF1618).
Morphological characteristics of Ceratocystiopsis pseudoyantaiensis sp. nov. (NFF1618). a. Fifteen-day-old culture on 2% MEA; b, c. Pesotum-like asexual morph; d–f. Hyalorhinocladiella-like asexual morph: conidiogenous cells and conidia. Scale bars: 100 μm (b); 20 μm (d–f).
Description.
Sexual morph not observed. Asexual morphs observed both synnematous and mononematous. Synnematous morph: pesotum-like, (47–)61–183(–283) μm long including conidiogenous apparatus, curving, faint yellow, the base dark yellow, aggregating into a mucilaginous spore drop, (36–)35–95(–165) μm in width. Mononematous morph: hyalorhinocladiella-like, conidiogenous cells arising directly from mycelium, (2.5–)4–24(–42) × (0.5–)1–1.5(–2) µm; conidia hyaline, smooth, unicellular, short oblong, measuring (1.5–)2–2.5(–3) × (0.5–)1–1.5(–2) μm.
Culture characteristics.
The colonies are faint yellow in color on 2%MEA, hyphae submerged in agar with aerial mycelium. The optimal temperature for growth is 30 °C, reaching 79 mm diam in 15 days. Growth slower at 5 °C, 14 mm diam in 15 days.
Ecology.
Isolated from migratory beetles living in Pinus hosts. Insect vector: H. ligniperda.
Distribution.
Currently known only from Shandong Province, China.
Additional specimens examined.
China • Shandong Province: Yantai City, from H. ligniperda, Nov. 2022, D. Xie (living culture NFF1619).
Notes.
Phylogenetic analyses showed that C. pseudoyantaiensis is phylogenetically closely related to C. yantaiensis. Morphologically, C. pseudoyantaiensis differs from C. yantaiensis in its conidiogenous cells; those of C. yantaiensis are branched, while those of C. pseudoyantaiensis are not (Chang et al. 2021). Based on both phylogenetic and morphological evidences, we propose the recognition of C. pseudoyantaiensis as a novel species.
Graphilbum
jiuguanense
Taxon classificationAnimaliaOphiostomatalesOphiostomataceae
D. Xie, H. W. Chen & D. F. Chi sp. nov.
40AC1594-7EB3-5448-A795-B2D1E7F5A7E6
856955
Etymology.
The epithet jiuguanense (Latin) refers to the Jiu Guan village from where this taxon was first isolated.
Diagnosis.
Graphilbum jiuguanense is phylogenetically distinct from all morphologically similar species, from which it can be readily distinguished using molecular sequence data for the ITS, the elongation factor 1-alpha (TEF1-α) and the calmodulin (CAL) regions (Suppl. materials 6–8).
Type.
China • Shandong Province: Yantai City, from H. ligniperda, Nov. 2022, Dan Xie (holotypeHMAS 354190, dried culture prepared from NFF1626; ex-holotype culture CGMCC3.28602 = NFF1626).
Description.
Sexual morph not observed. Asexual morphs observed both synnematous and mononematous. Synnematous morph: pesotum-like, (109–)179–309(–358) μm long including conidiogenous apparatus, curving, the base dark yellow, (32–)42–80(–141) μm in width. Conidiogenous cells hyaline, (14–)18–44(–53) × (1–)2–2.5(–3) μm. Conidia hyaline, one-celled, cylindrical to obovoid, (3–)4–5.5(–6) × (1–)1.5–2(–3) μm. Mononematous morph: hyalorhinocladiella-like, conidiogenous cells arising directly from mycelium, (9–)24–57(–92) × (0.5–)1–1.5(–2) µm; conidia hyaline, single-celled, smooth, ellipsoidal to ovoid, (2.5–)3–5(–6) × (1–)1.5–2(–2.5) μm.
Morphological characteristics of Graphilbum jiuguanense sp. nov. (NFF1626). a. Ten-day-old culture on 2% MEA; b, c. Pesotum-like asexual morph; d–g. Hyalorhinocladiella-like asexual morph: conidiogenous cells and conidia. Scale bars: 100 μm (b); 10 μm (c–e); 20 μm (f, g).
Culture characteristics.
Colonies on 2% MEA medium fast growing in the dark, hyphae submerged in agar with aerial mycelium, mycelial growing disorderly, white. The optimal temperature for growth is 30 °C, reaching 86 mm diam in 7 days. Growth slower at 35 °C, 7 mm diam in 7 days.
Ecology.
Isolated from migratory beetles living in Pinus hosts. Insect vector: H. ligniperda.
Distribution.
Currently known only from Shandong Province, China.
Additional specimens examined.
China • Shandong Province: Yantai City, from H. ligniperda, Nov. 2022, D. Xie (living culture NFF1627).
Notes.
Phylogenetic analyses show that Gr. jiuguanense is phylogenetically close to Gr. niveum. Morphologically, the colony of Gr. niveum is light brown, while that of Gr. jiuguanense is white with a disorganized mycelial growth pattern. Furthermore, Gr. jiuguanense produces both pesotum-like and hyalorhinocladiella-like asexual morphs, whereas Gr. niveum exclusively exhibits the hyalorhinocladiella-like morph (Chang et al. 2021). Based on both phylogenetic and morphological evidence, we propose the recognition of Gr. jiuguanense as a novel species.
Leptographium
ligniperdae
Taxon classificationAnimaliaOphiostomatalesOphiostomataceae
D. Xie, H. W. Chen & D. F. Chi sp. nov.
7B556177-85EA-5C11-A927-8BD96DF738F4
856952
Etymology.
The epithet ligniperdae (Latin) refers to the insect of Hylurgus ligniperda, the bark beetle vector of this species.
Diagnosis.
Leptographium ligniperdae is phylogenetically distinct from all morphologically similar species, from which it can be readily distinguished using molecular sequence data for the beta-tubulin (βT), the elongation factor 1-alpha (TEF1-α), and the calmodulin regions (CAL) (Suppl. materials 9–11).
Type.
China • Shandong Province: Yantai City, from H. ligniperda infesting Pinus thunbergii, Apr.2022, Dan Xie (holotypeHMAS 354189, dried culture prepared from NFF1605; ex-holotype culture CGMCC3.28598 = NFF1605).
Description.
Sexual morph not observed. Asexual morphs observed both synnematous and mononematous. Synnematous morph: pesotum-like, (110–)200–310(–356) μm long including conidiogenous apparatus, the base dark brown, expanding branches at the apex, (22–)28–58(–78) μm in width. Conidiogenous cells hyaline, cylindrical, (7–)9–16(–19) × 1–2.5 μm. Conidia hyaline, one-celled, cylindrical to obovoid, (5–)6.5–8 (–10) × (2.5–)3–3.5(–4) μm. Mononematous morph: leptographium-like, arising directly from mycelium; conidiophores, simple to strongly branched, hyaline, (41–)43–127(–226) µm long; conidiogenous cells (6–)8–18(–26) μm long; conidia hyaline, single-celled, smooth, oblong, (3.4–)4.8–7.6(–9.4) × (1.3–)1.7–2.5(–3) μm.
Culture characteristics.
Colonies on 2% MEA medium fast growing in the dark, reaching 88 mm in diam. in 7 days at 25 °C, growth rate up to 15 mm/day at the fastest. Hyphae submerged in agar with aerial mycelium, dark-olivaceous to brown. Optimal growth temperature 25–30 °C, no growth observed at 5 °C and 35 °C.
Morphological characteristics of Leptographium ligniperdae sp. nov. (NFF1605). a. Ten-day-old culture on 2% MEA; b, c. Pesotum-like asexual morph; d–f. Leptographium-like asexual morph: conidiogenous cells and conidia. Scale bars: 200 μm (b); 20 μm (c–f).
Ecology.
Isolated from beetles found on Pinus hosts. Host trees: Pinus thunbergii and Pinus densiflora. Insect vector: H. ligniperda.
Distribution.
Currently known only from Shandong Province, China.
Additional specimens examined.
China • Shandong Province: Yantai City, from H. ligniperda infesting Pinus densiflora, Apr.2022, D. Xie (living culture NFF1606).
Notes.
Leptographium ligniperda belongs to the L. olivaceum species complex and is most closely related to L. hizoidum and L. sagmatosporum in phylogenetic analyses. Distinguishing between these closely related species based on morphology alone is challenging due to their considerable morphological similarities in conidial size and shape, conidiogenous apparatus structure, and recorded asexual morphs (Yin et al. 2019).
Masuyamyces
xishanensis
Taxon classificationAnimaliaOphiostomatalesOphiostomataceae
D. Xie, H. W. Chen & D. F. Chi sp. nov.
E4B61B90-3201-5557-862F-3DCA625A7128
856953
Etymology.
The epithet xishanensis (Latin) refers to the Xi Shan village from where this taxon was first isolated.
Diagnosis.
Masuyamyces xishanensis is phylogenetically distinct from all morphologically similar species, from which it can be readily distinguished using molecular sequence data for the ITS and the beta-tubulin (βT) (Fig. 7, Suppl. material 12).
Type.
China • Shandong Province: Yantai City, from H. ligniperda, Apr. 2022, Dan Xie (holotypeHMAS 354188, dried culture prepared from NFF1608; ex-holotype culture CGMCC3.28599 = NFF1608).
Description.
Sexual morph not observed. Asexual morphs observed both synnematous and mononematous. Synnematous morph: pesotum-like, the base transparent or light brown, (112–)158–296(–345) μm tall, including the conidiogenous apparatus, (35–)87–117(–172) μm wide. Conidiogenous cells hyaline, (5–)9–18(–22) × 1–1.5 μm. Conidia hyaline, one-celled, cylindrical, (2–)2.5–4(–5) × (1–)1.5–2(–2) μm. Mononematous morph: hyalorhinocladiella-like, conidiogenous cells arising directly from mycelium, (5–)7–39(–61) × (0.5–)1–1.5(–2) µm; conidia hyaline, single-celled, smooth, obovoid to oblong, (2–)2.5–3.5(–4) × (0.5–)1–1.5(–2) μm.
Culture characteristics.
Colonies on 2% MEA medium slow growing in the dark, reaching 72 mm in diam. in 15 days at 25 °C, growth rate up to 4.5 mm/day at the fastest, the colonies margin smooth, part of the mycelium grows in the agar, pure white. Optimal growth temperature 30 °C, no growth observed at 5 °C, slow growth at 10 °C.
Morphological characteristics of Masuyamyces xishanensis sp. nov. (NFF1608). a. Fifteen-day-old culture on 2% MEA; b, c. Pesotum-like asexual morph; d. Pesotum-like asexual morph: conidiogenous cells and conidia; e–g. Hyalorhinocladiella-like asexual morph: conidiogenous cells and conidia. Scale bars: 200 μm (b, c); 20 μm (d–g).
Ecology.
Isolated from migratory beetles living in Pinus hosts. Insect vector: H. ligniperda.
Distribution.
Currently known only from Shandong Province, China.
Additional specimens examined.
China • Shandong Province: Yantai City, from H. ligniperda, Apr.2022, D. Xie (living culture NFF1609).
Notes.
Masuymyces xishanensis is phylogenetically most closely related to M. dongshanensis. Morphologically, M. xishanensis differs from M. dongshanensis by the absence of ascomatal necks, a more conspicuous pesotum-like stage, and a darker basal pigmentation. Based on both phylogenetic and morphological evidence, we propose the recognition of M. xishanensis as a novel species.
Masuyamyces
dongshanensis
Taxon classificationAnimaliaOphiostomatalesOphiostomataceae
D. Xie, H. W. Chen & D. F. Chi sp. nov.
64C276C5-75EE-587D-9C1E-83EC036549F0
856954
Etymology.
The epithet dongshanensis (Latin) refers to the Dong Shan village from where this taxon was first isolated.
Diagnosis.
Masuyamyces dongshanensis is phylogenetically distinct from all morphologically similar species, from which it can be readily distinguished using molecular sequence data for the ITS and the beta-tubulin (βT) (Fig. 7, Suppl. material 12).
Type.
China • Shandong Province: Yantai City, from H. ligniperda, Apr. 2023, D. an Xie (holotypeHMAS 354191, dried culture prepared from NFF1645; ex-holotype culture CGMCC3.28603 = NFF1645).
Description.
Sexual morph perithecial. Perithecia appeared after 35 days of cultivation on sterilized wooden chips or 30 days of cultivation on 2%MEA, superficial or partly embedded, globose, black, the outer layer with hyphal ornamentation, (249–)367–606(–727) μm diam. Ascomatal necks black, straight or slightly curved, (410–)480–1038(–1849) µm long, (77–)107–161(–195) µm wide at base, (18–)22–41(–52) µm wide at the apex. In culture, necks sometimes 2–4 per ascoma. Asci and ascospores were not observed. Asexual morphs observed both synnematous and mononematous. Synnematous morph: pesotum-like, the base transparent or light yellow, aggregating into a transparent mucilaginous spore drop, (64–)96–218(–316) μm tall, including the conidiogenous apparatus, (40–)58–108(–149) μm wide. Mononematous morph: hyalorhinocladiella-like, conidiogenous cells arising directly from mycelium, (22–)23–165(–359) × (2–)2.5–3.5(–4) µm; conidia hyaline, single-celled, smooth, obovoid to bacilliform, (6–) 7–9(–10) × (2–)2.5–3 (–4) μm.
Culture characteristics.
Colonies on 2% MEA medium slow growing in the dark, reaching 72 mm in diam. in 15 days at 25 °C, growth rate up to 4.5 mm/day at the fastest, part of the mycelium grows in the agar, pure white. Optimal growth temperature 30 °C, slow growth at 5–10 °C.
Morphological characteristics of Masuyamyces dongshanensis sp. nov. (NFF1645). a. Fifteen-day-old culture on 2% MEA; b, c. Perithecium; d, e. Pesotum-like asexual morph; f–h. Hyalorhinocladiella-like asexual morph: conidiogenous cells and conidia. Scale bars: 200 μm (b, c); 100 μm (d, e); 30 μm (f); 20 μm (g, h).
Ecology.
Isolated from migratory beetles living in Pinus hosts. Insect vector: H. ligniperda.
Additional specimens examined.
China • Shandong Province: Yantai City, from H. ligniperda, Apr. 2023, D. Xie (living culture NFF1646).
Distribution.
Currently known only from Shandong Province, China.
Notes.
Masuymyces dongshanensis is phylogenetically most closely related to M. xishanensis. Morphologically, it can be distinguished from M. xishanensis by the presence of ascomatal necks and its distinctive pesotum-like asexual morph, which is transparent, mucilaginous, and aggregates into irregularly edged, chip-like masses. Based on both phylogenetic and morphological evidence, we propose the recognition of M. dongshanensis as a novel species.
Isolation and intercontinental comparison analysis of ophiostomatoid fungi from Hylurgus
ligniperda in China
In this study, ophiostomatoid fungi were isolated from gallery-derived and trap-collected adults of H. ligniperda. Among these, O. ips exhibited the highest isolation frequency (36.55%), followed by L. radiaticola (14.94%) and L. koreanum (11.72%) (Table 3). Trap-captured adults yielded the greatest diversity of ophiostomatoid fungi (12 species), whereas gallery-derived adults showed the highest proportion of isolates (53.3%). Notably, the fungal diversity associated with gallery adults was lower (only seven species), though common ophiostomatoid species were isolated at higher frequencies compared with trap-captured adults. Known species such as Gr. translucens, along with new species C. pseudoweihaiensis, C. pseudoyantaiensis, Gr. jiuguanense, M. xishanensis, and M. dongshanensis, were isolated only from trap-captured adults. In contrast, the new species L. ligniperdae was only obtained from gallery adults. Ophiostomatoid fungi isolated from both gallery-derived and trap-collected adults included H. taylorii, G. huntii, L. koreanum, L. radiaticola, M. pallidulus, and O. ips. Comparative analysis of isolation frequencies between host sources revealed that G. huntii (χ² = 7.403, p = 0.0065), L. koreanum (χ² = 6.911, p = 0.0086), and L. radiaticola (χ² = 19.388, p < 0.0001) were isolated significantly more frequently from gallery adults than from trap-captured adults. In contrast, no significant differences in isolation frequency were observed between the gallery-derived and trap-collected adults for H. taylorii (χ² = 0.013, p = 0.91), M. pallidulus (χ² = 0.893, p = 0.345), or O. ips (χ² = 0.129, p = 0.719) (Suppl. material 1).
Table 3.: Strains numbers and percentage of ophiostomatoid fungi associated with adult Hylurgus ligniperda in Shandong Province, China.
The current results were compared with extensive research on the global distribution of ophiostomatoid fungi associated with H. ligniperda across five continents (Asia, Africa, Europe, the Americas, and Oceania) (Fig. 14, Suppl. material 2, and references therein). Based on available data, 45 species have been identified in association with H. ligniperda. Documented records indicate that Europe exhibits the highest species diversity (15 species), followed by North America (14 species), South Africa (13 species), East Asia (13 species), Oceania (ten species), and South America (six species). Ophiostomatoid fungal communities show significant divergence among continents. Seven species associated with H. ligniperda exhibit transcontinental distributions (occurring on ≥ three continents). The numbers of endemic fungal species in each region are as follows: East Asia (nine species), Europe (eight species), North America (seven species), South Africa (six species), South America (three species), and Oceania (one species). Ophiostoma ips is the only species shared across all six regions. Ophiostoma quercus (Georgev.) Nannf. has been reported in North America, South Africa, Europe, and Oceania but was not isolated in the present study. Our isolation of G. huntii aligns with earlier records from North America and Oceania. Similarly, M. pallidulus identified in this study has also been documented in Europe and Oceania in association with this bark beetle.
Upset diagram showing overlaps of the ophiostomatoid fungal communities associated with Hylurgus ligniperda in East Asia, Americas, South Africa, Europe and Oceania. The left bar graph shows the number of ophiostomatoid fungal species identified in each continent. The horizontally aligned dots on the right correspond to their respective continental regions shown on the left. Vertically, these dots connect to the bars above, representing the quantities of both endemic and shared ophiostomatoid fungal species across continents. The connecting lines represent shared fungal species. The colors of the bars match those of the fungal names in the diagram.
Discussion
Currently, H. ligniperda has successfully colonized and caused significant damage to coastal shelter forests in the Yanwei region of Shandong Province, China. In this study, fungi were isolated from adults H. ligniperda obtained from galleries and traps. A total of 425 strains of fungi associated with longhorn beetles were isolated, marking the first investigation into ophiostomatoid communities associated with H. ligniperda in China. Although fungal isolation is influenced by various environmental factors, including individual insect variability and environmental contamination during the isolation process, this study employed extensive sampling and isolation techniques. It is anticipated that the findings will provide a reliable representation of the fungal communities of ophiostomatoid fungi associated with H. ligniperda in Yantai and Weihai, Shandong Province, China. Based on the phylogenetic analysis of multigene fragments and observation of microscopic characteristics, 13 species belonging to six genera were identified. They included seven known species, viz. Gr. translucens, H.w. taylorii, G. huntii, L. koreanum, L. radiaticola, M. pallidulus, and O. ips. And six previously undescribed species, viz. C. pseudoweihaiensis, C. pseudoyantaiensis, Gr. jiuguanense, L. ligniperdae, M. xishanensis, and M. dongshanensis.
The ophiostomatoid fungal communities associated with gallery-derived and trap-captured H. ligniperda adults were significantly different. The symbiotic relationships between bark beetles and ophiostomatoid fungi appear to change with changes in the environmental conditions in which the bark beetles reside. The higher number of ophiostomatoid fungi isolated from bark beetles captured in traps may be attributed to the beetles’ exposure to a broader range of environmental fungi at this stage, rather than solely harboring specific fungi. This finding is consistent with previous studies reporting differences in species composition and isolation frequency of fungi associated with bark beetles across different countries and regions (Taerum et al. 2013; Wang et al. 2020), indicating that beetles acquire and carry environmental fungi. Furthermore, studies have shown that the fungi carried by beetles not only influence nutrient acquisition but also assist in degrading tree defensive compounds, thereby affecting the fitness of the beetles (Ayres et al. 2000; Cale et al. 2016; Six and Elser 2019; Zhao et al. 2019). The limited diversity of ophiostomatoid fungi species observed within the galleries, coupled with a high frequency of isolation, suggests that these fungi may facilitate the colonization of bark beetles in the region.
Currently, O. ips is distributed worldwide and associated with various beetles that infest conifers, contributing to wood bluing and, in conjunction with bark beetles, weakening trees and accelerating forest decline (Raffa and Smalley 1988; Klepzig et al. 1991; Pastirčáková et al. 2018). The fungus has also been isolated from several bark beetle-associated mites in Yunnan, China (Chang et al. 2017). There is significant interest in the relationship between O. ips and Bursaphelenchus xylophilus (Steiner & Bührer) Nickle (Niu et al. 2012; Suh et al. 2013; Zhao et al. 2014; Wang et al. 2018). In this study, O. ips exhibited the highest isolation rate, aligning with findings from previous research conducted on ophiostomatoid fungi associated with pines infected by Cryphalus piceae Ratzeburg (Chang et al. 2021), Bursaphelenchus xylophilus and Monochamus alternatus Hope (Wang et al. 2018) in Shandong, China. Ophiostoma ips was isolated from bark beetles collected across various countries, with the frequency of isolation varying by region (Zhou et al. 2004c; Ray et al. 2006; Davydenko et al. 2014; De Errasti et al. 2018). Ophiostoma ips has been reported from H. ligniperda across all six continents and exhibited the highest isolation frequency in the present study. This pattern may be attributed to its broad global distribution combined with its status as the most frequently isolated ophiostomatoid fungus in Shandong Province, China.
The genus Leptographium accommodates L. radiaticola, L. koreanum, G. huntii, and L. ligniperdae, all of which showed significantly higher isolation frequencies from gallery-derived than trap-captured adults, suggesting their specialized adaptation to the gallery environment. Leptographium radiaticola was the second most abundant ophiostomatoid fungus isolated in this study. Initially it was described as being isolated from P. radiata D. Don imported to Korea (Kim et al. 2005a), but this is not the first time the species has been spotted. Leptographium radiaticola has been previously identified in isolates from South Africa, Chile, Sweden, and California (Zhou et al. 2001; Zhou et al. 2004b; Kim et al. 2011). This is not the first instance of L. radiaticola being recorded in China. It was initially isolated from the gallery of Dendroctonus valens on P. tabuliformis in Shanxi Province, China (Lu et al. 2009). Subsequently, it was isolated again in Yunnan Province, China, during a survey of ophiostomatoid fungi affecting coniferous beetles and beetle-associated mites (Chang et al. 2017). These two reports involved different beetles that harm various conifers. This study reports the first occurrence of L. radiaticola in Shandong Province, China. Additionally, the sequences obtained here were distinct from those of strains previously isolated in the Yunnan region of China (Chang et al. 2017). This fungus is widely distributed globally and is particularly associated with H. ligniperda. The strain isolated in South Africa was obtained from H. ligniperda (Zhou et al. 2001), while those in Chile were isolated from Hylastes ater and H. ligniperda galleries on P. radiata (Zhou et al. 2004c). In California, the fungus was isolated from H. ligniperda, which poses a threat to P. halepensis Mill. and P. pinea L. (Kim et al. 2011). Leptographium radiaticola was isolated from H. ligniperda, an exotic bark beetle, within pine plantations in Argentina. The fungus has also been reported from H. ligniperda on P. taeda L. and P. elliotti Engelm. (De Errasti et al. 2018). Furthermore, during a survey of the fungal community of ophiostomatoid fungi associated with pine and pine bark beetles in southeastern Australia, this fungus was isolated from H. ater and H. ligniperda, both of which are detrimental to P. radiata (Trollip et al. 2021). It is likely that L. radiaticola from the Shandong region of China was introduced alongside H. ligniperda. Research has revealed that multiple beetle species harbor stable symbiotic partners (Lewinsohn et al. 1994; Six and Bentz 2003), and such bark beetle–fungus associations contribute to enhancing host adaptability by assisting the beetles in overcoming tree defense mechanisms (Six 2020). The close association between H. ligniperda and L. radiaticola may be attributed to the fungus’s ability to facilitate the colonization and development of H. ligniperda. This is further supported by the observation that the separation frequency within the adults’ galleries was significantly higher than that of the adults in traps in this study, suggesting that H. ligniperda in the gallery requires more L. radiaticola to withstand the host.
Leptographium koreanum was isolated and described from P. densiflora and P. koraiensis Siebold & Zucc. infested by Tomicus piniperda Linnaeus in Korea (Kim et al. 2005b). In a comprehensive survey of ophiostomatoid fungi associated with Dendroctonus valens in eastern North America, western North America, and China, it was observed that L. koreanum is predominantly confined to Asia. However, this fungus was also isolated in eastern North America, albeit at a lower prevalence compared to China (Taerum et al. 2013). Pathogenicity tests demonstrated that L. koreanum could induce significant necrosis in the bark of P. tabuliformis (Lu et al. 2009). Following its invasion of the Yanwei region of China, H. ligniperda established contact with L. koreanum. Numerous invasive beetles establish novel associations with indigenous fungi at new invasion sites (Jacobs et al. 2003; Hausner et al. 2005), leading to the hypothesis that L. koreanum may facilitate the colonization of H. ligniperda. Grosmannia huntii, a group of fungi well-suited for transmission by bark beetles, exhibits a broad geographical distribution, encompassing regions such as Chile, New Zealand, Argentina, North America, and Australia(Zhou et al. 2004c; Reay et al. 2005; Ray et al. 2006; Taerum et al. 2013; De Errasti et al. 2017; Trollip et al. 2021). Notably, H. ligniperda infestations have been documented in these regions, although G. huntii has not yet been reported in China. The strains isolated in this study displayed high similarity to Australian strains obtained from H. Ater and H. ligniperda (Trollip et al. 2021). It is postulated that G. huntii is closely associated with H. ligniperda and may have been introduced to China alongside H. ligniperda, where it has successfully colonized. Leptographium ligniperdae is a member of the L. olivaceum species complex (De Beer et al. 2022). In this study, it was exclusively isolated from adult H. ligniperda within their galleries. Multigene phylogenetic analysis revealed that it is closely related to L. rhizoidum and L. sagmatosporum. Leptographium rhizoidum was isolated and characterized from H. ater and H. attenuatus Erichson, which infest P. radiata in Spain. Currently known insect vectors include H. ater, H. attenuatus, Hylurgops palliatus Gyllenhal, and Ips sexdentatus Börner (Yin et al. 2019). Leptographium sagmatosporum was initially isolated from bark beetle galleries and fresh wood surfaces on Picea mariana (Mill.) Britton, Sterns & Poggenb., P. resinosa Aiton, and P. strobus L. (Yin et al. 2019). The discovery of L. ligniperdae not only contributes to the diversity of the L. olivaceum species complex but also provides valuable insights into the ophiostomatoid fungal community associated with H. ligniperda.
Species of Masuyamyces isolated in this study include M. pallidulus and two novel species, M. xishanensis and M. dongshanensis (De Beer et al. 2022). In Finland, M. pallidulus was initially isolated from P. sylvestris, which is associated with various beetle species (Linnakoski et al. 2010). In Poland, M. pallidulus was isolated from the roots of dead trees and subsequently identified during a survey of ophiostomatoid fungi linked to root-eating beetles on P. sylvestris in the region (Jankowiak and Bilański 2013). Masuyamyces pallidulus is also present in Australia, including isolates from H. ligniperda (Trollip et al. 2021). In the genus Hawksworthiomyces, H.w. taylorii was first isolated from Eucalyptus poles within the soil layer in South Africa, with an optimal growth temperature of 30 °C (De Beer et al. 2016b). This fungus had not been previously isolated from H. ligniperda. Given that H. ligniperda primarily feeds on the roots of its host and interacts with the soil environment, it is hypothesized that the fungus is a soil-associated organism.
Certain fungi belonging to Ceratocystiopsis and Graphilbum were exclusively isolated from adults of H. ligniperda in traps. In the genus Ceratocystiopsis, C. pseudoweihaiensis is closely related to C. weihaiensis, while C. pseudoyantaiensis is closely related to C. yantaiensis. Both C. weihaiensis and C. yantaiensis have been isolated and reported from the Cryphalus piceae gallery on P. thunbergii in Shandong Province, China (Chang et al. 2021). Within the genus Graphilbum, Gr. translucens, and Gr. jiuguanense are identified. Multilocus phylogenetic analysis indicated that Gr. jiuguanense formed a distinct lineage closely related to Gr. niveum. Furthermore, both Gr. translucens and Gr. niveum were isolated from Cryphalus piceae galleries on P. densiflora and P. thunbergii in Shandong Province, China (Chang et al. 2021). This aligns with the ecological niche of H. ligniperda in China, suggesting that H. ligniperda acquires native fungi from the environment, although some fungi may undergo changes as they adapt to new host insects. The absence of these fungi in adult samples within the gallery may be attributed to their weak association with H. ligniperda.
The significant divergence in ophiostomatoid fungal communities observed across continents demonstrates that regional variation serves as the primary driver of fungal community differentiation in H. ligniperda associations. In this study, six novel fungal species along with several long-term fungal partners were isolated from H. ligniperda adults, indicating that the beetle develops specialized fungal assemblages adapted to local environments across different regions to facilitate its colonization and expansion into new habitats.
Conclusions
Our findings revealed a fungal community of ophiostomatoid fungi associated with H. ligniperda, which has successfully colonization in Yanwei, Shandong Province, China. We identified a total of 13 species across six genera, including six novel species. Ophiostoma ips and L. radiaticola emerged as the dominant species, with isolation frequencies of 36.55% and 14.94%, respectively. Variations in the ophiostomatoid communities associated with H. ligniperda across different regions may be attributed to environmental differences. Nevertheless, the presence of certain fungi, such as O. ips, L. radiaticola, G. huntii, and M. pallidulus, remains consistent. Notably, the L. radiaticola and G. huntii strains isolated in this study clustered into a single clade with high sequence similarity to strains from southeast Australia, also isolated from H. ligniperda. It is hypothesized that the source population of H. ligniperda established in Shandong, China, is the same as that of the Australian population. Furthermore, native fungi and newly identified species closely related to native fungi were isolated from H. ligniperda, suggesting that insects in newly invaded areas establish interactions with native fungi, which in turn adapt to these insects through evolutionary changes. Understanding this novel association formed by H. ligniperda in the Yantai and Weihai regions of China is crucial for future management strategies of this insect. The successful colonization of alien invasive insects in new environments is influenced by numerous factors, among which the fungi associated with bark beetles play a significant role. Consequently, it is imperative to comprehend the relationship between bark beetles and their associated fungi, as this knowledge will facilitate timely control of alien invasive insect populations and help prevent substantial economic losses.
Supplementary Material
XML Treatment for Ceratocystiopsis pseudoweihaiensis
XML Treatment for Ceratocystiopsis pseudoyantaiensis
XML Treatment for Graphilbum jiuguanense
XML Treatment for Leptographium ligniperdae
XML Treatment for Masuyamyces xishanensis
XML Treatment for Masuyamyces dongshanensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ayres MP Wilkens RT (2000) Nitrogen budgets of phloem-feeding bark beetles with and without symbiotic fungi. Ecology 81: 2198–2210. 10.1890/0012-9658(2000)081[2198:NBOPFB]2.0.CO;2. · doi ↗
- 2Bedoya CL Brockerhoff EG (2019) Acoustic communication of the red-haired bark beetle Hylurgus ligniperda.Physiological Entomology 44: 252–265. 10.1111/phen.12301 · doi ↗
- 3Bentz BJ Six DL (2006) Ergosterol content of fungi associated with Dendroctonus ponderosae and Dendroctonus rufipennis (Coleoptera: Curculionidae, Scolytinae). Annals of the Entomological Society of America 99: 189–194. 10.1603/0013-8746(2006)099[0189:ECOFAW]2.0.CO;2 · doi ↗
- 4Biedermann PHWMüller J (2019) Bark beetle population dynamics in the anthropocene: challenges and solutions.Trends in Ecology & Evolution 34: 914–924. 10.1016/j.tree.2019.06.00231262532 · doi ↗ · pubmed ↗
- 5Biedermann PHW Vega FE (2020) Ecology and evolution of insect–fungus mutualisms.Annual Review of Entomology 65: 431–455. 10.1146/annurev-ento-011019-02491031610133 · doi ↗ · pubmed ↗
- 6Cale JA Collignon RM (2016) Fungal volatiles can act as carbon sources and semiochemicals to mediate interspecific interactions among bark beetle-associated fungal symbionts. P Lo S ONE 11: e 0162197. 10.1371/journal.pone.0162197 PMC 500877027583519 · doi ↗ · pubmed ↗
- 7Chang R Duong TA (2017) Ophiostomatoid fungi associated with conifer-infesting beetles and their phoretic mites in Yunnan, China.Myco Keys 28: 19–64. 10.3897/mycokeys.28.21758 PMC 580414029559821 · doi ↗ · pubmed ↗
- 8Chang R Zhang X (2021) Ophiostomatoid species associated with pine trees (Pinus spp.) infested by Cryphalus piceae from eastern China, including five new species.Myco Keys 83: 181–208. 10.3897/mycokeys.83.7092534720643 PMC 8528803 · doi ↗ · pubmed ↗