Resistance, virulence and genetic diversity of Salmonella Typhimurium in South Africa (1999–2021)
Nkagiseng Moatshe, Emmanuel Seakamela, Khanyisile R. Mbatha, Linda A. Bester, Nombasa Ntushelo, Itumeleng Matle

TL;DR
This study analyzed antibiotic resistance, virulence genes, and genetic diversity of Salmonella Typhimurium in South Africa from 1999 to 2021, revealing high resistance and diverse genetic profiles.
Contribution
The study provides a comprehensive analysis of resistance patterns, virulence genes, and plasmid diversity in S. Typhimurium isolates from multiple sources in South Africa.
Findings
High resistance levels to antibiotics like ciprofloxacin, ceftriaxone, and piperacillin were observed in S. Typhimurium isolates.
Class 1 integrons and resistance genes such as blaPSE, blaCMY-2, tetA, and qnrB were commonly detected.
Genetic diversity was identified through ERIC-PCR, revealing 44 clusters with six major ones.
Abstract
Salmonella Typhimurium is a major cause of human and animal salmonellosis, impacting global socioeconomic factors. This study examined antibiotic resistance patterns, virulence genes and plasmids in S. Typhimurium isolates from nine South African provinces (1999–2021). Among 180 randomly selected isolates, 129 were confirmed as S. Typhimurium from animal (n = 94), food (n = 16), environment (n = 7) and feed (n = 12) sources using polymerase chain reaction (PCR). Phenotypic resistance was assessed against 13 antibiotics, revealing high resistance levels: 86.8% to ciprofloxacin, 69.0% to ceftriaxone and 65.1% to piperacillin. Multidrug resistance (up to 11 antibiotics) was observed. Genetic analysis identified resistance genes, including blaPSE (32.6%), blaCMY-2 (21.7%), tetA (24.0%), tetB (22.5%), qnrB (21.7%) and qnrA (20.2%). Class 1 integrons were found in 47.4% of isolates. Virulence…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
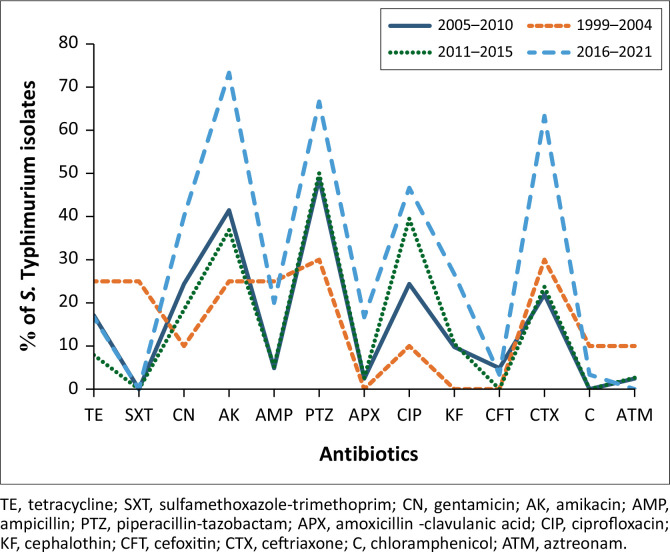 FIGURE 1
FIGURE 1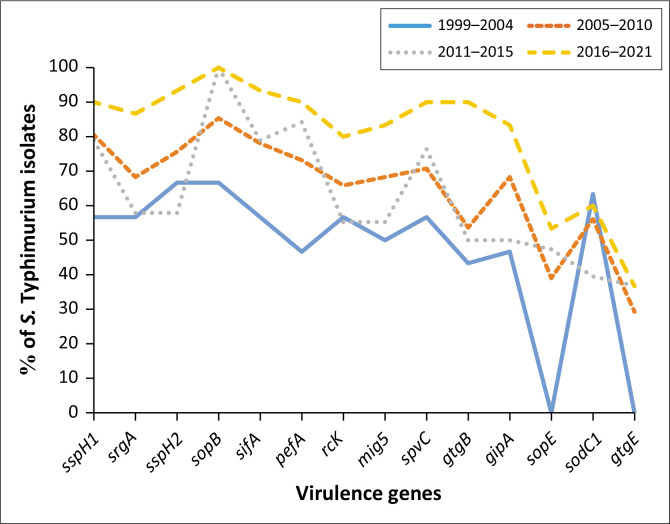 FIGURE 2
FIGURE 2 FIGURE 3
FIGURE 3| Primer | Primer sequence (5’ to 3’) | Size (bp) |
|---|---|---|
| STM0716F | AACCGCTGCTTAATCCTGATGG | 187 |
| STM0716R | TGGCCCTGAGCCAGCTTTT | |
| STM1350F | TCAAAATTACCGGGCGCA | 171 |
| STM1350R | TTTTAAGACTACATACGCGCATGAA | |
| STM0839F | TCCAGTATGAAACAGGCAACGTGT | 137 |
| STM0839R | GCGACGCATTGTTCGATTGAT | |
| STM4525F | TGGCGGCAGAAGCGATG | 114 |
| STM4525R | CTTCATTCAGCAACTGACGCTGAG | |
| STM4538F | TGGTCACCGCGCGTGAT | 93 |
| STM4538R | CGAACGCCAGGTTCATTTGT | |
| STM2150F | CATAACCCGCCTCGACCTCAT | 101 |
| STM2150R | AGATGTCGTGAGAAGCGGTGG |
| Antibiotic class | Gene | Primer sequence (5’ to 3’) | Size (bp) | Melting temperature (oC) | Reference |
|---|---|---|---|---|---|
| Tetracycline |
| F:GGCGGTCTTCTTCATCATCATGC | 502 | 59 | Pavelquesi et al. ( |
|
| F:CGCCCAGTGCTGTTGTTGTC | 173 | |||
| Trimethoprim |
| F:CGGTCGTAACACGTTCAAGT | 220 | 55 | Matayoshi et al. ( |
|
| F:AAATTCCGGGTGAGCAGAAG | 429 | |||
|
| F:GCAGTCGCCCTAAAACAACG | 294 | |||
| Sulphonamides |
| F:CGGCGTGGGCTACCTGAACG | 433 | 63 | Pavelquesi et al. ( |
|
| F:GCGCTCAAGGCAGATGGCATT | 293 | |||
|
| F:CAACGGAAGTGGGCGTTGTGGA | 244 | |||
| Phenicol |
| F:CTTGTCGCCTTGCGTATAAT | 508 | 53 | Odoch et al. ( |
|
| F:CTGAGGGTGTCGTCATCTAC | 673 | |||
|
| F:CGCCACGGTGTTGTTGTTAT | 394 | |||
| β-lactams |
| F:TTAACTGGCGAACTACTTAC | 247 | 55 | Karolina et al. ( |
|
| F:GACAGCCTCTTTCTCCACA | 1000 | |||
|
| F:AGGATTGACTGCCTTTTTG | 393 | |||
|
| F:TGCTTCGCAACTATGCTAC | 438 | |||
| Quinolones |
| F:TCAGCAAGAGGATTTCTCA | 516 | 53 | Takaichi et al. ( |
|
| F:GATCGTGAAAGCCAGAAAGG | 469 | |||
|
| F:ACGACATTCGTCAACTGCAA | 417 |
| Target gene | Primer | Primer sequence (5’ to 3’) | PCR conditions | Number of cycles | Size (bp) | Reference |
|---|---|---|---|---|---|---|
|
| Int1F | GCCTTGCTGTTCTTCTACGG | 94 °C for 5 min | 35 | 558 | Ramatla et al. ( |
|
| Int2F | CACGGATATGCGACAAAAAGG | 94 °C for 5 min, | 32 | 740 | |
|
| Int3F | GCCTCCGGCAGCGACTTTCAG | 94 °C for 10 min, | 35 | 650 | Rowe-Magnus and Mazel ( |
| Gene | Melting temperature (°C) | Size (bp) | Primer sequence (5’ to 3’) | References |
|---|---|---|---|---|
|
| 64 | 220 | F-GGACCGGCCAGCAACAAAACAAAGAAGAAG | Skyberg, Logue and Nolan ( |
|
| 57.5 | 436 | F – TGCACGGGGAAAACTACTTC | Chiu and Ou ( |
|
| 244 | F – ACAGTGCTCGTTTACGACCTGAAT | ||
|
| 60 | 246 | F – TGCAGAAAAAGGGGAATACG | Capuano et al. ( |
|
| 57.5 | 362 | F – CGAGTAAAGACCCCGCATAC | Capuano et al. ( |
|
| 570 | F – ACTCCTTGCACAACCAAATGCGGA | ||
|
| 66.5 | 157 | F – GCGCCGCTCAGCCGAACCAG | Skyberg et al. ( |
|
| 449 | F- TTTGCCGAAGAACGCGCCCCCCACACG | ||
|
| 58 | 212 | F – GCAAGCTGTACATGGCAAAG | Capuano et al. ( |
|
| 50 | 467 | F – TATTGTCGCTGGTAGCTG | Capuano et al. ( |
|
| 1113 | F – AGGAGGAGTGTAAAGGT | Ntivuguruzwa ( | |
|
| 58 | 248 | F – AACCAACCAGACCAACCTTC | Capuano et al. ( |
|
| 189 | F – AACGGACGGAACACAGAGTC | Ntivuguruzwa ( | |
|
| 58 | 203 | F – GCACAACTGGCTGAAGATGA | Capuano et al. ( |
|
| 344 | F – TGTTCCGGTCATAATGCAGA |
| Antibiotic class | Name of antibiotics | Interpretation in % |
| |
|---|---|---|---|---|
| Sensitive | Resistant | |||
| Penicillins or β-lactams | Ampicillin (AMP) | 75.9 | 24.1 | < 0.0001 |
| Piperacillin (TZP) | 34.9 | 65.1 | < 0.0001 | |
| Amoxycillin (AMC) | 83.7 | 16.3 | < 0.0001 | |
| Cephalosporins | Cephalothin (KF) | 48.8 | 51.2 | < 0.0001 |
| Cefoxitin (FOX) | 86.0 | 14.0 | < 0.0001 | |
| Ceftriaxone (CRO) | 31.0 | 69.0 | 0.0658 | |
| Aminoglycoside | Gentamycin (GN) | 53.5 | 46.5 | < 0.0001 |
| Amikacin (AK) | 38.8 | 61.2 | 0.0001 | |
| Tetracyclines | Tetracycline (TE) | 69.0 | 31.0 | < 0.0001 |
| Fluoroquinolones | Ciprofloxacin (CIP) | 13.2 | 86.8 | < 0.0001 |
| Monobactams | Aztreonam (ATM) | 76.7 | 23.3 | < 0.0001 |
| Sulphonamides | Trimethoprim (STX) | 87.6 | 12.4 | < 0.0001 |
| Phenicols | Chloramphenicol (CHL) | 70.5 | 29.5 | < 0.0001 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSalmonella and Campylobacter epidemiology · Vibrio bacteria research studies · Antibiotic Resistance in Bacteria
Introduction
Salmonella Typhimurium is a Gram-negative, facultative anaerobic bacterium belonging to the family Enterobacteriaceae. It is a leading serovar implicated in salmonellosis, a zoonotic disease of considerable public health and economic importance globally (Galán-Relaño et al. 2023). Alongside S. Enteritidis, S. Typhimurium is one of the most frequently reported serotypes in South Africa, contributing significantly to the global burden of foodborne illnesses, with estimated 93.8 million human cases annually (Majowicz et al. 2010; Ramatla et al. 2022).
In humans, S. Typhimurium infection typically results from consuming contaminated food products derived from animals, particularly poultry, eggs and dairy. The bacterium can also be transmitted via the faecal–oral route (Mkangara 2023). Salmonella Typhimurium infection in humans often manifests as self-limiting gastroenteritis characterised by diarrhoea, abdominal pain, vomiting and fever, but in immunocompromised individuals or those with co-morbidities, it can lead to severe invasive disease, such as bacteraemia or systemic septicaemia (Wright et al. 2005). The invasive non-typhoidal S. Typhimurium strains (iNTS), particularly those prevalent in sub-Saharan Africa, pose heightened risks because of their ability to cause life-threatening systemic disease in vulnerable populations, including infants and human immunodeficiency virus (HIV)-positive individuals (Hajra, Nair & Chakravortty 2023; Okoro et al. 2012).
In animals, S. Typhimurium infections present diverse clinical manifestations ranging from asymptomatic carriage to severe systemic disease, such as septicaemia or enteritis. These infections pose challenges to animal health, agricultural productivity and food safety, with livestock serving as both reservoirs and amplifiers of the pathogen (Hoelzer, Moreno Switt & Wiedmann 2011). The adaptability of S. Typhimurium to environmental stressors and host immune responses is underpinned by complex regulatory systems governing gene expression, enabling survival and replication across diverse ecological niches (Ilyas, Tsai & Coombes 2017).
The pathogenicity of S. Typhimurium is mediated by numerous virulence factors encoded on the chromosomal pathogenicity islands (SPIs) and plasmids. SPI-1 facilitates host cell invasion through a type III secretion system (T3SS), delivering effector proteins that induce cytoskeletal rearrangements and membrane ruffling in epithelial cells (Lou et al. 2019). SPI-2, another critical virulence determinant, enables intracellular survival and replication within macrophages, protecting the bacterium from immune-mediated clearance (Ramatla et al. 2020). In addition, the 90-kb pSLT virulence plasmid encodes genes such as spvRABCD, which enhance systemic infection, and rck and pef, which confer resistance to complement-mediated killing and promote adhesion, respectively (Rotger & Casadesús 1999; Silva, Puente & Calva 2017).
The emergence and dissemination of multidrug resistant (MDR) S. Typhimurium strains have exacerbated the global public health challenge, complicating treatment options. Resistance is often associated with mobile genetic elements such as plasmids, transposons and integrons. Class 1 integrons, commonly identified in S. Typhimurium, harbour gene cassettes encoding resistance to multiple antibiotics, including sulphonamides (sul1, sul2), chloramphenicol (cat1, cat2) and tetracyclines (tetA, tetB) (Gillings 2014; Wang et al. 2014). Resistance to critical drugs such as β-lactams (blaCTX-M, blaTEM-1, blaSHV) and fluoroquinolones (qnrA, qnrB) further complicates management strategies (Adesiji et al. 2014; Eguale et al. 2017). The global spread of MDR S. Typhimurium phage type DT104, known for its resistance to at least five antibiotics, underscores the urgency of addressing antimicrobial resistance (Wang et al. 2019; Threlfall 2002).
Despite the significant burden of S. Typhimurium infections, data on its prevalence, epidemiology and economic impact in sub-Saharan Africa remain fragmented. In South Africa, the lack of integrated surveillance systems limits comprehensive understanding and control of this pathogen in both human and animal populations (Mthembu, Zishiri & El Zowalaty 2019). Surveillance efforts rely heavily on laboratory reports from abattoirs, diseased animals and feed monitoring, which provide valuable insights but fail to capture the full scope of its impact (Kidanemariam et al. 2010; Khumalo & Mbanga 2014). The overuse and misuse of antibiotics in livestock exacerbate the dissemination of resistant strains, posing a dual threat to public health and food security by affecting livestock productivity and the safety of animal-derived products.
Therefore, the aim of this study was to investigate the antibiotic resistance patterns, virulence gene profiles and genetic diversity of S. Typhimurium isolated from the environment, animals and food products in South Africa over a 22-year period (1999–2021). By analysing isolates from diverse sources, the study sought to elucidate the prevalence of antimicrobial resistance, characterise virulence determinants and assess the genetic relatedness of strains, contributing to a deeper understanding of the epidemiology of S. Typhimurium and its implications for public health, food safety and antimicrobial stewardship in the region.
Research methods and design
Study design and isolates selection
The study was a retrospective cohort analysis based on laboratory-confirmed isolates of S. Typhimurium collected from diverse sources across South Africa between 1999 and 2021. These isolates were initially recovered from samples submitted for diagnostic purposes and preserved by freeze-drying, ensuring long-term viability. Samples were stored at the General Bacteriology Laboratory of the Onderstepoort Veterinary Research, South Africa. Based on the simple random sampling without replacement, a total of 180 isolates of S. Typhimurium from (1) various geographical locations in the country, (2) different sources of isolation (animal, animal products, feed and environmental samples) and (3) animal species (livestock, companion animals, wildlife) were included in this study. To prepare the isolates for further analysis, each freeze-dried sample was carefully revived. The isolates were inoculated into a brain heart infusion (BHI) broth and incubated at 37 °C for 18 h – 24 h to restore bacterial viability and ensure optimal growth conditions for downstream testing.
Deoxyribonucleic acid extraction and polymerase chain reaction confirmation of S. Typhimurium
The genomic deoxyribonucleic acid (DNA) was extracted from culture using the boiling method as described by Karimnasab et al. (2013). Isolates were confirmed using a multiplex polymerase chain reaction (PCR) as described by Kim et al. (2006). In brief, a 25 µL-PCR mixture comprising 10.5 µL of Taq 2× Master Mix RED (Ampliquor, Denmark), 0.5 µL (5 pmol/µL) of each primer (Inqaba Biotechnical Industries (Pty) Ltd., South Africa) (Table 1), 4.5 µL UltraPure DNase/RNase-Free Distilled Water (Thermo Fisher Scientific, United States [US]) and 4 µL DNA template. The thermocycler (A 9700 Applied Biosystems, US) was set as follows: initial denaturation at 94 °C for 5 min, 40 cycles of denaturation at 94 °C for 30 s, annealing temperature 62 °C for 30 s, extension at 72 °C for 1 min and final extension at 72 °C for 3 min. The PCR amplicons were analysed by electrophoresis on a 1.5% agarose gel containing 4 µL ethidium bromide using ultraviolet (UV) light and photographed (Omega Fluor, Aplegen). Salmonella Typhimurium (ATCC 14028) strain and ribonuclease/deoxyribonuclease (RNase/DNase) free water were used as a positive and negative controls, respectively.
Antimicrobial susceptibility testing
All isolates were subjected to antimicrobial susceptibility test using Kirby Bauer disk diffusion method and interpreted according to the European Committee on Antimicrobial Susceptibility Testing (EUCAST 2021). The isolates were tested against 13 antibiotics (Table 5) (Thermo Fisher Scientific, United Kingdom [UK]). These antibiotics were selected based on the consultation with veterinarians, the World Health Organization (WHO) ranking of antimicrobials in human medicine as well as the availability of EUCAST antibiotic breakpoints. In brief, the overnight pure cultureson nutrient agar were inoculated into sterile saline and diluted to the equivalent concentration of 0.5 McFarland standard. The bacterial suspension was inoculated aseptically onto Mueller–Hinton agar plates (Thermo Fisher Scientific, UK) and allowed to stand for 5 min. Antibiotic discs were then placed per inoculated plate followed by incubation at 37 °C for 24 h. The antibiotic susceptibility of each bacterial isolate was reported as sensitive or resistant according to the zone diameters described in the EUCAST (2021).
Determination of antibiotic-resistant genes
Various multiplex PCR were performed for the screening of 18 resistance genes (Table 2). The 25 µL-PCR reaction mixture comprised 12.5 µL of Taq 2× Master Mix RED (Ampliquor, Denmark), 0.512 µL (5 pmol/µL) of each primer (Inqaba Biotechnical Industries (Pty) Ltd., South Africa), 4.5 µL (with the exception β-lactams 3.5 µL and tetracycline 5.5 µL) UltraPure DNase/RNase-Free Distilled Water (Thermo Fisher Scientific, US) and 5 µL DNA template. The thermocycler (Applied Biosystems, US) was set as follows: initial denaturation at 94 °C for 3 min, 30 cycles of denaturation at 94 °C for 30 s, annealing temperature variable (Table 2) for 30 s, extension at 72 °C for 1 min and final extension at 72 °C for 10 min. The PCR amplicons were analysed by electrophoresis on a 2.5% agarose gel containing 4 µL ethidium bromide using UV light and photographed (Omega Fluor, Aplegen).
Molecular detection of integrons
Isolates were screened for int1, int2 and int3 genes carried by class 1, 2 and 3 integrons, respectively using PCR. Briefly, 25 µL PCR reactions used comprised 12.5 µL of Taq 2× Master Mix RED (Ampliquor, Denmark), 2 µL (10 mM) of each primer (Inqaba Biotechnical Industries (Pty) Ltd., South Africa), 4.5 µL UltraPure DNase/RNase-Free Distilled Water (Thermo Fisher Scientific, US) and 4 µL DNA template. A 9700 PCR machine (Applied Biosystems, US) was used for amplification using PCR conditions (Table 3). The PCR products were electrophoresed in 1.5% agarose gel containing 4 µL ethidium bromide at 120 V for 60 min and photographed under UV light (Omega Fluor, Aplegen).
Determination of virulence genes
A screening for 15 virulence genes was also performed on all S. Typhimurium isolates (Table 4). The 25 µL PCR reaction mixture comprised 12.5 µL of Taq 2× Master Mix RED (Ampliquor, Denmark), 0.5 µL (5 pmoL/µL) of each primer (Inqaba Biotechnical Industries (Pty) Ltd., South Africa), 4.5 µL (with exception sopB, gipA and sspH1 6.5 µL) UltraPure DNase/RNase-Free Distilled Water (Thermo Fisher Scientific, US) and 4 µL DNA template. Amplification was carried out as described by Ntivuguruzwa (2016). In brief, a thermocycler (9700 Applied Biosystems, US) was set as follows: initial denaturation at 94 °C for 3 min, 30 cycles of denaturation at 94 °C for 30 s, annealing temperature variable (Table 4) for 1 min, extension at 72 °C for 1 min and final extension at 72 °C for 5 min. PCR amplicons were analysed by electrophoresis on a 1.5% agarose gel containing 4 µL ethidium bromide using UV light and photographed (Omega Fluor, Aplegen).
Detection of plasmids of Salmonella Typhimurium
Plasmid DNA was extracted using the ZymoPURE plasmid midiprep kit (Zymo Research, US) as per manufacturer’s instructions. Extracted plasmid DNA was run on a 0.7% agarose gel (Thermo Fisher Scientific, US) at 120 V for 1 h and visualised under UV light and photographed using an Omega Fluor gel documentation system (Omega Fluor, Aplegen). The Inqaba Biotechnologies (Pretoria, South Africa) extended 1 kb ladder and the E. coli 0157:H7 were used as the DNA marker and positive control, respectively.
ERIC polymerase chain reaction
The ERIC PCR method was performed as described by Almeida et al. (2016). Primers (Inqaba Biotechnical Industries (Pty) Ltd., Johannesburg, South Africa): ERIC-R: ATG AAG CTC CTG GGG ATT CAC and ERIC-F: AAG TAA GTG ACT GGG GTG AGC G were used. In brief, the 25 µl PCR reaction mixture comprised 12.5 µL of Taq 2× Master Mix RED (Ampliquor, Denmark), 2 µL (5 pmoL/µL) of each primer (Inqaba Biotechnical Industries (Pty) Ltd., South Africa), 4.5 µL UltraPure DNase/RNase-Free Distilled Water (Thermo Fisher Scientific, US) and 4 µL DNA template. A thermocycler (9700 Applied Biosystems, US) was set as follows: initial denaturation at 94 ^o^C for 5 min, followed by 40 cycles of denaturation at 94 ^o^C for 60 s, annealing at 40 ^o^C for 90 s and extension at 72 ^o^C for 60 s with a single cycle of final extension at 72 ^o^C for 7 min. The PCR products were electrophoresed in 1.5% agarose gel containing 4 µL ethidium bromide at 120 V for 60 min and images were captured using Omega Fluor gel documentation systems (Omega Fluor, Aplegen). A 1 kb-plus DNA ladder (Biolabs, New England, UK) and S. Typhimurium (accession number: SRX10785603) were used as the DNA marker and a positive control, respectively.
Data analysis
The data were subjected to a Chi-square test using the Frequency Procedure (PROC FREQ) of Statistical Analysis System (SAS) statistical software version 9.4 (Clark 2004). The following formula was used to calculate:
where A is the total number of resistance determinants recorded in the population, N is the number of isolates in the population and Y is the total number of antibiotics tested (Andriyanov et al. 2021). ERIC PCR dendrogram and analysis were analysed using BioNumerics software 6.6 (Applied Maths NV, Belgium) using the Dice coefficient and the unweighted pair group (UPGMA) with arithmetic averages using 1% tolerance and 0.5% optimisation settings to analyse electrophoretic patterns. Clustering was identified using a similarity cut-off of 70%. For better analysis, the years in this study were categorised into four periods spanning 5 years: 1999–2004; 2005–2010; 2011–2015; 2016–2021.
Results
Confirmation
A total of 180 isolates were initially selected for this study, of which only 92.0% (n = 140) proved to be viable on culture media. Of the 140 (92.1%) isolates, 129 were confirmed as S. Typhimurium using PCR, while 11 isolates were identified as other Salmonella species. Among the confirmed isolates, 72.9% (n = 94) were isolated from animal (bovine, caprine, equine, ovine, porcine, poultry) samples while 12.4% (n =16), 5.4% (n = 7) and 9.3% (n = 12) were from food, the environment and feed samples, respectively.
Antimicrobial susceptibility test and antibiotic resistance index
A total of 129 isolates were tested against 13 antimicrobial agents and showed high resistance to ciprofloxacin (n = 112; 86.8%), ceftriaxone (n = 89; 69.0%), piperacillin (n = 84; 65.1%), amikacin (n = 79; 61.2%), cephalothin (n = 66; 51.2%), gentamycin (n = 60; 46.5%) and tetracycline (n = 40; 31.0%) (Table 5). The antibiotic resistance results showed statistical significance (p < 0.0001) with the exception of ceftriaxone (p = 0.0658). The Antibiotic Resistance Index (ARI) of all tested isolates was less than 0.2. This indicates a relatively low level of antibiotic resistance, suggesting that most of the S. Typhimurium isolates were exposed to limited selective pressure from antibiotics in their respective environments.
Antimicrobial resistance trends over the years
Antibiotic resistance by year showed that during the 1999–2004 period, piperacillin–tazobactam (30%) and ceftriaxone (30%) ranked higher followed by tetracycline, sulfamethoxazole-trimethoprim, amikacin and ampicillin with 25% each, while no resistance was observed in amoxicillin–clavulanic acid, cephalothin and cefoxitin. The year 2005–2010 was dominated by piperacillin–tazobactam (49%), amikacin (41%), gentamycin (24%), ciprofloxacin (24%) and ceftriaxone (22%) (Figure 1). During the third period (2011–2015), the highest resistance was observed in piperacillin–tazobactam (50%), ciprofloxacin (39%) and amikacin (37%) while the fourth period (2016–2021) was dominated by amikacin (73%), piperacillin–tazobactam (67%), ceftriaxone (63%) and ciprofloxacin (47%). Overall, sulfamethoxazole–trimethoprim, cefoxitin, chloramphenicol and aztreonam remained sensitive (≤ 5%) throughout the years while gentamycin and ciprofloxacin showed an increase from 10% to 47% in the period 1994–2001. Tetracycline remained constant throughout the years while resistance in amikacin, and piperacillin–tazobactam and ceftriaxone showed an increase from 25% to over 60% in 2021 (Figure 1).
Antibiotic resistance trend of Salmonella Typhimurium from 1999 to 2021.
Virulence genes trends over the years
Figure 2 shows that during the period under investigation, all virulence genes, except sopE and gtgE were detected in over 40% – 67% in the first period (1999–2004) and increased steadily to over 80% in the fourth period (2016–2021).
Virulence genes trend in Salmonella Typhimurium isolates from 1999 to 2021.
Antibiotic resistance patterns and multi-resistant isolates of Salmonella Typhimurium
Among the 129 isolates tested, 23 antimicrobial resistance patterns were observed. Resistance to three or more antimicrobials was seen across all species tested in this study. Only one isolate was resistant to 11 antimicrobials (TE, SXT, AK, AMC, CIP, KF, AMP, FOX, CHL, TZP and GN). At least 7.4% of porcine and 12.5% ovine isolates showed resistance to seven (AK–GN–TZP–CIP–FOX–KF–CRO) antibiotics. Resistance to the pattern AK–GN–FOX–KF–CIP–TZP (six antibiotics) was observed in 4.7% of poultry and 18.5% porcine isolates, while resistance to five antibiotics (GN–AK–TZP–CIP–KF) was observed in the following species: bovine (30.8%), poultry (13.9%), equine (11.1%) and other (25.0%). It was observed that 12.5%, 7.7%, 4.7% of isolates from ovine, bovine and poultry, respectively were resistant to four antimicrobials (TE–GN–AK–TZP). Resistance to three antibiotics was seen in the following species: bovine (11.5%), poultry (4.7%), porcine (14.8%), caprine (50.0%) and ovine (12.5%). Resistance to two antibiotics yielded three patterns: pattern 1 (TZP-CIP) – porcine (3.7%), caprine (25.0%), equine (11.1%), ovine (12.5%) and other (16.7%). Pattern 2 (AK-TZP) – porcine (14.8%), equine (11.1%) and ovine (12.5%). Pattern 3 (CIP-CRO) – bovine (11.5%), poultry (20.9%), porcine (14.8%) and equine (22.2%).
Presence of antimicrobial resistant genes in Salmonella Typhimurium isolates
A total of 129 isolates were screened for the presence of 18 resistance genes. The blaPSE gene was detected in 32.6% (n = 42) while blaCMY-2, blaTEM and blaSHV genes were present in 21.7% (n = 28), 18.6% (n = 24) and 17.5% (n = 23) of the isolates, respectively. Furthermore, isolates harboured 18.6 % (n = 24), 7.8% (n = 10) and 6.2% (n = 8) of sul1, sul2 and sul3 genes, respectively. The qnrA gene was detected in 20.2% (n = 26), followed by qnrB (21.7%; n = 28) and qnrS (10.9%; n = 14) while tetA and tetB were detected in 24.0% (n = 31) and 22.5% (n = 29) of the isolates, respectively. Among the trimethoprim encoding genes, resistance was detected in dfrXI (18.6%; n = 24) and dfrXII (15.5 %; n = 20), while no resistance was detected for dfrxIII (0.0%).
Presence of integrons in Salmonella Typhimurium isolates
In this study, isolates were screened for class 1 (int1), 2 (int2) and 3 (int3) integrons with int1 gene detected in 47.3% (n = 61) of the isolates, followed by int2 (36.4%; n = 47) and int3 (24.8%; n = 32) (class 3) encoding for the int3 gene. Class 2 (p = 0.0021) and class 3 (p-value = 0.0001) results were considered statistically significant.
Presence of virulence genes among Salmonella Typhimurium
Among the 129 S. Typhimurium isolates investigated in this study, InvA (100%; n =129) was predominant, followed by sopB gene (95.3%; n = 123) sspH1 (82.9%, n =107), sifA (82.9%; n =107), pefA (79.8%; n = 103), spvC (79.1%; n = 102), sspH2 (77.5%; n =100) and srgA (71.3%; n =92). Furthermore, the mig5 and rcK genes were both detected in 69.0% while gipA, gtgB, sopE, sodC1 and gtgE were detected in 66.7%, 62.8%, 60.5%, 58.1% and 19.4%, respectively.
Presence of plasmids in Salmonella Typhimurium isolates
Conducting plasmid extraction from 129 S. Typhimurium isolates, revealed the presence of a diverse range of plasmid sizes from 2 kb to 90 kb. Remarkably, among these isolates, a significant majority, constituting 71.3% (n = 92), were found to possess the substantial 90 kb plasmid. In contrast, only a small fraction, 2.3% (n = 3) of the isolates, carried the diminutive 2 kb plasmid, while a noteworthy 31.0% (n = 40) of isolates contained the intermediate-sized 8 kb plasmid.
ERIC polymerase chain reaction
The DNA fingerprinting of 129 isolates was done using ERIC PCR to determine the relationship of isolates (Figure 3). Furthermore, isolates were grouped into clusters based on a cluster index of 70% and clonal relation was determined at a similarity index of 90%. The isolates were categorised into 44 different ERIC types (A – RR), with six major clusters being identified: F (n = 8), J (n = 10), L (n = 16), R (n = 9), W (n =1 4) and X (n = 7). Clonal relatedness (three or more isolates) was observed among the isolates in clusters F, J, L, R, W and CC. It was also noted that 80% (n = 8) of isolates from cluster J were from animal sources (porcine [n = 3], bovine [n = 1], ovine [n = 1] and poultry [n = 3]) and meat (n = 2). In cluster L, 75% (n = 12), 12.5% (n = 2) and 12.5% (n = 2) isolates were from animals [porcine (n = 4), bovine (n = 4), equine (n =1) and poultry (n = 3)], meat and feed, respectively.
A dendrogram illustrating the genetic similarity, source, sample type and year of S. Typhimurium isolates. The ERIC PCR cluster grouping is based on a cluster index of 70% and similarity index of 90%.A dendrogram illustrating the genetic similarity, source, sample type and year of S. Typhimurium isolates. The ERIC PCR cluster grouping is based on a cluster index of 70% and similarity index of 90%.A dendrogram illustrating the genetic similarity, source, sample type and year of S. Typhimurium isolates. The ERIC PCR cluster grouping is based on a cluster index of 70% and similarity index of 90%.
Cluster W had 42.9% (n = 6) isolates from animals (equine [n =1], bovine [n = 2], ovine [n = 1] and poultry [n = 2]), 35.7% (n = 5) from meat and 21.4% (n = 3) from feed. Among these isolates, 50% (n = 7) were isolated from Gauteng province; moreover, 50% (n = 7) isolates were clonally related. Cluster F consisted of 62.5% isolates originating (n = 5) from animals (porcine [n = 1], bovine [n = 3] and poultry [n = 1], 12.5% [n = 1] from environment, 12.5% [n = 1] from feed and 12.5% [n = 1]) from meat and X consisting of 71.4% isolates (n = 5) from animals (porcine [n = 1], bovine [n = 1], equine [n = 1] and poultry [n = 2]), 14.3% (n = 1) meat and 14.3% (n = 1) environment. Cluster R consisted of 77.8% (n = 7) of isolates taken from animals (poultry [n = 3], porcine [n = 2] and ovine [n = 2]). Furthermore, in cluster R, two isolates indicated 100% similarity and were both from KwaZulu-Natal (Figure 3).
Discussion
In the current study, 12% – 87% of isolates showed resistance to 13 antibiotics, with the highest proportion towards ciprofloxacin (86.8%), ceftriaxone (69.0%), piperacillin (65.1%), amikacin (61.2%), cephalothin (51.2%), gentamycin (46.5%) and tetracycline (31.0%). Salmonellosis caused by S. Typhimurium is self-limiting in healthy individuals. However, systemic infections in young children, the elderly and immunocompromised individuals require antimicrobial treatment (Nazir et al. 2025). The recommended regimen for Salmonella includes third-generation cephalosporins, quinolones and macrolides (Collignon et al. 2016).
Resistance towards ciprofloxacin in this study was observed in 86.8% of the isolates. Ciprofloxacin is an antibiotic of choice for the treatment of invasive Salmonella infections in adults (Parry & Threlfall 2008). The result of the current study exceeds the 64%, 19.5% and 0% reported in China, Iran and Turkey, respectively (Guo et al. 2023; Moghadam et al. 2023; Şik & Akan 2024). However, the result of our study was lower than the result (100%) reported by Siddiky et al. (2024) in Bangladesh.
Sixty-nine per cent and 51.6% of the isolates showed resistance towards ceftriaxone and cephalothin, respectively. Cephalothin, a first-generation cephalosporin is known to treat serious infections caused by both Gram positive and negative bacteria including Salmonella while ceftriaxone is an empirical choice for the treatment of Salmonella infections in children (Shi et al. 2021). Furthermore, ceftriaxone is a preferred antibiotic for invasive infections when bacteria are resistant to ciprofloxacin (Arizpe et al. 2016). The resistance towards cephalothin in our study was found to be higher than 0% reported in Malaysia by Adzitey, Rusul and Huda (2012), but lower than 75.7% reported by Elshebrawy et al. (2022) from Egypt, while the high and low resistance towards ceftriaxone in comparison to our study was reported from Nigeria (100.0%) and Bangladesh (13.8%), respectively (Igbinosa et al. 2023; Rahman et al. 2024).
Resistance to piperacillin was recorded in 50.4% of the isolates. Consequently, the presence of piperacillin-resistant S. Typhimurium in non-human sources raises the possibility of a hypothetical transmission from human strains to non-human origins. The current results were found to be lower than the 100% rate reported in S. Typhimurium isolates by Igbinosa et al. (2023). The disparity in study results can be attributed to variations in antibiotic usage patterns influenced by geographical location. However, Moghadam et al. (2023) reported results (58.3%) similar to this study in Iran.
In terms of aminoglycosides, resistance was observed in amikacin (61.2%) and gentamycin (46.5%). Amikacin and gentamycin are critically important antibiotics used to treat enterococcal endocarditis and MDR tuberculosis (WHO 2007). Resistance to these antibiotics may be fuelled by their use in treating animal infections. The resistance in our study towards amikacin and gentamycin was lower than those reported in a study from Italy which reported 100.0% resistance towards each antibiotic while studies in Egypt and Bangladesh reported 10.8% and 13.3%, respectively (Elshebrawy et al. 2022; Siddiky et al. 2024).
In this study, 31.0% of isolates were resistant to tetracycline. Tetracycline is a broad-spectrum antibacterial agent used in both human and veterinary medicine to treat and prevent Salmonella infections (Lugo-Melchor et al. 2010). The use of tetracycline to promote growth in animals resulted in an increase in resistant isolates. However, high levels of resistance to tetracycline were reported in studies from Iran: 72.2% by Moghadam et al. (2023), Bangladesh: 86.6% by Siddiky et al. (2021) and Malaysia: 78.4% by Adzitey et al. (2012). The results of the current study are in correlation with those reported in Egypt: 32.4% (Elshebrawy et al. 2022). The extensive use of tetracycline worldwide has resulted in the development of resistance among Salmonella spp. (Pavelquesi et al. 2021).
The high resistance towards critically and highly important antimicrobials in this study is of great concern as it may impair treatment efficacy (Ahmed et al. 2024). Furthermore, antimicrobial resistance in Salmonella is associated with horizontal gene transfer which may influence the pathogenic characteristics of Salmonella strains in the future (Collignon et al. 2016). It is worth noting that the use of antibiotics as growth promoters has been banned in the European Union in 2006 (Henton et al. 2011). However, the use of antibiotics for animal production in African countries including South Africa continues to date. Although there is a move on firm awareness on the usage of antibiotics in animal production in South Africa, stringent measures need to be taken to remedy this situation.
During the period under investigation, the antibiotic and virulence trends of isolates in our study showed low proportion in the early 2000s with steady increase in both the resistance and virulence which was as expected. Our results were consistent with the previous studies that reported the resistance of Salmonella species to commonly used antibiotics including tetracyclines even beyond 1999; hence most of the isolates were already showing resistance towards most of the antibiotics tested (Ali et al. 2025; Hur, Jawale & Lee 2012; Nonga et al. 2010). Furthermore, our study supports the understanding that antimicrobial resistance is a complex process that is fuelled by many factors including the misuse and overuse of antibiotics in both human and veterinary medicine. Therefore, prioritising antimicrobial stewardship is essential.
One isolate from poultry was resistant to 11 antibiotics (TE–SXT–AK–AMC–CIP–KF–AMP–FOX–CHL–TZP–GN). Siddiky et al. (2021) found one S. Typhimurium isolate to be resistant to 12 antibiotics which exceeded our results. However, a study in Italy found one isolate to be resistant to 10 antibiotics (Lauteri et al., 2022). The notable rise in resistance to critical antibiotics such as ciprofloxacin, third-generation cephalosporins and aminoglycosides emphasises the urgent need for prudent antibiotic use and effective surveillance to address antibiotic resistance effectively (O’neil 2016). In addition, these results highlight the significance of ongoing monitoring of antibiotic susceptibility to guide optimal treatment plans and public health interventions (WHO 2017). Isolates in this study were MDR, with an increase over the years.
Resistance to β-lactams in S. enterica is mainly because of the production of acquired β-lactamases. In the current study, blaPSE and blaTEM, which are frequently associated with the ampicillin and amoxicillin/clavulanate resistance were detected in 32.6% and 18.6% while blaCMY-2, and blaSHV genes which code for the resistance of third-generation cephalosporins and penicillins were present in 21.7% and 17.5%, respectively (De Toro et al. 2011; Zhao et al. 2009). Zhao et al. (2017) and Oh et al. (2016) reported 12.5% and 1.1% blaPSE detection, respectively, which were lower than our results. The blaTEM results in our study were lower than those reported by studies in Pakistan (100%) and Bangladesh (64.8%) (Fatima et al. 2023; Rahman et al. 2024). In this study, the blaCMY-2 detection was higher than those reported by Thong and Modarressi (2011) in Malaysia while the blaSHV was reported in 12.5% of the isolates by Rahman et al. (2024). The presence of bla genes suggest that the S. Typhimurium isolates encoding these genes can effectively resist β-lactams. Furthermore, these genes can be transferred to other isolates.
In the current study, sul1, sul2 and sul3 genes, which are responsible for conferring resistance to sulphonamides, were detected in 18.6 %, 7.8% and 6.2% of the isolates, respectively. These results were consistent with the phenotypic resistance profiles. In a study conducted in Iran, sul1, sul2 and sul3 were detected in 84%, 50% and 17%, respectively, which were higher than the results of our study (Moghadam et al. 2023).
Tetracycline-encoding tetA and tetB genes were detected in 24.0% and 22.5% of Salmonella isolates, respectively, aligning with the observed phenotypic resistance patterns. These genes are responsible for encoding efflux pumps that actively expel tetracycline antibiotics from bacterial cells, thereby lowering the intracellular concentration of the antibiotic and diminishing its efficacy (Møller et al., 2016; Pavelquesi et al. 2021). It was observed that the occurrence of tetA and tetB genes in Salmonella is a consequence of intricate interactions involving various factors, such as antibiotic utilisation, bacterial genetics and environmental conditions (Mthembu et al. 2021). For instance, Moghadam et al. (2023) reported notably varying detection rates of 72% for tetA and 23% for tetB genes in tetracycline-resistant Salmonella Typhimurium isolates in Iran. Lauteri et al. (2022) assessed antimicrobial resistance in Salmonella Typhimurium strains isolated from Italian swine food chain, with tetA and tetB, genes identified in 26.3% and 73.7% of these isolates, respectively.
The emergence of plasmid-mediated fluoroquinolone resistance in Salmonella is driven by the presence of qnrA, qnrB and qnrS genes. In the current study, it was observed that 21.7%, 20.2% and 10.8% of S. Typhimurium isolates carried the qnrB, qnrA and qnrS genes, respectively. These findings contrast with those reported by Zhao et al. (2017) who detected qnrA and qnrB in 81.3% of the isolates for each. However, Zhao et al. (2017) and Pribul et al. (2017) observed a qnrS detection of 3.1% and 3.6%, respectively which was lower than our results. Importantly, the occurrence of qnrA, qnrB or qnrS genes in Salmonella strains can vary significantly depending on geographic location and Salmonella serotype. Despite the presence of these genes associated with quinolone and fluoroquinolone resistance in this study, the phenotypic results indicated a high level of resistance to ciprofloxacin. This discrepancy might be attributed to the absence of an efficient promoter region, or the possibility of these genes conferring greater resistance or susceptibility to ciprofloxacin. However, the presence of qnrA, qnrB and qnrS genes in our isolates is of concern, especially as quinolones are approved for therapeutic and preventive use in animals in South Africa (Eagar, Swan & Van Vuuren 2012). Therefore, their misuse could contribute to the emergence of antibiotic-resistant bacteria in humans (Henton et al. 2011).
In this study, the presence of sifA (82.9%) and sopB (95.3%) were detected within S. Typhimurium isolates which agrees with the findings of other studies (Hughes et al. 2008; Skyberg et al. 2006). These genes play pivotal roles in the invasion of macrophages, with sifA being particularly responsible for the pathogen’s survival, contributing to Salmonella infection (Ibarra & Steele-Mortimer 2009).
Plasmid encoding virulence genes (mig5, rcK, srgA, spvC and pefA) were detected in a range of 69.0% – 79.8% of the isolated strain. These findings were higher than results reported by Proroga et al. (2019) and Capuano et al. (2013) in Italy. The spv gene encodes the main components for S. Typhimurium plasmid-mediated virulence and is carried into host cells T3SS2 (Browne et al. 2008; Ibarra & Steele-Mortimer 2009). These genes (rck, SrgA, SpvC, PefA) are involved in infection stages such as adhesion, invasion, adaptation, intracellular survival within host macrophages and also the evasion of host immune responses (Ahmer, Tran & Heffron 1999; Kendall & Sperandio 2014; Koczerka et al. 2021; Long et al. 2022).
Bacteriophages encoding virulence genes gtgB and sodC1 were detected in 71.4% and 68.6% of the isolated strains, respectively, while 19.3% of the isolates carried gtgE. These findings diverge slightly from those reported by Capuano et al. (2013), where gtgE was absent in all the isolates. These genes (gtgE, gtgB and sodC1) facilitate Salmonella’s survival within a host and enhance its pathogenicity, as described by Foley et al. (2013), making Salmonella infections challenging to effectively treat.
In this study, the presence of the Gifsy-1 encoded gene gipA, which aids Salmonella in invading host cells, was detected in 66.7% of the isolates (Nguyen Thi et al. 2020). These findings align with the observations made by Capuano et al. (2013) and Proroga et al. (2019) in Italy. However, current results are lower than the 1.43% reported by Sharma et al. (2019) in chickens. In addition, this study revealed the presence of the sspH1 gene in 82.9% of the S. Typhimurium isolates, which is higher compared to the findings reported by Capuano et al. (2013) but consistent with the results reported by Long et al. (2022). The elevated prevalence of these virulence genes in our study underscores the potential of the isolated S. Typhimurium strains to cause infections.
The 90 kb plasmid known as the virulence plasmid that primarily functions to boost *Salmonella’*s growth during the systemic phase of the illness was recorded in 71.3% of isolates in this study (Gulig & Doyle 1993). This plasmid is capable of mobilisation but lacks conjugative abilities (Ahmer et al. 1999). These results were lower than the 44.0% (n = 16/46) reported by Sameshima et al. (2000) in Japan and 48.5% reported by Benacer et al. (2010) in Malaysia. However, no isolates were reported to carry this plasmid by Guerra et al. (2002) in Spain. In a study by Khasa, Singh and Sidhu (2018) in India, 100% of S. Typhimurium isolates carried the 90 kb plasmid.
The ERIC PCR fingerprinting has been shown to provide increased discrimination and is fast, simple and cheap (Kumar, Surendran & Thampuran 2008). Using ERIC PCR, the S. Typhimurium isolates were differentiated and grouped into 44 clusters (A-RR). In this study, Cluster F consisted of isolates originating from feed, meat, environment and animals and cluster X consisted of isolates originating from animals, environment and meat. This could indicate a possible cross-contamination among the different sources of isolation although these isolates were collected from different provinces. All isolates in cluster J originated from animals and were widely distributed between 2001 and 2021. Understanding the source of Salmonella in the food chain is crucial for minimising occurrences in humans (Magwedere et al. 2015). The clonal relatedness among isolates (50%) in cluster W suggests that isolates from different provinces in South Africa are related. Cluster L consisted of isolates from animals and feed from Gauteng province. These findings suggest that there might be a cross-contamination between animals and feed in Gauteng province. In cluster R, a pair of clones were reported from KwaZulu-Natal in different species. Findings in this study suggest that S. Typhimurium is widely distributed and this is in agreement with studies by Gelaw, Nthaba and Matle (2018) and Magwedere et al. (2015).
These findings highlight the urgent need for robust antimicrobial stewardship programmes and stringent regulatory measures to control antibiotic usage in both health care and agricultural practices. In addition, continuous surveillance and monitoring of antimicrobial resistance patterns are critical to inform policy decisions and implement effective interventions aimed at curbing the spread of resistant S. Typhimurium strains. Addressing this issue is paramount to safeguarding public health and ensuring the continued efficacy of vital antimicrobial therapies.
Conclusion
Salmonella Typhimurium remains a significant global foodborne pathogen with profound implications for both public and animal health. This study has demonstrated that S. Typhimurium isolates in South Africa exhibit resistance to critically and highly important antibiotics including ciprofloxacin, which is a cornerstone of treatment for invasive salmonellosis. In addition, most isolates were found to harbour virulence factors associated with adhesion, invasion and intracellular survival. The combination of antimicrobial resistance (AMR) and pathogenicity factors highlights the dual threat posed by these isolates, making them a critical public health risk. Therefore, the authors recommend the implementation of stringent antimicrobial stewardship policies in both human and veterinary medicine as well as robust surveillance programmes to monitor the epidemiology of S. Typhimurium and other foodborne pathogens.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adesiji, Y.O., Deekshit, V.K. & Karunasagar, I., 2014, ‘Antimicrobial-resistant genes associated with Salmonella spp. isolated from human, poultry, and seafood sources’, Food Science and Nutrition 2(4), 436–442. 10.1002/fsn 3.11925473501 PMC 4221842 · doi ↗ · pubmed ↗
- 2Adzitey, F., Rusul, G. & Huda, N., 2012, ‘Prevalence and antibiotic resistance of Salmonella serovars in ducks, duck rearing and processing environments in Penang, Malaysia’, Food Research International 45(2), 947–952. 10.1016/j.foodres.2011.02.05122285201 · doi ↗ · pubmed ↗
- 3Ahmed, S.K., Hussein, S., Qurbani, K., Ibrahim, R.H., Fareeq, A., Mahmood, K.A. et al., 2024, ‘Antimicrobial resistance: Impacts, challenges, and future prospects’, Journal of Medicine Surgery and Public Health 2, 100081. 10.1016/j.glmedi.2024.100081 · doi ↗
- 4Ahmer, B.M.M., Tran, M. & Heffron, F., 1999, ‘The virulence plasmid of Salmonella Typhimurium is self-transmissible’, Journal of Bacteriology 181(4), 1364–1368. 10.1128/JB.181.4.1364-1368.19999973370 PMC 93521 · doi ↗ · pubmed ↗
- 5Ali, H.R., Hefny, E.G., Koraney, N.F., Ali, S.F., Abd Allah, M.I., Fadel, M.A. et al., 2025, ‘Antibiotic residues correlate with antibiotic resistance of Salmonella Typhimurium isolated from edible chicken meat’, Scientific Repots 15, 15165. 10.1038/s 41598-025-98189-4PMC 1204393540307359 · doi ↗ · pubmed ↗
- 6Almeida, F., Medeiros, M.I.C., Kich, J.D. & Falcão, J.P., 2016, ‘Virulence-associated genes, antimicrobial resistance and molecular typing of Salmonella Typhimurium strains isolated from swine from 2000 to 2012 in Brazil’, Journal of Applied Microbiology 120(6), 1677–1690. 10.1111/jam.1311026913828 · doi ↗ · pubmed ↗
- 7Andriyanov, P.A., Zhurilov, P.A., Liskova, E.A., Karpova, T.I., Sokolova, E. V., Yushina, Y.K. et al., 2021, ‘Antimicrobial resistance of Listeria monocytogenes strains isolated from humans, animals, and food products in Russia in 1950–1980, 2000–2005, and 2018–2021’, Antibiotics 10(10), 2000–2005. 10.3390/antibiotics 10101206 PMC 853277634680788 · doi ↗ · pubmed ↗
- 8Arizpe, A., Reveles, K.R., Patel, S.D. & Aitken, S.L., 2016, ‘Updates in the management of cephalosporin-resistant gram-negative bacteria’, Current Infectious Disease Reports 18, 39. 10.1007/s 11908-016-0552-727743202 PMC 11801173 · doi ↗ · pubmed ↗