Taxogenomic Analysis of a Novel Yeast Species, Lachancea rosae Sp. Nov. F.A., Isolated From the Wild Rose Rosa californica
Yakendra Bajgain, Quinn K. Langdon, Cara M. Krien, Martin Jarzyna, Kelly V. Buh, Max A. B. Haase, Anthony Pasles, John F. Wolters, Marizeth Groenewald, Chris Todd Hittinger, Dana A. Opulente

TL;DR
Scientists discovered a new yeast species, Lachancea rosae, from a wild rose in California and used genomic data to link its genes to how it processes carbon and nitrogen.
Contribution
A new yeast species is described with taxogenomic insights into carbon and nitrogen gene-trait correlations.
Findings
Strain yHQL494 belongs to a new species, Lachancea rosae, within the Lachancea genus.
Genomic and physiological analyses showed consistent gene-trait correlations across Lachancea species.
The new species exhibits distinct physiological traits like hyphae formation and carbon/nitrogen assimilation.
Abstract
A novel Saccharomycotina yeast strain, yHQL494, was isolated from the rose hip of the wild rose Rosa californica from Castle Crags State Park, California, USA. Phylogenetic analyses of both whole genome data and the sequences from the D1/D2 region of the large ribosomal subunit (LSU) rRNA gene placed strain yHQL494 within the genus Lachancea and grouped it into a clade with Lachancea lanzarotensis and Lachancea meyersii. Taxogenomic analyses were conducted on publicly available genome sequences to gain a deeper insight into the carbon and nitrogen gene‐trait associations across the Lachancea clade. The results of these analyses were found to be consistent across Lachancea species. Growth assays and microscopic analyses were conducted to determine the physiological characteristics of strain yHQL494, including the presence of hyphae or pseudohyphae, ascospore formation, fermentation…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
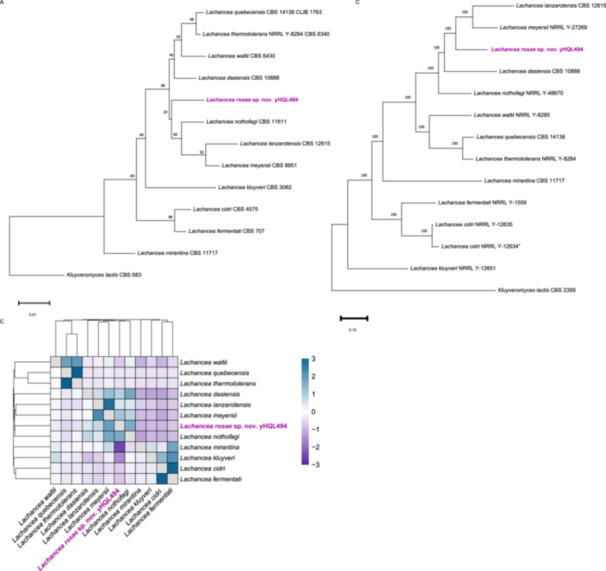 Figure 1
Figure 1 Figure 2
Figure 2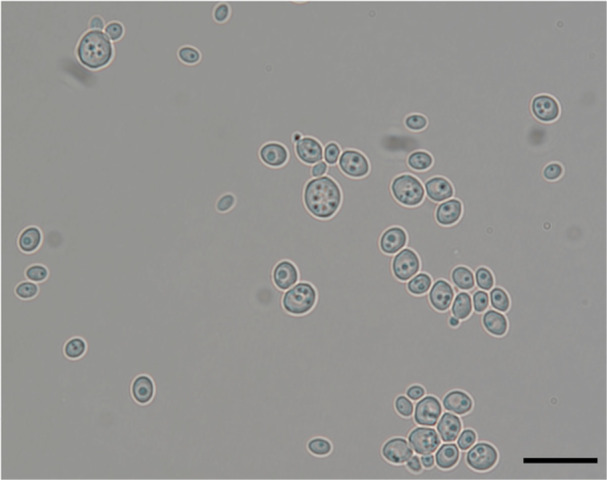 Figure 3
Figure 3| yHQL494 |
|
| ||
|---|---|---|---|---|
|
| Glucose | + | + | + |
| Mannose | + | NA | NA | |
| Fructose | d | NA | NA | |
| Inulin | + | v | — | |
| Sucrose | + | + | + | |
| Raffinose | + | + | + | |
| Melibiose | + | — | — | |
| Galactose | d | + | v | |
| Lactose | — | — | — | |
| Trehalose | + | + | + | |
| Maltose | + | + | + | |
| Melezitose | + | + | + | |
| Methyl‐alpha‐D‐glucoside | + | + | + | |
| Cellobiose | — | — | — | |
| Salicin | w/d | — | — | |
| L‐Sorbose | + | v | — | |
| Rhamnose | — | — | — | |
| Xylose | w | — | — | |
| L‐Arabinose | — | — | — | |
| D‐Arabinose | — | — | — | |
| Ribose | — | — | — | |
| Methanol | — | — | — | |
| Ethanol | + | + | + | |
| Glycerol | d | + | v | |
| Adonitol (syn. Ribitol) | d | v | — | |
| Erythritol | — | — | — | |
| Xylitol | — | — | v | |
| Galactitol | — | — | — | |
| Mannitol | d | + | + | |
| Sorbitol | + | + | + | |
| Myo‐inositol | — | — | — | |
| DL‐Lactate | — | — | — | |
| Succinate | — | — | — | |
| Citrate | — | — | — | |
| Gluconate | — | v | + | |
| Glucosamine | — | — | — | |
| N‐Acetyl‐D‐glucosamine | — | — | NA | |
| Hexadecane | — | — | NA | |
|
| Creatinine | — | — | NA |
| Nitrate | — | — | — | |
| Nitrite | — | — | — | |
| Lysine | + | + | — | |
| Allantoin | + | NA | NA | |
|
| Glucose | + | + | + |
| Galactose | — | — | v | |
| Sucrose | + | + | + | |
| Maltose | + | + | + | |
| Lactose | — | — | — | |
| Raffinose | — | + | NA | |
| Xylose | — | NA | NA | |
|
| 22°C | + | + | + |
| 30°C | + | + | + | |
| 37°C | — | — | — | |
| 42°C | — | — | — | |
| 45°C | — | — | — | |
| 50°C | — | — | — |
- —We thank the University of Wisconsin Biotechnology Center DNA Sequencing Facility for providing DNA sequencing facilities and services; Bill Vagt for providing the samples from which the strain was is
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFermentation and Sensory Analysis · Yeasts and Rust Fungi Studies · Horticultural and Viticultural Research
Introduction
1
The genus Lachancea belongs to the order Saccharomycetales and family Saccharomycetaceae (Groenewald et al. 2023). It was first proposed by Dr. Cletus P. Kurtzman in 2003 and was named in honor of Prof. Marc‐André Lachance. Currently, the genus consists of 11 described species (Kurtzman 2003; Porter et al. 2019; Opulente et al. 2024). The first five species described were previously classified in other genera, sometimes multiple genera, and were reassigned to Lachancea as Lachancea fermentati (previously Zygosaccharomyces fermentati), Lachancea cidri (previously Zygosaccharomyces cidri), Lachancea thermotolerans (previously Zygosaccharomyces thermotolerans), Lachancea kluyveri (previously Saccharomyces kluyveri), and Lachancea waltii (previously Kluyveromyces waltii), with L. thermotolerans as the type species for the genus (Kurtzman 2003; Porter et al. 2019; Opulente et al. 2024). Since the circumscription of Lachancea (Kurtzman 2003), six additional species have been formally described, Lachancea meyersii, Lachancea dasiensis, Lachancea nothofagi, Lachancea mirantina, Lachancea lanzarotensis, and Lachancea quebecensis (Fell et al. 2004; Lee et al. 2009; Mestre et al. 2010; Pereira et al. 2011; González et al. 2013; Freel et al. 2015).
Lachancea spp. have been isolated from multiple ecological niches, including plants, soil, food products, and beverages (Kurtzman et al. 2011; Opulente et al. 2018, 2024; Porter et al. 2019; Spurley et al. 2022). With a recent focus in exploring the enological importance of non‐Saccharomyces yeast spp., L. fermentati (Bellut et al. 2020), L. thermotolerans (Jolly et al. 2014), and L. lanzarotensis have been shown to be economically important due to their fermentative capabilities. In particular, L. thermotolerans has been shown to influence the aroma and flavor of wine through the production of esters, terpenes, and 3‐methylthio‐1‐propanol (Jolly et al. 2014; Benito 2018). Even though members of the genus Lachancea have not been implicated in any fungal diseases, there was a case where L. fermentati was attributed to causing fungemia in an immunocompromised individual (Leuck et al. 2014). However, such cases are rare, and Lachancea species are mostly associated with fermentation processes.
Using previously established yeast enrichment and isolation protocols (Sylvester et al. 2015; Opulente et al. 2019; Spurley et al. 2022), we identified a candidate for a novel Lachancea species, represented by strain yHQL494, that was isolated from the rose hips of the wild rose Rosa californica in Castle Crags State Park, California. Strain yHQL494 was suggested to represent a novel species after the internal transcribed spacer region (ITS) and the D1/D2 region of the large ribosomal subunit (LSU) rRNA gene were sequenced and compared to publicly available sequences (Kurtzman and Robnett 1998, 2013). In addition to traditional taxonomic methods, we performed whole genome sequencing and gene presence analyses to provide further insight into its genetic makeup, growth patterns, and phylogenetic relationship with known Lachancea species.
Materials and Methods
2
Species Isolation and Identification
2.1
Established enrichment protocols were used to isolate strain yHQL494 from a rose hip sample (Sylvester et al. 2015; Opulente et al. 2019; Spurley et al. 2022). The sample was collected by a community scientist, Bill Vagt, from the wild rose R. californica in Castle Crags State Park, California (41°9'9.825“N X 122°18'40.743“W), and the strain was isolated and identified by Quinn K. Langdon (Table S1). The sample was incubated at 30०C in liquid synthetic complete (SC) medium (yeast nitrogen base without amino acids, ammonium sulfate, or glucose 6.7 g/L; ammonium sulfate 5 g/L; drop‐out mix 1.7 g/L) with 8% glucose. Once there was visible growth, a secondary enrichment step was conducted in fresh SC medium with 8% glucose at 30०C before the cultures were plated on a yeast extract‐peptone‐dextrose‐agar (YPDA—yeast extract 10 g/L, peptone 20 g/L, glucose 20 g/L, agar 20 g/L) and also incubated at 30°C. Unique colony morphologies were sequenced using Sanger sequencing of the ITS and the D1/D2 region of the LSU rRNA gene for preliminary species identification.
Morphological and Physiological Characterization
3
Characteristics of the novel species were described through microscopic examination, as well as growth tests in liquid and solid media. Standard methods proposed by Kurtzman et al. (2011) were used to determine the physiological characteristics(Kurtzman et al. 2011). Briefly, to test the ability of the strain to assimilate specific carbon sources, a colony of strain yHQL494 was pre‐cultured in 0.1% glucose minimal medium (MM—yeast nitrogen base without amino acids, ammonium sulfate, or glucose 6.7 g/L; ammonium sulfate 5 g/L; 1 g/L glucose) and grown at 22°C ± 1°C for 1 day. The resulting culture was inoculated into liquid media containing different carbon sources, each at a concentration of 1%, and growth was visually inspected for 4 weeks at 22°C ± 1°C (Kurtzman et al. 2011). To test the ability of the strain to assimilate specific nitrogen sources, a colony of strain yHQL494 was pre‐cultured in YPD medium and grown at 22°C ± 1°C for 1 day. The resulting culture was inoculated into liquid minimal media (MM—yeast nitrogen base without amino acids, ammonium sulfate, or glucose 6.7 g/L; 10 g/L glucose) containing different nitrogen sources and grown for 1 week (Kurtzman et al. 2011). Cultures were then inoculated into fresh media for each nitrogen source, and growth was visually inspected for 4 weeks at 22°C ± 1°C. The Dalmau Plate method was used to determine the formation of true hyphae or pseudohyphae on YPDA plates. The colony morphology was determined using plate cultures grown on 2% Glucose, 0.5% yeast extract, 1% peptone, and 1.5% agar (GYPA) for 7 days at 25°C. Additionally, the strain was grown on GYPA, YPDA, malt‐extract agar (MEA), V8‐juice agar (V8), yeast extract‐malt extract agar (YMA), and yeast carbon base with ammonium sulfate (YCBAS) agar plates and incubated at 25°C for up to 2 months at 10°C and 25°C, and the plates were examined every week under a microscope to determine the formation of ascospores.
Taxogenomic Analyses
4
Genome Assembly
4.1
Whole genome sequence analyses were performed using the publicly available genome sequence JAJMGS000000000 and short read data SRR16988996.
Phylogenetic Analyses
4.2
The bioinformatic pipeline HybPiper (Johnson et al. 2016) was used to isolate and assemble the raw paired‐end reads of the D1/D2 region of the LSU rRNA gene and ITS region for strain yHQL494 using D1/D2 and ITS sequences from Saccharomyces cerevisiae. GenBank numbers for the ITS and the D1/D2 region of the LSU rRNA gene sequences for strain yHQL494 are ON040667 and ON064074, respectively. Sequences that have been extracted from genomes using bioinformatic tools such as HybPiper can be verified by using alignment tools such as BLAST to ensure accuracy. In the case of barcode sequences, you can align to closely related species. YeastIP, a database that contains the multiple barcode sequences of many species of yeasts in the subphylum Saccharomycotina (Weiss et al. 2013), along with GenBank and the Westerdijk Fungal Biodiversity Institute (CBS database), were used to obtain the publicly available Lachancea ITS and D1/D2 sequences (Table S2). These sequences were used to determine the relatedness of strain yHQL494 with known Lachancea species, phylogenies were made for the ITS region and D1/D2 region of the LSU rRNA gene of known Lachancea species and strains in MEGA (Kumar et al. 2018). The default parameters for CLUSTALW were used to align those sequences and a neighbor‐joining phylogeny was constructed using the default settings in MEGA with 1000 bootstrap replicates.
Pairwise ANI values were calculated from publicly available genomes using fastANI() with fragLen set to 500 bp (Opulente et al. 2024). The R package pheatmap (v. 1.0.12) was used to visualize the results of the ANI analysis. The ANI values were converted to a distanced by subtracting 100 from each value and then scaled using the scale parameter available in the function. Hierarchical clustering was performed using the average method.
Gene Trait Analyses
4.3
The methods and sequences described in Opulente et al. (2018) were used to detect gene presence for all publicly available genomes from Lachancea species and strain yHQL494. Most of the gene sequences are from S. cerevisiae (Opulente et al. 2018). However, the sequence for LAC4 was from Kluyveromyces lactis, XYL1 and XYL3 was from Schefferomyces stipitis, and YNR1, YNA1, and YNA2 were from Ogataea polymorpha (Opulente et al. 2018). The BLAST algorithm was used (tblastx), and an e‐value cutoff of 10^−10^ was used to infer gene presence (Altschul et al. 1990; Opulente et al. 2023). Genes that are known paralogs were combined and only reported once (Voordeckers et al. 2012; Opulente et al. 2018).
Results and Discussions
5
Phylogenetic Analyses
5.1
Initial BLAST alignments were used to assess sequence similarity between strain yHQL494 and the type strains of other closely related Lachancea species. For the 524‐nt D1/D2 region of the LSU rRNA gene, strain yHQL494 showed the highest sequence with L. dasiensis (98.04%; 9 substitutions, and 1 indel), followed by L. nothofagi (97.54%; 11 substitutions, 2 indels), L. meyersii (97.05%; 11 substitution, 5 indels), and L. lanzarotensis (96.60%; 15 substitutions, 3 indels) (Table S3). In the 604‐nt ITS region, strain yHQL494 shared 96.01% identity with L. meyersii (21 substitutions, 6 indels), 95.67% with L. nothofagi (24 substitutions, 2 indels), 94.67% with L. dasiensis (28 substitutions, 4 indels), and 94.52% identity with L. lanzarotensis (27 substitutions, 6 indels) (Table S3).
A phylogenetic analysis based on the D1/D2 region of the LSU rRNA gene placed strain yHQL494 in a clade with L. meyersii, L. lanzarotensis, and L. nothofagi (Figure 1A). Given the discrepancies between the BLAST results and the D1/D2 phylogeny, we extended our analysis to include additional Lachancea strains for all species in the genus for both the ITS region and D/D2 region (Figure S1A‐B, Table S4). In both our expanded ribosomal gene trees, strain yHQL494 was consistently in a clade with L. meyersii, L. lanzarotensis, and L. nothofagi.
(A) Neighbor‐Joining tree for the taxonomic type strains of the 11 described Lachancea species and Lachancea rosae sp. nov. (yHQL494) based on the D1/D2 region of the LSU rRNA gene. Bootstrap values (n = 1000) are indicated by numbers at the nodes. (B) Whole genome phylogeny pruned from (Opulente et al. 2024). Bootstrap values are indicated by numbers at the nodes. (C) Heatmap and hierarchical cluster analysis of pairwise average nucleotide index (ANI) values. Blue boxes indicate species that have higher ANI values, while purple boxes indicate lower ANI values. Hierarchical clustering was performed using the average method.
We compared our D1/D2 phylogeny (Figure 1A) with a pruned genome‐scale phylogenetic tree (Figure 1B) and found a strong topological similarity, with strain yHQL494 again in a clade with L. meyersii and L. lanzarotensis (Opulente et al. 2024). To (Figure 1C, Table S4). To further assess genomic relatedness, we calculated pairwise average nucleotide identity (ANI) values among Lachancea type strains. Hierarchical clustering of ANI values revealed species‐level structure, with strain yHQL494 sharing its highest ANI (75.72%) with L. meyersii (Figure 1C, Table S5).
Genomic and Phenotypic Analyses
5.2
The steady incorporation of whole genome sequences in the identification and taxonomic classification of novel yeast and bacterial species has delineated previously unknown genotype‐phenotype associations and helped construct robust phylogenies (Shen et al. 2018; Čadež et al. 2021; Opulente et al. 2024). We determined gene presence across Lachancea genomes to associate genome content with the utilization capacity of the yeasts on the different carbon and nitrogen sources that were tested (Figure 2). Our analyses contained multiple genes implicated in the metabolism of various carbon and nitrogen sources across yeasts (Table S6). In many cases, there was good correspondence in the genotype‐phenotype map (Figure 2). For example, all species except L. waltii possess GAL1, GAL7, and GAL10 and are capable of metabolizing galactose. The inability of L. waltii to grow on galactose is consistent with previous studies (Haase et al. 2021). Below we focused on the exception to the genotype‐phenotype map.
Metabolic traits and underlying genes for the carbon sources galactose, lactose, cellobiose, sucrose, raffinose, maltose, and xylose, as well as for the nitrogen sources nitrate and nitrite. Filled in squares indicates the ability to utilize a carbon or nitrogen source. Filled in circles correspond to gene presence. Gene presence was determined by BLAST analyses.
The absence of YNA1 and YNA2 from the genome of the majority of Lachancea species corroborated their inability to assimilate nitrite and nitrate compounds. Both YNA1 and YNA2 encode transcription factors involved in regulating nitrate and nitrite metabolism (Shen et al. 2018; Opulente et al. 2023). These genes were a more reliable indicator of this metabolism than homologs of the reductases encoded by YNR1, which was present in all genomes, a phenomenon previously noted (Opulente et al. 2023). Consistent with other literature (Nalabothu et al. 2023), the presence of xylose metabolism genes (XYL1, XYL2, and XYL3) did not always predict growth on xylose; 60% (9/15) of the Lachancea species possessed the genes necessary for growth on xylose but were unable to assimilate it (Figure 2, Table S6). Conversely, we observed that approximately 18% (3/15) of Lachancea species were able to utilize cellobiose; however, we were able to detect multiple BGL1 homologs, which encode β‐glucosidases associated with the utilization of cellobiose in some yeasts (Gurgu et al. 2011; Shen et al. 2018; Opulente et al. 2023), in all Lachancea genomes. This discrepancy suggests that not all BGL1 homologs are capable of supporting growth on cellobiose. Additionally, L. waltii lacks the ability to grow on maltose, despite containing homologs of all of the genes required for its metabolism (MAL11, MAL12, MAL13, IMA1‐4, and IMA5). This inability to grow in a certain medium despite containing the genes required for the metabolism of carbon sources in that medium has been attributed to the factors beyond gene presence or absence, including patterns of gene expression regulation and enzyme specificity (Riley et al. 2016).
Description of L. rosae F.A., Sp. Nov
6
Y. Bajgain, Q.K. Langdon, C.M. Krien, M. Jarzyna, K.V. Buh, M.A.B. Haase, A. Pasles, M. Groenew., Hittinger, D.A. Opulente. Mycobank no. MB857814.
Growth in 2% glucose‐yeast extract peptone broth and agar: After 3 days of aerobic growth at 24°C on YPDA the colony is butyrous, tannish‐white, smooth with an entire border, and dull (Figure 3A). The cells are spherical to ellipsoidal, 2−5 × 2.5−6 μm and occur singly or in pairs (Figure 3B). No sexual reproduction was observed.
Cell morphology of Lachancea rosae sp. nov. (yHQL494) grown on YPDA at 25°C. Bars = 5 μm.
Sexual reproduction: No sexual structures (asci or ascospores) are observed, so it is designated f.a. for forma asexualis (Lachance 2012).
Fermentation: While maltose is fermented well, glucose and sucrose are weakly fermented; galactose, lactose, raffinose, and xylose are not fermented (Table 1).
Assimilation of carbon compounds: Growth is observed in glucose, sorbitol, maltose, ethanol, and sucrose media. Delayed growth is observed in mannitol, glycerol, adonitol (syn. ribitol), fructose, and galactose media. Weak growth is observed in d‐xylose and citrate media, and no growth is observed in ribose, succinate, DL‐lactate, cellobiose, d‐arabinose, myo‐inositol, l‐rhamnose, lactose, xylitol, glucosamine, and gluconate (Table 1).
Assimilation of nitrogen compounds: Growth occurs in allantoin. Weak growth is observed in lysine, while creatine, nitrate, and nitrite show no growth (Table 1).
Temperature growth: Growth is observed at room temperature and 30°C. No growth is observed at 37°C.
Holotype: Strain yHQL494^T^ was isolated from the hip of the rose R. californica in Castle Crags State Park, California (GPS: 41.152729, −122.311318) and is cryopreserved in a metabolically inactive state in the yH strain collection at the University of Wisconsin‐Madison. Ex‐type strains are CBS 18,575^T^ and NRRL Y‐64858^T^.
Etymology: The epithet ro.sae L. fem. gen. n., rosae refers to Rosa, the genus of the host plant, R. californica, from which the type strain was isolated.
Genome sequence data: Whole genome analyses were performed using the publicly available genome JAJMGS000000000 and short read data SRR16988996 (Opulente et al. 2024).
Conclusions
7
The availability of whole genome sequencing data for multiple yeast species and advances in sequencing technologies and bioinformatic tools have greatly aided in the exploration of yeast biodiversity. In this study, we propose the name L. rosae for a new species, isolated from the hip bud of R. californica in Castle Crags State Park. Phylogenetic analyses place it within the genus Lachancea. While physiological and genomic data supported some genotype‐phenotype associations, others revealed mismatches, highlighting the complexity of metabolic pathways. Taxogenomic studies like this one contribute to resolving such discrepancies and laying the groundwork for future work, including machine learning approaches, to uncover alternative mechanisms underlying yeast traits. Overall, this study adds to the growing collection of diverse yeasts and demonstrates the power of integrated approaches in discovering and characterizing novel species.
Author Contributions
Dana A. Opulente, Quinn K. Langdon, and Chris Todd Hittinger conceived and designed the study. Yakendra Bajgain, Cara M. Krien, Anthony Pasles, Martin Jarzyna, Kelly V. Buh, Max A. B. Haase, Dana A. Opulente, and Quinn K. Langdon performed experiments. Yakendra Bajgain, Dana A. Opulente, Quinn K. Langdon, John F. Wolters, Martin Jarzyna, Kelly V. Buh, Max A. B. Haase, and Marizeth Groenewald analyzed data. Dana A. Opulente, Quinn K. Langdon, and Chris Todd Hittinger provided mentorship. Quinn K. Langdon and Chris Todd Hittinger secured funding. Yakendra Bajgain, Dana A. Opulente, and Chris Todd Hittinger wrote the paper with editorial input and approval from all authors.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Figure S1: A) Neighbor‐Joining tree for strains of the 11 described Lachancea species and Lachancea rosae sp. nov. (yHQL494) based on the D1/D2 region of the LSU rRNA gene (Table S2). Bootstrap values (n = 1000) are indicated by numbers at the nodes. B) Neighbor‐Joining tree for strains of the 11 described Lachancea species and Lachancea rosae sp. nov. (yHQL494) based on the ITS region (Table S2). Bootstrap values (n = 1000) are indicated by numbers at the nodes.
Table S1: Isolation information and ITS and D1/D2 sequences for yHQL494. Table S2: Strain information and accession numbers for ITS and D1/D2 sequences used in phylogenetic analyses (Figure 1A, Figure S1A – B). Table S3: Comparison of LSU and ITS sequences between yHQL494 and its closest relatives, Lachancea meyerii, Lachancea lanzarotensi, Lachancea nothofagi, and Lachancea dasiensis. Table S4: Newick tree of Lachancea species for the D1/D2 region of the LSU rRNA gene. Table S5: Pairwise average nucleotide index (ANI) values. Table S6: Results from TBLASTX for all genes in Figure 2.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Altschul, S. F. , W. Gish , W. Miller , E. W. Myers , and D. J. Lipman . 1990. “Basic Local Alignment Search Tool.” Journal of Molecular Biology 215: 403–410. 10.1016/S 0022-2836(05)80360-2.2231712 · doi ↗ · pubmed ↗
- 2Bellut, K. , K. Krogerus , and E. K. Arendt . 2020. “ Lachancea fermentati Strains Isolated From Kombucha: Fundamental Insights, and Practical Application in Low Alcohol Beer Brewing.” Frontiers in Microbiology 11: 764. 10.3389/fmicb.2020.00764.32390994 PMC 7191199 · doi ↗ · pubmed ↗
- 3Benito, S. 2018. “The Impacts of Lachancea thermotolerans Yeast Strains on Winemaking.” Applied Microbiology and Biotechnology 102: 6775–6790. 10.1007/s 00253-018-9117-z.29876604 · doi ↗ · pubmed ↗
- 4Čadež, N. , N. Bellora , R. Ulloa , et al. 2021. “Hanseniaspora Smithiae Sp. Nov., a Novel Apiculate Yeast Species From Patagonian Forests That Lacks the Typical Genomic Domestication Signatures for Fermentative Environments.” Frontiers in Microbiology 12: 679894.34367085 10.3389/fmicb.2021.679894 PMC 8334367 · doi ↗ · pubmed ↗
- 5Fell, J. W. , A. Statzell‐Tallman , and C. P. Kurtzman . 2004. “ Lachancea meyersii sp. nov., an Ascosporogenous Yeast From Mangrove Regions in the Bahama Islands.” Studies in Mycology 50: 359–363.
- 6Freel, K. C. , G. Charron , J. B. Leducq , C. R. Landry , and J. Schacherer . 2015. “ Lachancea quebecensis sp. nov., a Yeast Species Consistently Isolated From Tree Bark in the Canadian Province of Québec.” International Journal of Systematic and Evolutionary Microbiology 65: 3392–3399. 10.1099/ijsem.0.000426.26297665 · doi ↗ · pubmed ↗
- 7González, S. S. , J. Alcoba‐Flórez , and F. Laich . 2013. “ Lachancea lanzarotensis sp. nov., an Ascomycetous Yeast Isolated From Grapes and Wine Fermentation in Lanzarote, Canary Islands.” International Journal of Systematic and Evolutionary Microbiology 63: 358–363. 10.1099/ijs.0.046029-0.23125316 · doi ↗ · pubmed ↗
- 8Groenewald, M. , C. T. Hittinger , K. Bensch , et al. 2023. “A Genome‐Informed Higher Rank Classification of the Biotechnologically Important Fungal Subphylum Saccharomycotina .” Studies in Mycology 105: 1–22. 10.3114/sim.2023.105.01.38895705 PMC 11182611 · doi ↗ · pubmed ↗