Postnatal development of the dentate gyrus vascular niche
Nidhi Devasthali, India Carter, Angela I. Saulsbery, Elizabeth D. Kirby

TL;DR
This study shows that the blood vessel network in the dentate gyrus of mice continues to develop and refine after birth, influencing the positioning of neural stem cells.
Contribution
The study reveals how the vascular niche in the dentate gyrus matures postnatally, affecting neural stem cell organization.
Findings
Blood vessel density in the SGZ remains stable from 2 to 9 weeks of age in mice.
The average distance from NSPC somas to blood vessels decreases progressively during postnatal development.
Quiescent neural stem cells gradually shift towards the granule cell layer as development progresses.
Abstract
Lifelong neurogenesis in the dentate gyrus (DG) of the hippocampus supports cognitive and emotional functions in most adult mammals. The subgranular zone (SGZ) of the DG contains dense vasculature where neural stem and progenitor cells (NSPCs) reside in close proximity to local capillaries. This arrangement likely supports NSPCs by providing access to oxygen, circulating molecules, and endothelial-derived factors. While SGZ vessel density and NSPC association with vessels are well established in adulthood, when these niche attributes emerge in development remains unclear. Here, we show that while blood vessel density in the SGZ remained stable from initial layer formation (2 weeks of age) into young adulthood (9 weeks of age) in male and female mice, the average distance from NSPC somas to the nearest blood vessel decreased progressively over postnatal development. This finding was…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
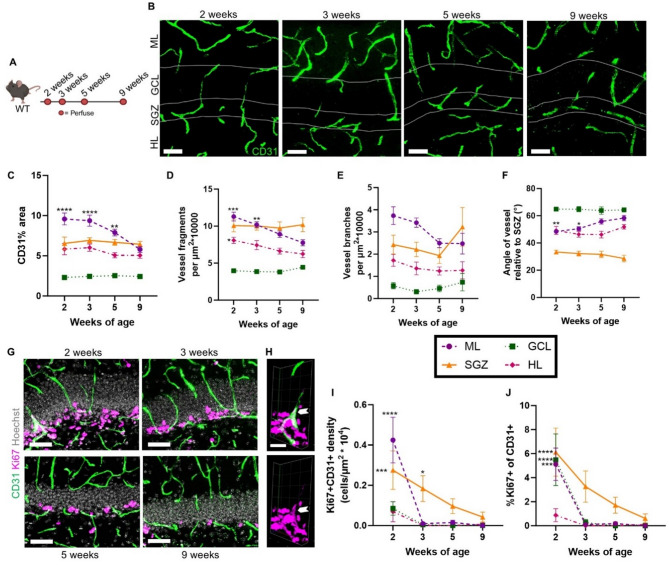 Figure 1
Figure 1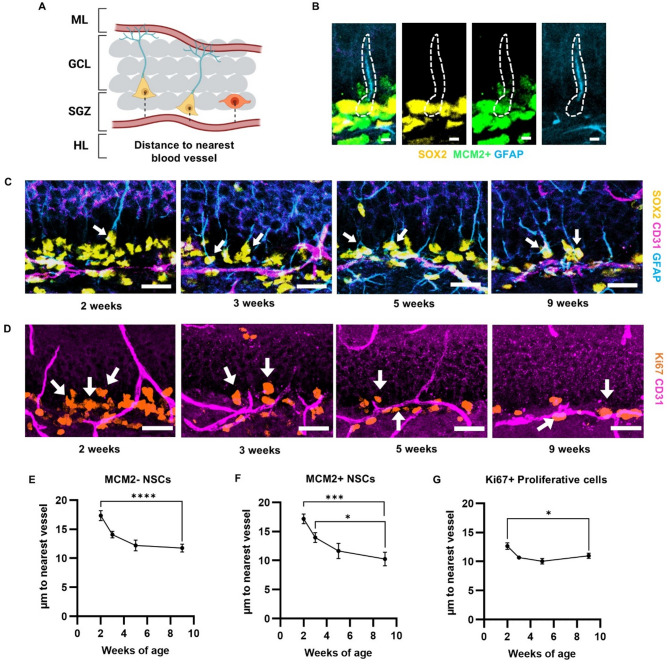 Figure 2
Figure 2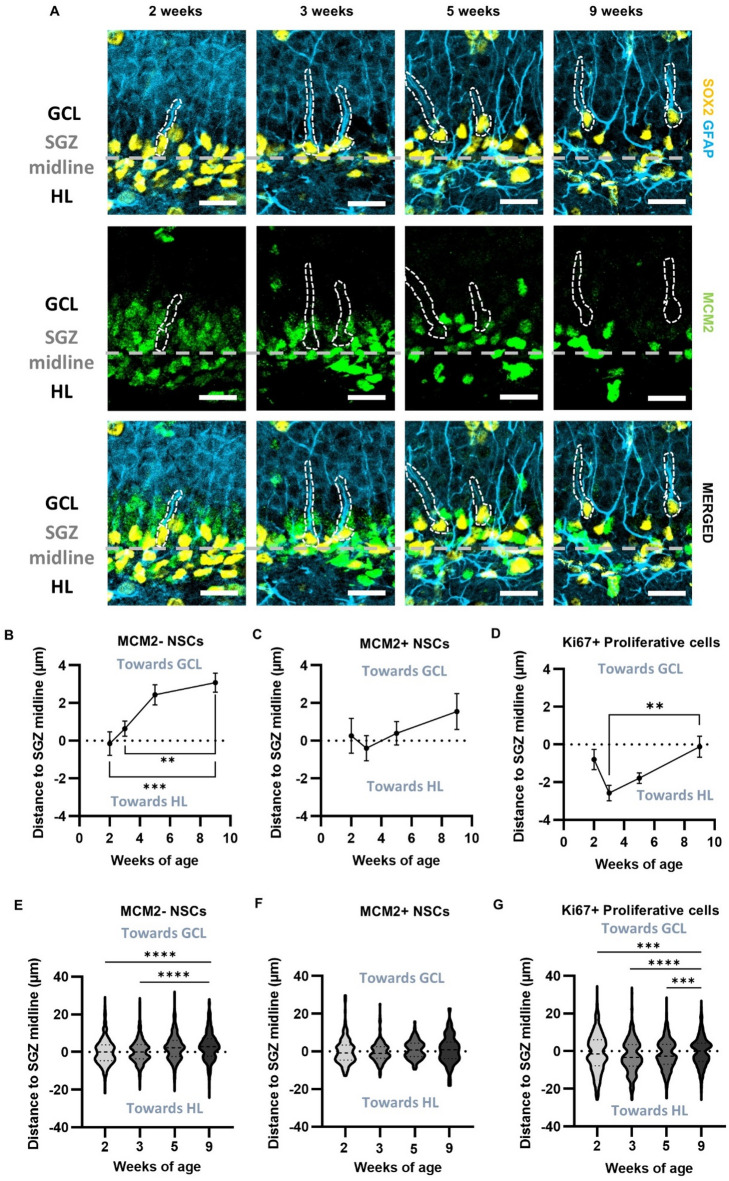 Figure 3
Figure 3- —National Science Foundation
- —https://doi.org/10.13039/100000065National Institute of Neurological Disorders and Stroke
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurogenesis and neuroplasticity mechanisms · Memory and Neural Mechanisms · Axon Guidance and Neuronal Signaling
Introduction
Adult neurogenesis occurs in two main neurogenic niches in the adult mammalian brain: the subventricular zone and the dentate gyrus (DG) of the hippocampus. In the mature DG, neurogenesis begins when radial glia-like neural stem cells (NSCs) in the subgranular zone (SGZ) divide to produce intermediate progenitor cells (IPCs), which subsequently mature into new neurons. This process of birth, survival, and maturation of new DG neurons is conserved across most mammalian species^1–3^. Studies in rodents have demonstrated that newborn DG neurons integrate in and modulate existing circuits, thereby supporting spatial learning and memory and affective behaviors^4–8^. Dysregulation of adult neurogenesis has been linked to neurological diseases such as Alzheimer’s disease and may even be a potential therapeutic target in other diseases and disorders with impaired hippocampal memory^7,9–11^.
In the adult DG, a major, essential feature of the stem cell niche is the local blood vessel network^12–15^. DG blood vessels originate as capillaries at the hippocampal fissure, expanding into a branched network in the molecular layer (ML). ML capillaries then converge and feed into capillaries that descend perpendicularly through the granule cell layer (GCL)^16^. These descending capillaries then turn 90° to run parallel to the SGZ, where they form a dense vascular network^16^. Within this dense, planar vasculature of the SGZ, NSCs (which are largely quiescent) and proliferating cells reside in close proximity to the blood vessels^13,17^. NSCs also wrap around blood vessels in the inner ML of the DG via their apical process^18,19^. This close association of neural stem and progenitor cells (NSPCs) to vessels may provide these neurogenic cells with heightened access to circulating oxygen, signaling molecules, and/or nutrient metabolites^16,20^. It may also uniquely expose neurogenic cells to endothelial-derived soluble factors which can inhibit differentiation, enhance neurogenesis, and promote self-renewal of NSCs^21,22^. Blood vessels may also serve as scaffolds guiding migration of IPCs and their differentiating progeny^17^.
Although the key features of vessel density and NSPC association with vessels are well characterized in the adult brain, the developmental progression of these features remains unclear. In rodents, the structure of the DG begins to develop prenatally from NSCs in the hippocampal primordium but is not complete until the second postnatal week when the layers of the DG are fully formed^23^. During the first postnatal week in mice, most of the granule neurons that will comprise the adult DG are generated from NSCs^23^. From early postnatal age, around postnatal day (PD) 17, to adulthood, around PD 60, there is a significant reduction in neurogenesis in the mouse DG^24^. This reduction in neurogenesis is accompanied by a decline in the number of proliferating cells during the same PD 17 to PD 60 time period^24^.
While the postnatal development of the neurogenic cell lineage has become better characterized in recent years, relatively little is known about how the local vasculature develops as these cell populations emerge and mature. Angiogenesis, the creation of new blood vessels from previously established ones, is generally considered complete in the brain by 3 weeks of age^25^. But these estimates are not specific to the unique DG neurogenic niche. When the vascular features of the DG layers arise and when the NSPC association with that vasculature is formed remain unclear. The DG vascular features and NSPC proximity to vasculature could develop in step with the formation of the DG and exhibit the adult phenotype as early as 2 weeks of age (when DG layers are first established), or conversely, they could continue to mature through postnatal development and into adulthood. To address this gap, we examined the vascular density across DG layers and the physical distance between NSPCs and local blood vessels from the juvenile age when DG layers are first formed (2 weeks) through young adulthood (9 weeks).
Results
Vascular architecture in the DG is dynamic over the course of postnatal development
To characterize changes in vascular architecture of each of the DG layers over postnatal development, we used immunolabeling for CD31 (a vascular endothelial marker) in brains harvested from male and female C57Bl/6J mice at 2, 3, 5 and 9 weeks of age (Fig. 1A, B). These ages span from the juvenile period when the DG layers are first formed (2 weeks) to young adulthood (9 weeks). To quantify vascular architecture, we measured vascular coverage, vessel orientation, number of blood vessel fragments, and number of vessel branch points. To determine vascular coverage, we measured percent area covered by CD31 immunoreactivity in the ML, GCL, SGZ, and hilus (HL) of the DG (Fig. 1C, data separated by sex in Fig S1). We found that vascular coverage did not change significantly between 2 and 9 weeks of age in the SGZ (all data reported as mean ± SEM; 2 weeks: 6.55 ± 0.78%; 3 weeks: 6.91 ± 0.33%; 5 weeks: 6.66 ± 0.37%; 9 weeks: 6.44 ± 0.36%), GCL (2 weeks: 2.30 ± 0.20%; 3 weeks: 2.45 ± 0.25%; 5 weeks: 2.53 ± 0.15%; 9 weeks: 2.44 ± 0.24%), and HL (2 weeks: 5.83 ± 0.70%; 3 weeks: 6.02 ± 0.45%; 5 weeks: 5.07 ± 0.33%; 9 weeks: 5.05 ± 0.36%). In the ML, vascular density decreased with postnatal age. CD31+ immunolabeled area at 9 weeks of age (5.80 ± 0.38%) was significantly lower than at 2 weeks (9.58 ± 0.74%), 3 weeks (9.37 ± 0.70%), and 5 weeks (7.89 ± 0.41%) of age.
To determine the number of vessel fragments, we counted the number of CD31+ vessel structures per unit area (fragment density) in the ML, GCL, SGZ, and HL of the DG (Fig. 1D, data separated by sex in Fig S1). The fragment density showed similar patterns as overall vascular coverage, with SGZ, HL and GCL showing stable fragment density across ages while the ML fragment density decreased with age. Specifically, we found that the density of vessel fragments did not change significantly between 2 and 9 weeks of age in the SGZ (2 weeks: 10.06 ± 0.83 fragments/µm^2^ * 10^4^; 3 weeks: 10.02 ± 0.56 fragments/µm^2^ * 10^4^; 5 weeks: 9.75 ± 0.82 fragments/µm^2^ * 10^4^; 9 weeks: 10.20 ± 0.94 fragments/µm^2^ * 10^4^) and GCL (2 weeks: 3.97 ± 0.21 fragments/µm^2^ * 10^4^; 3 weeks: 3.85 ± 0.21 fragments/µm^2^ * 10^4^; 5 weeks: 3.80 ± 0.23 fragments/µm^2^ * 10^4^; 9 weeks: 4.44 ± 0.24 fragments/µm^2^ * 10^4^). Blood vessel fragment density decreased with postnatal age in the HL and ML. In the HL, the density of blood vessel fragments at 9 weeks of age (5.14 ± 0.49 fragments/µm^2^ * 10^4^) was significantly lower than at 2 weeks (8.08 ± 0.41 fragments/µm^2^ * 10^4^). HL fragment density at 3 weeks (7.43 ± 0.62 fragments/µm^2^ * 10^4^) and 5 weeks (6.64 ± 0.40 fragments/µm^2^ * 10^4^) did not differ significantly from 9 weeks. In the ML, blood vessel fragment density at 9 weeks of age (7.75 ± 0.41 fragments/µm^2^ * 10^4^) was significantly lower than at 2 weeks (11.29 ± 0.59 fragments/µm^2^ * 10^4^) and 3 weeks (10.18 ± 0.40 fragments/µm^2^ * 10^4^), but did not differ from 5 weeks (8.91 ± 0.44 fragments/µm^2^ * 10^4^).
To quantify vessel branching, we quantified the density of points from which multiple CD31 + vessel structures appeared to originate in the ML, GCL, SGZ, and HL of the DG (Fig. 1E, data.
Fig. 1. During postnatal development vascular coverage of the DG is stable in most layers and angiogenesis declines to near undetectable levels. (A) Experimental timeline to investigate the postnatal development of the DG neurogenic niche. (B) Representative images of immunolabeling for CD31. Scale bar = 50 μm. (C) Proportion of DG area covered by CD31 immunolabeling. 2-way repeated measures ANOVA age x layer (F_9,120_) = 9.06, p < 0.0001; age (F_3,40_) = 2.20, p = 0.10; layer (F_3,120_) = 315.8, p < 0.0001. **- p = 0.0038, ****- p < 0.0001 from Dunnett’s multiple comparisons to 9 weeks within layer. (D) Density of CD31 + vessel fragments. 2-way repeated measures ANOVA age x layer (F_9,120_) = 3.044, p = 0.0026; age (F_3,40_) = 2.284, p = 0.0936; layer (F_2.274,90.94_) = 146.9, p < 0.0001. *- p = 0.0265; **- p = 0.0010; ***- p = 0.0004 from Dunnett’s multiple comparisons to 9 weeks within layer. (E) Density of CD31 + vessel branch points. 2-way repeated measures ANOVA age x layer (F_9,120_) = 2.292, p = 0.0207; age (F_3,40_) = 0.8007, p = 0.5009; layer (F_1.972,78.87_) = 60.14, p < 0.0001. (F) Angle of CD31 + vessel structures relative to SGZ orientation. 2-way repeated measures ANOVA age x layer (F_9,120_) = 2.236, p = 0.0240; age (F_3,40_) = 1.348, p = 0.2725; layer (F_2.705,108.2_) = 179.2, p < 0.0001. *- p = 0.0122, ***- p = 0.0011 from Dunnett’s multiple comparisons to 9 weeks within layer. (G) Representative images of immunolabeling with CD31 and Ki67. Scale bar = 50 μm. (H) 3D representation of CD31 + Ki67 + immunolabeling from area outlined in (G) with white arrowhead indicating CD31 + Ki67 + double labeled cell. Scale bar = 20 μm. (I) Density of CD31 + Ki67 + double labeled cells. 2-way repeated measures ANOVA age x layer (F_9,120_) = 5.09, p < 0.0001; age (F_3,40_) = 11.28, p < 0.0001; layer (F_3,120_) = 12.41, p < 0.0001. *- p = 0.04; ***- p = 0.0005; ****- p < 0.0001 from Dunnett’s multiple comparisons to 9 weeks within layer. (J) Proportion of CD31 + cells co-labeled with Ki67. 2-way repeated measures ANOVA age x layer (F_9,120_) = 2.27, p = 0.02; age (F_3,40_) = 12.60, p < 0.0001; layer (F_3,120_) = 8.39, p < 0.0001. ***- p = 0.0001, ****-p < 0.0001 from Dunnett’s multiple comparisons to 9 weeks within layer. (n = mice) 2 (n = 10), 3 (n = 12), 5 (n = 11) and 9 (n = 11) weeks.
separated by sex in Fig S1). In general, the GCL showed the lowest branching point density with the ML and SGZ showing higher branching. However, there were no significant age-related changes in branching density between 2 and 9 weeks of age in any layer: the SGZ (2 weeks: 2.43 ± 0.42 branch points/µm^2^ * 10^4^; 3 weeks: 2.21 ± 0.32 branch points/µm^2^ * 10^4^; 5 weeks: 1.94 ± 0.35 branch points/µm^2^ * 10^4^; 9 weeks: 3.23 ± 0.87 branch points/µm^2^ * 10^4^), GCL (2 weeks: 0.58 ± 0.14 branch points/µm^2^ * 10^4^; 3 weeks: 0.31 ± 0.10 branch points/µm^2^ * 10^4^; 5 weeks: 0.46 ± 0.15 branch points/µm^2^ * 10^4^; 9 weeks: 0.74 ± 0.39 branch points/µm^2^ * 10^4^), HL (2 weeks: 1.73 ± 0.27 branch points/µm^2^ * 10^4^; 3 weeks: 1.36 ± 0.29 branch points/µm^2^ * 10^4^; 5 weeks: 1.25 ± 0.18 branch points/µm^2^ * 10^4^; 9 weeks: 1.27 ± 0.39 branch points/µm^2^ * 10^4^) or ML (2 weeks: 3.74 ± 0.40 branch points/µm^2^ * 10^4^; 3 weeks: 3.42 ± 0.21 branch points/µm^2^ * 10^4^; 5 weeks: 2.50 ± 0.20 branch points/µm^2^ * 10^4^; 9 weeks: 2.48 ± 0.47 branch points/µm^2^ * 10^4^).
To determine vessel orientation, we measured the angle of CD31+ vessel structures relative to the orientation of the SGZ in the ML, GCL, SGZ, and HL of the DG (Fig. 1F, data separated by sex in Fig S1). As expected, GCL vessels showed orientations closest to 90°, reflecting their widely reported perpendicular orientation relative to the SGZ layer. SGZ vessels, in contrast, were closest to 0°, reflecting their well-established planar orientation parallel to the SGZ layer. ML and HL vessels showed intermediate average orientation. Vessel orientation was largely stable across ages in SGZ, GCL and HL, but changed in the ML. Specifically, ML vessels were more perpendicular to the SGZ with increasing postnatal age. The mean vessel angle at 9 weeks of age (58.28 ± 1.94°) was significantly greater than at 2 weeks (48.41 ± 0.93°) and 3 weeks (50.52 ± 1.43°), but not at 5 weeks (55.83 ± 1.20°) of age. Vessel orientation did not change significantly between 2 and 9 weeks of age in the SGZ (2 weeks: 33.35 ± 1.38°; 3 weeks: 32.20 ± 1.79°; 5 weeks: 31.60 ± 2.73°; 9 weeks: 28.62 ± 2.43°), GCL (2 weeks: 64.89 ± 1.15°; 3 weeks: 64.83 ± 1.86°; 5 weeks: 63.80 ± 2.69°; 9 weeks: 64.36 ± 0.74°), and HL (2 weeks: 49.25 ± 2.95°; 3 weeks: 46.35 ± 2.11°; 5 weeks: 46.15 ± 2.48°; 9 weeks: 51.87 ± 1.63°). Put together, these findings suggest that the vascular architecture of most of the DG remains stable over postnatal development, with the exception of the ML and HL showing moderate decreases in vessel fragments between juvenile and young adult ages, and the ML also showing a decrease in vascular density and a more perpendicular vessel orientation.
Angiogenesis in the DG is rare in postnatal development
We next combined CD31 immunolabeling with labeling for Ki67 (a cell cycle protein) to assess angiogenesis. We were able to identify only a small number of Ki67+ CD31+ cells in all layers of the DG at 2 weeks of age (Fig. 1G, H, I, data separated by sex in Fig. S2) (ML: 0.43 ± 0.11 cells/µm^2^ * 10^4^; GCL: 0.09 ± 0.03 cells/µm^2^ * 10^4^; SGZ: 0.28 ± 0.10 cells/µm^2^ * 10^4^; HL: 0.07 ± 0.05 cells/µm^2^ * 10^4^). By 3 weeks of age, Ki67+ CD31+ cells were barely detectable in all DG layers (ML: 0.01 ± 0.01 cells/µm^2^ * 10^4^; GCL: 0.00 ± 0.00 cells/µm^2^ * 10^4^; SGZ: 0.18 ± 0.06 cells/µm^2^ * 10^4^; HL: 0.00 ± 0.00 cells/µm^2^ * 10^4^). Ki67+ CD31+ cells were largely absent in all DG layers by 5 weeks of age (ML: 0.02 ± 0.01 cells/µm^2^ * 10^4^; GCL: 0.00 ± 0.00 cells/µm^2^ * 10^4^; SGZ: 0.10 ± 0.04 cells/µm^2^ * 10^4^; HL: 0.00 ± 0.00 cells/µm^2^ * 10^4^), a finding which was maintained at 9 weeks of age (ML: 0.00 ± 0.00 cells/µm^2^ * 10^4^; GCL: 0.00 ± 0.00 cells/µm^2^ * 10^4^; SGZ: 0.04 ± 0.03 cells/µm^2^ * 10^4^; HL: 0.01 ± 0.01 cells/µm^2^ * 10^4^). The rarity of proliferative CD31+ endothelia could also be appreciated when assessed as a proportion of CD31+ cells that were Ki67+ (Fig. 1J, data separated by sex in Fig. S2). At 2 weeks of age, less than 7% of CD31+ cells were Ki67+ (ML: 5.12 ± 1.35%; GCL: 5.49 ± 2.15%; SGZ: 6.13 ± 2.00%; HL: 0.88 ± 0.54%). By 3 weeks of age, less than 4% of CD31+ cells were Ki67+ (ML: 0.10 ± 0.06%; GCL: 0.26 ± 0.26%; SGZ: 3.26 ± 1.30%; HL: 0.00 ± 0.00%), then less than 2% by 5 weeks of age (ML: 0.18 ± 0.13%; GCL: 0.00 ± 0.00%; SGZ: 1.71 ± 0.66%; HL: 0.00 ± 0.00%). By 9 weeks, the percent of CD31+ co-labeled for Ki67 was less than 1% (ML: 0.00 ± 0.00%; GCL: 0.00 ± 0.00%; SGZ: 0.61 ± 0.38%; HL: 0.10 ± 0.10%). Overall, these findings suggest that angiogenesis is rare in juvenile mice and declines to near or below detection limits in adulthood.
Proliferative and quiescent NSPC soma proximity to vasculature increases over postnatal development
To examine proximity of NSPC somas to blood vessels over postnatal development, we quantified the shortest distance from proliferative and quiescent cell bodies in the SGZ to CD31+ blood vessel structures at 2, 3, 5, and 9 weeks of age (Fig. 2A). We first quantified NSPC distance to vessel in projected 2D images, similar to the original study establishing the close association of proliferative cells with blood vessels^13^. We identified NSCs as SOX2+ cell bodies in the SGZ with a GFAP+ apical process extending through the GCL. Quiescent NSCs (qNSCs) were distinguished from proliferating NSCs by co-labeling for MCM2, a nuclear cell cycle protein (Fig. 2B). The general proliferative population, which is primarily IPCs, was identified as Ki67+ nuclei in the SGZ. Quiescent and proliferative subsets of NSPCs all showed a similar pattern of being found closer to vessels as postnatal age advanced from juvenile ages (2 weeks) into young adulthood (9 weeks) (Fig. 2C-G, data separated by sex in Fig. S3). Both MCM2- and MCM2+ NSCs were significantly closer to blood vessels at 9 weeks of age as compared to earlier time points (MCM2- NSCs: 2 weeks 17.35 ± 0.8526 μm; 3 weeks 14.08 ± 0.5590 μm; 5 weeks 12.20 ± 0.9253 μm; 9 weeks 11.78 ± 0.6851 μm; MCM2+ NSCs: 2 weeks 17.18 ± 0.8464 μm; 3 weeks 13.95 ± 0.8353 μm; 5 weeks 11.64 ± 1.327 μm; 9 weeks 10.27 ± 1.169 μm) (Fig. 2E, F). Similarly, Ki67+ proliferative cells were significantly closer to vessels at 9 weeks of age than at 2 weeks of age (Ki67+ proliferative cells: 2 weeks 12.66 ± 0.5507 μm; 3 weeks 10.69 ± 0.2427 μm; 5 weeks 10.03 ± 0.4764 μm; 9 weeks 10.98 ± 0.4422 μm) (Fig. 2G). At younger ages (2 and 3 weeks of age), we observed a small but detectable presence of Ki67+ cells outside the SGZ, in the inner GCL. We therefore repeated the analysis of Ki67+ cell proximity to vessel including cells in both the SGZ and GCL. When all Ki67+ cells in the SGZ and GCL were considered across age, the data were largely indistinguishable from the data from SGZ alone (Fig. S4).
To confirm that these findings were replicable when distance was measured in 3 dimensions that included tissue depth, we generated 3D reconstructions from z-stacks of immunolabeled sections from 2 to 9 weeks of age and measured distance from NSCs and Ki67+ proliferative cells to the nearest vessel. Similar to the 2D analysis, 3D analysis revealed that both NSCs and proliferative cells were closer to vessels at 9 weeks than at 2 weeks (NSCs: 2 weeks 18.98 ± 0.9842 μm; 9 weeks 14.09 ± 0.8970 μm; Ki67+ cells: 2 weeks 13.84 ± 0.4126 μm; 9 weeks 12.46 ± 0.5433 μm) (Fig S5). Together, these findings suggest that NSC and IPC association with blood vessels continues to mature well into postnatal development.
Quiescent NSCs become located closer to the GCL over postnatal development
To further investigate the NSPC niche throughout postnatal development, we quantified the position of quiescent and proliferative NSPCs within the DG relative to the midline of the SGZ at 2, 3, 5 and 9 weeks of age by measuring the shortest distance from the center of NSPC somas to the midline of the SGZ.
Distances toward the GCL were assigned positive values and distances toward the HL were assigned negative values (Fig. 3A). We found that the mean location of GFAP+ SOX2+ MCM2- (quiescent) NSC somas shifted significantly toward the GCL over postnatal development (Fig. 3A, B, data separated by sex in Fig S6). The MCM2- NSCs were significantly closer to the GCL at 9 weeks of age as compared to both 2 and 3 weeks of age (MCM2- NSCs: 2 weeks − 0.1503 ± 0.6201 μm; 3 weeks 0.6368 ± 0.4042 μm; 5 weeks 2.436 ± 0.5384 μm; 9 weeks 3.084 ± 0.5020 μm) (Fig. 3B).
Fig. 2NSCs and proliferative cells in the DG get closer to blood vessels with age. (A) Graphic representation of the DG and the distance measurement taken from the center of the soma of NSCs and proliferative cells to the nearest blood vessel. NSCs are shown with yellow cell bodies and blue apical processes. Proliferative cells are shown as orange cell bodies. (B) Representative images of identification of an activated NSC using SOX2, MCM2, and GFAP immunolabeling. White outline is an identified NSC. Scale bar = 5 μm. (C) Representative images of CD31, SOX2, and GFAP immunolabeling. White arrows are pointing to identified NSCs. Scale bar = 20 μm. (D) Representative images of CD31 and Ki67 immunolabeling. White arrows are pointing to identified proliferative cells. Scale bar = 20 μm. (E) MCM2- NSC average distance to nearest CD31+ vessel. One-way ANOVA p < 0.0001, F_(3,38)_ = 10.83, ****- p < 0.0001 from Dunnett’s multiple comparisons to 9 weeks of age. (F) MCM2+ NSC average distance to nearest CD31+ vessel. One-way ANOVA p = 0.0003, F_(3,38)_ = 7.936. ***- p = 0.0002, *- p = 0.0413 from Dunnett’s multiple comparisons to 9 weeks of age. (G) Ki67+ proliferative cell average distance to nearest CD31+ vessel. One-way ANOVA p = 0.0011, F_(3,38)_ = 6.556. * - p = 0.0264 from Dunnett’s multiple comparisons to 9 weeks of age. Mean ± SEM shown throughout with n = individual mice. 2 weeks (n = 10), 3 weeks (n = 12), 5 weeks (n = 10) and 9 weeks (n = 10).
The general pattern seen in the mean soma location of the MCM2- NSCs was not found in either proliferative cell population, where mean soma location was largely stable over age (MCM2+ NSCs: 2 weeks 0.2577 ± 0.9202 μm; 3 weeks − 0.3914 ± 0.6713 μm; 5 weeks 0.3959 ± 0.6227 μm; 9 weeks 1.548 ± 0.9527 μm, Ki67+ proliferative cells: 2 weeks − 0.7986 ± 0.5421 μm; 3 weeks − 2.276 ± 0.4840 μm; 5 weeks − 1.784 ± 0.2807 μm; 9 weeks − 0.1159 ± 0.5567 μm) (Fig. 3C, D).
To look more closely at each cell type’s spatial distribution in the DG, we used violin plots to visualize cell soma location distribution relative to the SGZ midline. As expected based on the mean cell soma location data, the majority of cells in the MCM2- NSC population were found gradually closer to the GCL border of the SGZ with age (Fig. 3E). The MCM2+ NSC population, in contrast, was spread more evenly across the SGZ and this distribution was relatively stable across ages (Fig. 3F). The Ki67+ proliferative population was also centered in the SGZ across ages, though it became more concentrated around the SGZ midline throughout postnatal development as the population of Ki67+ qualitatively dwindled (Fig. 3G). The relative locations of the NSCs and Ki67+ cells were similar when expressed as a location relative to the total width of the GCL (Fig S7). All together, these results suggest that the physical location of NSPCs in the local DG niche continue to mature throughout development.
Discussion
The hallmark features of the DG neurogenic vascular niche are widely hypothesized to be critical for supporting lifelong neurogenesis. Here, we show that while some aspects of the vascular structure of the DG are present when the DG layers are first formed at 2 weeks of age in mice, the vascular niche continues to mature throughout postnatal development into young adulthood. Overall, vascular properties including vessel density, fragment density, branching and orientation remained stable in the SGZ and GCL. ML vasculature showed the most changes, with progressive decreases in vessel coverage and fragment density accompanied by a net shift in orientation to be more perpendicular to the GCL/SGZ layers with increasing postnatal age. Within the SGZ, NSPCs were particularly dynamic, showing progressive coalescence of multiple cell classes around vessels with advancing age, along with shifts in average anatomical location of cell somas within the SGZ niche. Together, our findings reveal a DG vascular niche that continues to refine postnatally.
During embryonic development, the majority of brain vascularization is achieved via angiogenesis, the division of existing vascular endothelial cells to create new blood vessels^26^. Angiogenesis is generally considered complete by 3 weeks of age in the mouse brain, though this data is mostly based on measurement of cortical tissue^27^. Consistent with these previous findings, we found almost no evidence of angiogenesis in the DG beyond 2 weeks of age, as reflected by the scarcity of CD31+ endothelial cells expressing the cell cycle marker Ki67. We further found that the vascular density in the GCL, SGZ, and HL layers of the DG niche remained constant from juvenile ages to young adulthood. The maintenance of vascular coverage along with the absence of angiogenesis suggests that vascular endothelial cells in the GCL, SGZ and HL likely elongate or widen the diameter of blood vessels to ensure consistent vascular coverage as total brain volume increases during postnatal development. The ML layer of the DG, in contrast, lost vascular density between 2 and 9 weeks of age. The total volume of the ML increases by similar amounts to the GCL over postnatal development^28^. Our finding therefore raises the question of why the ML vasculature fails to maintain growth to keep pace with overall brain volume changes like other layers of the DG do.
Fig. 3. Quiescent NSCs in the DG are located closer to the GCL with age. (A) Representative images of SOX2, GFAP, and MCM2 immunolabeling. The SGZ midline is denoted by a dashed grey line with the GCL above and HL below. White outlines are identifying quiescent MCM2- NSCs. Scale bar = 20 μm. (B) MCM2- NSC average distance to SGZ midline. One-way ANOVA p = 0.0002, F_(3,38)_ = 8.447. ***- p = 0.0003, **- p = 0.0040 from Dunnett’s multiple comparisons to 9 weeks of age. (C) MCM2+ NSCs average distance to SGZ midline. One-way ANOVA p = 0.3820, F_(3,38)_ = 1.049. (D) Ki67+ proliferative cells average distance to SGZ midline. One-way ANOVA p = 0.0121, F_(3,38)_ = 4.158. **- p = 0.0069 from Dunnett’s multiple comparisons to 9 weeks of age. (B – D) Mean ± SEM shown throughout with n = individual mice. 2 weeks (n = 10), 3 weeks (n = 12), 5 weeks (n = 10) and 9 weeks (n = 10). (E) Violin plots of distance to SGZ midline from individual MCM2- NSCs. p < 0.0001 between 9 and 2 weeks and p < 0.0001 between 9 and 3 weeks of age from Kolmogorov-Smirnov test. 2 weeks (n = 632 cells), 3 weeks (n = 838 cells), 5 weeks (n = 763 cells), and 9 weeks (n = 760 cells). (F) Violin plots of distance to SGZ midline from individual MCM2+ NSCs. 2 weeks (n = 181 cells), 3 weeks (n = 192 cells), 5 weeks (n = 85 cells), 9 weeks (n = 55 cells). (G) Violin plots of distance to SGZ midline from individual Ki67+ proliferative cells. p = 0.0002 between 9 and 2 weeks, p < 0.0001 between 9 and 3 weeks, and p = 0.0001 between 9 and 5 weeks of age from Kolmogorov-Smirnov test. 2 weeks (n = 856 cells), 3 weeks (n = 1043 cells), 5 weeks (n = 892 cells), 9 weeks (n = 847 cells).
In the adult mammalian DG, the close proximity of NSPC bodies to blood vessels in the SGZ has 2 major hypothesized functions: (1) to bring NSC progeny in proximity to vessels that they will use as scaffolding for migrating along as they differentiate^17^, and (2) to provide specialized access to blood and/or endothelial signals that support their survival and proliferation^16,20,22,29^. Our data show the average distance between blood vessels and NSPC somas decreased over juvenile development into young adulthood, suggesting an increasing association of NSPCs to vessels as the DG matured. This increasing association of NSPCs to vessels during development was observed in both quiescent and proliferating NSPCs, suggesting that the developmental emergence of NSPC-vessel association did not depend on the proliferation state of NSPCs.
The progressive increase in average association of proliferative NSPCs with local vessels suggests functional importance of this relationship. In the proliferative cell population, this increase in vascular association with age also aligns well with the cellular compression around the SGZ midline that occurred in parallel. The SGZ is more highly vascularized than the neighboring layers, particularly the GCL, so a cellular location closer to the middle of the SGZ will bring cells on average closer to more blood vessels.
Quiescent NSCs also showed increasing proximity to SGZ vessels with age but in this case, this greater vascular association was coupled with a shift further away from the center of the highly vascularized SGZ, closer to the vascularly sparse GCL. This combination of concentrating in a generally less vascular area while increasing average vessel association strongly suggests that quiescent NSC proximity to blood vessels is important for NSC persistence. It is also worth noting that, in adulthood, the SGZ-GCL border is tightly packed with the cell bodies of dentate granule cells, with little space in between for NSCs^19^. NSCs are therefore being found in a generally more avascular space that is also more densely packed than regions closer to the hilus. Combined, these findings suggest some strong driving force for either selectively maintaining or drawing in quiescent NSCs to this region that cannot be explained by general vascular density or available extracellular space. What signals drive NSCs away from the SGZ midline and toward the GCL remain a topic for future study.
Our data do not establish the cellular or molecular mechanisms by which NSPC-vessel proximity develops postnatally. One possible cellular mechanism explaining how NSPC somas become closer to vessels on average over time is through the selective maintenance or proliferation of NSPCs closest to blood vessels. Over the course of postnatal development, the number of NSPCs decreases dramatically^28,30^. This decrease is particularly notable in the proliferative cell population as NSCs become progressively more quiescent and IPCs and their progeny differentiate^31,32^. If proximity to blood vessels promotes maintenance and/or proliferation of NSPCs, then the loss of NSPCs farther away from blood vessels would decrease the average NSPC-vessel distance over time. An additional possible mechanism for increased NSPC soma association with vessels over postnatal development is that NSPCs may migrate towards blood vessels as the niche matures. Finally, a third possibility is that vessels migrate towards NSPCs. All three of these cellular mechanisms could be operating simultaneously or in any combination to drive greater NSPC-vessel association. Ultimately, further investigation into the mechanisms driving the development of NSPC-vessel proximity is needed to determine how the association develops.
The molecular signals that direct increased postnatal NSPC-vessel association are also unclear. Broadly speaking, blood-derived signals seem to be likely candidates which could promote NSPC survival and proliferation near vessels, and/or migration towards vessels. For example, growth factors like insulin-like growth factor and brain-derived neurotrophic factor are found in the blood and promote NSPC proliferation and survival embryonically and during adulthood^33–37^. Blood-borne signals have been shown to drive adult neurogenesis changes in response to systemic stimuli like exercise^36,38–41^ and aging^22,29,42,43^, and this type of blood-borne regulation may extend to this postnatal refinement of the neurogenic vascular nice. NSPC-derived signals also could play a role in postnatal maturation of NSPC-vessel association. For example, NSPCs secrete a variety of factors that could affect their own migration or the physiology of local blood vessels^44–48^. Most notably, NSPCs synthesize vascular endothelial growth factor (VEGF)^48,49^. VEGF is required for normal vascularization of the embryonic brain, with endothelial cells growing towards the VEGF secreted by embryonic NSCs^50,51^. In adulthood, we recently showed that DG NSPC-derived VEGF is essential for maintaining NSPC-vessel association^52,53^. Surprisingly, this dependence appeared to rely on self-stimulated NSPC motility and cell attachment, rather than changes in vascularization. In the current study, NSPC-VEGF could play a role in vessels migrating towards NSPCs and/or maintaining existing NSPC-vessel proximity. More work is needed to define the molecular signals that guide postnatal vascular niche maturation.
There are several limitations to our study. First, while we restricted our investigation of NSPC-vessel association to the SGZ, previous literature has demonstrated that NSC processes wrap around blood vessels in the ML, and potentially gain access to blood factors at these contact points^18,19^. In this present study, we visualized the main radial process of NSCs using GFAP immunolabeling. Though GFAP immunolabel allows for identification of NSCs based on the existence of that radial process, it does not fill the farther out fine processes that make vessel contact, meaning that vessel contact could not be quantified here. Future work could measure vessel wrapping using transgenic reporter mice to visualize NSC processes coupled with immunolabeling to visualize blood vessels. Electron microscopy would also be advisable to achieve the resolution required to fully characterize wrapping^19^. Exploring NSC-vessel wrapping at functional and structural levels during postnatal development, a time when the vascular architecture of the ML in particular is changing, could lend greater insight into the hypothesized functional importance of vessel wrapping on NSC survival and proliferation. Second, our analysis of vascular coverage by DG layer reflects broad changes in vascular patterns, but would be unable to reveal any subtle movements of vessels within DG layers. Studies using live imaging of blood vessels could identify vasculature migration patterns, and determine if migration of blood vessels contributes to increasing NSPC-vessel association. Third, our 2D and 3D analyses of NSPC proximity to blood vessels only account for NSPC proximity to a single vessel. The proximity of NSPCs to all vessels in its vicinity is not a measure we quantify here, and has not been reported for the DG vascular niche to our knowledge. Such a quantification would be of interest when considering that a cell being moderately close to multiple vessels may be equally beneficial as a cell being extremely close to one vessel. Fourth, we did not investigate the development of the blood-brain barrier (BBB) in the DG during postnatal development. It is known that the BBB in the adult DG neurogenic niche is not leaky^16^, which stands in contrast to the well-known leakiness of the BBB in the other major neurogenic niche, the subventricular zone^54,55^. The BBB in rodents mostly matures during the first 3 postnatal weeks (including the rat hippocampus) during which time the hippocampal BBB is already less leaky compared to cortical regions^56,57^. When the BBB specifically in the DG completes development is not known. More work is needed to investigate when the BBB develops in the DG, and how the varying vascular density of the different DG layers may affect the maturation of the BBB.
In conclusion, we have found that the DG neurogenic niche continues to mature through postnatal development. Our characterization of the DG niche during juvenile development forms a framework for future investigation into the mechanisms guiding the changes we observed. Identifying the mechanisms of niche development and determining their functional relevance would ultimately help better understand how the hallmark features of the adult DG neurogenic niche arise. Understanding the development of key niche features could also aid in creating microenvironments receptive to stem cell transplants, paving the way for treatments for several neurological disorders and diseases.
Methods
Authors complied with ARRIVE guidelines while conducting and reporting experiments described here.
Mice
Timed pregnant wild-type C57BL/6J female mice (n = 7) were obtained from Jackson labs (#000664) and were housed individually with a toy in The Ohio State University Psychology Building mouse vivarium. All mice were housed in standard ventilated cages on a 12/12 hour light/dark cycle with ad libitum access to food and water. The pups were weaned at 3 weeks of age and housed in same sex groups of 2–5. All animal use was in accordance with institutional guidelines approved by The Ohio State University Institutional Animal Care and Use Committee.
Experimental design
Mice were randomly selected from 7 litters at 2 weeks (n = 10, 5 M/5F), 3 weeks (n = 12, 8 M/4F), 5 weeks (n = 11, 5 M/6F), and 9 weeks (n = 11, 5 M/6F) of age. Each timepoint had mice from at least 6 of the 7 litters. No mice with viable immunolabeling were excluded from final analyses. Immunolabeling was insufficiently clear or absent in one or more channels for 2 mice in the GFAP/SOX2/MCM2/CD31 immunolabeling only (1 mouse from 5 weeks of age group, 1 mouse from 9 weeks of age group). All mice received intraperitoneal injections of ethynyl-deoxyuridine (Click Chemistry Tools, #1149 − 500), dissolved fresh in sterile physiological saline at 10 mg/ml at dosage 150 mg/kg 2 h before being perfused. Mice were euthanized at the indicated tissue harvest time by anesthesia with a mixture of 87.5 mg/kg ketamine and 12.5 mg/kg xylazine, then transcardial perfusion with 0.1 M phosphate buffered saline (PBS).
Tissue processing
Mouse brains were post-fixed with 4% paraformaldehyde in 0.1 M PBS at 4 °C for 24 h. They were then equilibrated in 30% sucrose in 0.1 M PBS at 4 °C. Brains were then sliced on a freezing microtome in a 1:12 series of coronal sections at 40 μm thick. Sections were stored in cryoprotectant medium at -20 °C until used for immunofluorescent staining.
Immunolabeling
For Ki67 and CD31 immunolabeling, free-floating sections were rinsed in PBS, blocked in 0.3% triton-100X and 1% normal donkey serum in PBS for 30 min at room temperature, then incubated in primary antibody in blocking solution overnight on a shaker at 4 °C (Table 1). The next day, the sections were rinsed with PBS then incubated in secondary antibodies (Table 1) in blocking solution for 2 h on a shaker at room temperature. The sections were then labeled with Hoechst 33,342 (1:2000 in PBS, Fisher #H3570), followed by final rinses in PBS.
For GFAP/SOX2/CD31/MCM2 labeling, slices were first labeled for SOX2 as above, followed by a 10 min fixation step in 4% paraformaldehyde at room temperature. Then they were labeled for GFAP/CD31/MCM2 following the protocol above.
Immunolabeled sections were mounted on SuperFrost Plus slides, dried overnight at room temperature, and cover slipped with Prolong Gold Antifade mounting medium. For long term storage, slides were placed in the dark at 4 °C.
Table 1. Antibodies used for immunolabeling.PrimaryantibodyVendor/product noDilutionSecondaryantibodyVendor/product noDilutionRat anti-CD31BDPharmingen 5502741:100DonkeyAlexafluor594anti-ratFisherA212091:500Rabbitanti-Ki67Cell SignalingTechnology 91291:400DonkeyAlexafluor647anti-rabbitFisherA315731:500Ratanti-SOX2AffymetrixeBioscience 14-98111:1000Fab fragmentGoat AlexaFluor594JacksonImmuno112- 587-0081:500Mouseanti-GFAPEMD MilliporeMAB3601:1000DonkeyAlexa fluor350antimouseFisherA100351:1000Rabbitanti-MCM2Cell SignalingTechnology 40071:500DonkeyAlexa fluor647antirabbitFisherA-315731:500
Microscopy
The DG was imaged at 20x magnification in 15 μm z-stacks with 1 μm steps using Zeiss Axio Observer Z.1 with apotome digital imaging system and axiocam 506 monochrome camera (Zeiss blue version 2.3, http://www.zeiss.com).
Image quantification
For all image quantification, 3 DG images from 2 to 3 sections were used per mouse. Layers of the DG for all image quantification were defined as – ML: the region between the suprapyramidal blade of the granule cell layer and the stratum lacunosum-moleculare of the CA1 region combined with the region between the infrapyramidal blade of the granule cell layer and the end of the hippocampal structure; GCL: the dense layer made up of cell bodies of granule cells excluding the SGZ; SGZ: the zone spanning 2 cell body widths between the granular cell layer and the hilus; HL: the region between the suprapyramidal and infrapyramidal blades of the granule cell layer, excluding the SGZ and CA3 regions.
For vascular coverage analysis, layers of the DG (ML, GCL, SGZ, and HL) were outlined in maximum intensity projection images of the z-stacks. CD31 immunolabeling signal was processed using the despeckle tool in ImageJ (version 1.54d, http://imagej.org) to reduce noise, then thresholded by intensity. The %Area covered by thresholded signal in each layer was measured.
For vessel orientation analysis, the SGZ was outlined in maximum intensity projection images of the z-stacks. Using the Angle (disconnected) tool in ZEN software (version 2.3 (blue edition), http://www.zeiss.com) the direction of 5 CD31 immunolabeled vessel structures per layer per image was compared to the direction of the midline axis of the SGZ. All resulting values were expressed as acute angles (0°-90°).
For vessel fragments and branching analyses, fragments and branches of CD31 immunolabeled vessel structures were identified in the DG layers. Branches were defined as points where 2 or more vessel structures appeared to contact each other. Fragments were defined as the uninterrupted portion of CD31 immunolabeled vessel structures between 2 branch points, between a branch point and the apparent “beginning/end” of a CD31 immunolabeled vessel structure, or the portion of a CD31 immunolabeled vessel structure between apparent “beginning/end” points. To express counts as density measures, fragment and branch counts were divided by the area sampled as measured by ZEN.
For proliferating vascular endothelial cell counts, Hoechst+ nuclei in CD31 immunolabeled vessel structures were identified in the DG layers then categorized as Ki67+ or Ki67-. To express counts as density measures, Ki67+ CD31+ counts were divided by the area sampled as measured by ZEN. For percent Ki67+ CD31+ cells, the number of double positive cells was divided by the total number of CD31+ cells in that layer to yield a percent measure.
For analysis of NSC distance to vessels, 90 cells/mouse that had a SOX2+ nucleus in the SGZ and a GFAP+ apical process extending through the GCL were identified and the shortest distance from the center of their cell body to the nearest CD31+ vessel structure was measured using the ZEN line measure tool. The identity of the NSCs was confirmed and whether their nucleus was MCM2+ or MCM2- was determined in the full Z-stack images.
For analysis of proliferative cell distance to vessels, 90 cells/mouse that were Ki67+ in the SGZ were identified and their distance to the nearest CD31+ vessel structure (regardless of vessel orientation) was measured using the ZEN line measure tool.
For analysis of NSPC distance to vessels in 3D, z-stacks were visualized in Imaris (version 10.2.0, imaris.oxinst.com) 3D imaging software. 30 NSCs and 30 proliferative cells were identified per mouse. NSCs were identified as a SOX2+ nucleus in the SGZ and a GFAP+ apical process extending through the GCL. Proliferative cells were identified as Ki67+ nuclei in the SGZ. The shortest distance from the center of their cell body to the nearest CD31+ vessel structure (regardless of vessel orientation) was measured using the Measurement Points tool.
For the analysis of both NSC and Ki67+ location data, the SGZ midline was identified as the border between the GCL and HL and outlined using the ZEN contour tool. The shortest distance to SGZ midline from the center of the cell body of the 90 cells/mouse previously identified was measured using the ZEN line measure tool. Positive distance measurement values were assigned to cells that were closer to the GCL from the SGZ midline. Negative distance measurement values were assigned to cells that were closer to the HL from the SGZ midline. For analysis of Ki67+ location relative to the SGZ midline, GCL-HL border was identified using Hoechst labeling as described above. For the analysis of NSC location relative to the SGZ midline, the SGZ midline was identified as falling directly below the cell dense negative GCL layer in the CD31 labeling.
For the analysis of both NSC and Ki67+ location data relative to the total width of the GCL, the total width of the GCL in the location of 30 NSCs and 30 Ki67+ cells per mouse was measured using the ZEN line measure tool. The GCL total width was defined as the length between the ML border of the GCL and the lower HL border of the GCL (also referred to as the SGZ midline). The distance from each cell to the SGZ midline (also the 0.0 μm point of the GCL total width) was then divided by the total width of the GCL in that cells location and multiplied by 100 to get a percentage distance into the GCL. Positive distance measurement values were assigned to cells that were on the side of the SGZ midline closer GCL. Negative distance measurement values were assigned to cells that were on the side of the SGZ midline closer to the hilus.
Statistical analysis
All statistical tests were conducted using GraphPad Prism (version 10.0.3 (275), www.graphpad.com), except for 3-way repeated measures ANOVA that was conducted using IBM SPSS Statistics (version 29.0.1.0 (171), www.ibm.com). Data on CD31+ vascular coverage, and density and percent of CD31+ cells that were Ki67+, were analyzed by 2-way repeated measures ANOVA (age x layer) followed by Dunnett’s multiple comparisons test within layer compared to the control age of 9 weeks. Data on distance of MCM2+ NSCs, MCM2- NSCs and Ki67+ proliferative cells to nearest CD31+ vessel were analyzed by ordinary one-way ANOVAs for each cell type followed by Dunnett’s multiple comparisons test for each time point compared to the control age of 9 weeks. Data on distance of NSCs and Ki67+ proliferative cells to nearest CD31+ vessel at 2 and 9 weeks of age in 3D were analyzed by Mann-Whitney U tests for each cell type. Data on location of NSCs and Ki67+ proliferating cells relative to SGZ midline were analyzed by ordinary one-way ANOVAs for each cell type followed by Dunnett’s multiple comparisons test for each time point compared to the control age of 9 weeks. The overall cell populations of MCM2+ NSCs, MCM2- NSCs and Ki67+ proliferating cells location relative to the SGZ midline were also analyzed by a Kolmogorov-Smirnov test for each time point compared to the control age of 9 weeks. 3-way repeated measures ANOVAs (layer x age x sex) were performed for CD31+ vascular coverage data, and density and percent of CD31+ cells that were Ki67+. 2-way ANOVAs (age x sex) were performed for MCM2+ NSC, MCM2- NSC, and Ki67+ proliferating cells data to determine whether sex effects were present. All data were approximately normally distributed, all statistical tests were two-tailed with α = 0.05. No strong patterns of sex differences were observed in any measure so main data analysis shows data collapsed by sex.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Charvet, C. J. & Finlay, B. L. Comparing adult hippocampal neurogenesis across species: translating time to predict the tempo in humans. Front. Neurosci.12, 1452 (2018).10.3389/fnins.2018.00706 PMC 618207830344473 · doi ↗ · pubmed ↗
- 2He, N., Zhang, L., Cui, J. & Li, Z. Bone marrow vscular niche: home for hematopoietic stemcells. Bone Marrow Res.2014, 128436 (2014).10.1155/2014/128436 PMC 400911324822129 · doi ↗ · pubmed ↗
- 3Sun, G. J. et al. Tangential migration of neuronal precursors of glutamatergic neurons in the adult mammalian brain. Proc. Natl. Acad. Sci.112, 9484–9489 (2015).10.1073/pnas.1508545112 PMC 452276326170290 · doi ↗ · pubmed ↗
- 4Nicola, Z., Fabel, K. & Kempermann, G. Development of the adult neurogenic niche in the hippocampus of mice. Front. Neuroanat.9, 7456 (2015).10.3389/fnana.2015.00053 PMC 442345025999820 · doi ↗ · pubmed ↗
- 5Nieto-Estévez, V., Defterali, Ç. & Vicario-Abejón, C. IGF-I: A key growth factor that regulates neurogenesis and synaptogenesis from embryonic to adult stages of the brain. Front. Neurosci.10, 4526 (2016).10.3389/fnins.2016.00052 PMC 476306026941597 · doi ↗ · pubmed ↗
- 6Karakatsani, A., Shah, B. & Ruiz de Almodovar, C. Blood vessels as regulators of neural stem cell properties. Front. Mol. Neurosci.12, 1452 (2019).10.3389/fnmol.2019.00085 PMC 647303631031591 · doi ↗ · pubmed ↗
- 7Kirby, E. D., Kuwahara, A. A., Messer, R. L. & Wyss-Coray, T. Adult hippocampal neural stem and progenitor cells regulate the neurogenic niche by secreting VEGF. Proc. Natl. Acad. Sci.112, 4128–4133 (2015).10.1073/pnas.1422448112 PMC 438639725775598 · doi ↗ · pubmed ↗
- 8Miller, L. N. et al. Neural stem and progenitor cells support and protect adult hippocampal function via vascular endothelial growth factor secretion. Mol. Psychiatry 2024, 1–16 10.1038/s 41380-024-02827-8 (2024).10.1038/s 41380-024-02827-8PMC 1201438039528687 · doi ↗ · pubmed ↗