Effects of dietary oyster peptide supplement on litter performance, immunological response of sows and growth performance of piglets
Xiaofeng Tian, Jialu Wang, Congcong Yao, Muyang Shao, Yatian Qi, Zihao Gao, Hongguo Wang, Wei Xia, Zhigang Wang, Chenyu Tao, Junjie Li

TL;DR
This study shows that adding oyster peptides to sows' diets improves piglet health, survival, and immune response by enhancing placental nutrient transport and reducing inflammation.
Contribution
The novel finding is that oyster peptides improve sow and piglet outcomes through specific molecular mechanisms in placental transport and inflammation.
Findings
Oyster peptide supplementation increased healthy piglets and weaning survival rates.
Oyster peptides enhanced immunoglobulin levels in colostrum and placental transporter gene expression.
Oyster peptides reduced pro-inflammatory markers and increased anti-inflammatory and antioxidant gene levels in the placenta.
Abstract
Animal breeding and reproduction techniques has led to an increase in the number of offspring of sows. However, weak piglets and low survival rate of piglets occurs frequently. During pregnancy, the late gestation is the most crucial period for fetal weight gain. Oyster peptides (OPI) are regarded as functional active substances derived from oysters with significant nutritional and medicinal value, exhibiting considerable potential for application. It remains unclear whether oyster peptides can play an important role in the field of sow reproduction. In this study, the objective was to investigate the impact of incorporating OPI into the diets of sows during the late gestation and lactation phases on litter performance, immunological response of sows and the growth and development of piglets. 100 sows were selected and were randomly divided into the control group (CON, basic diet) and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
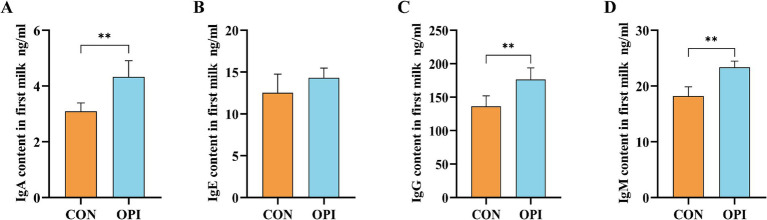 Figure 1
Figure 1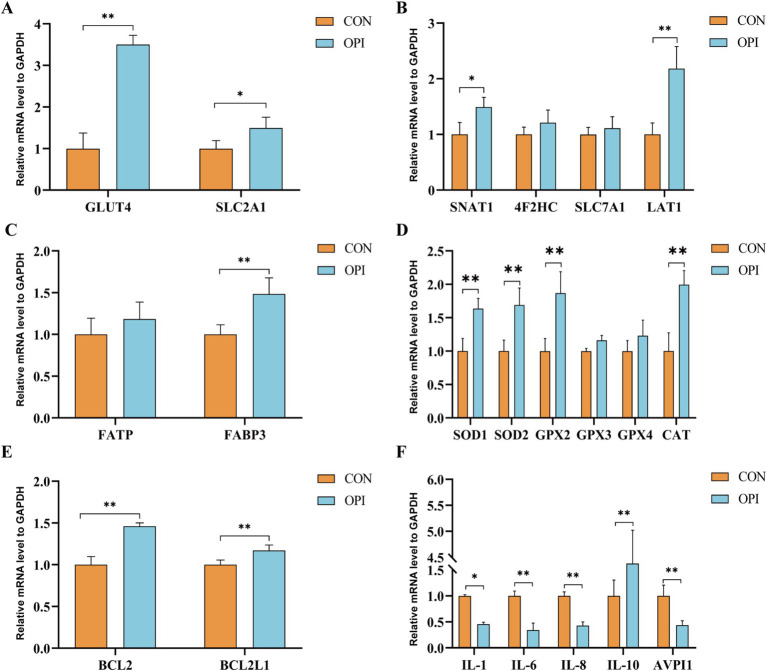 Figure 2
Figure 2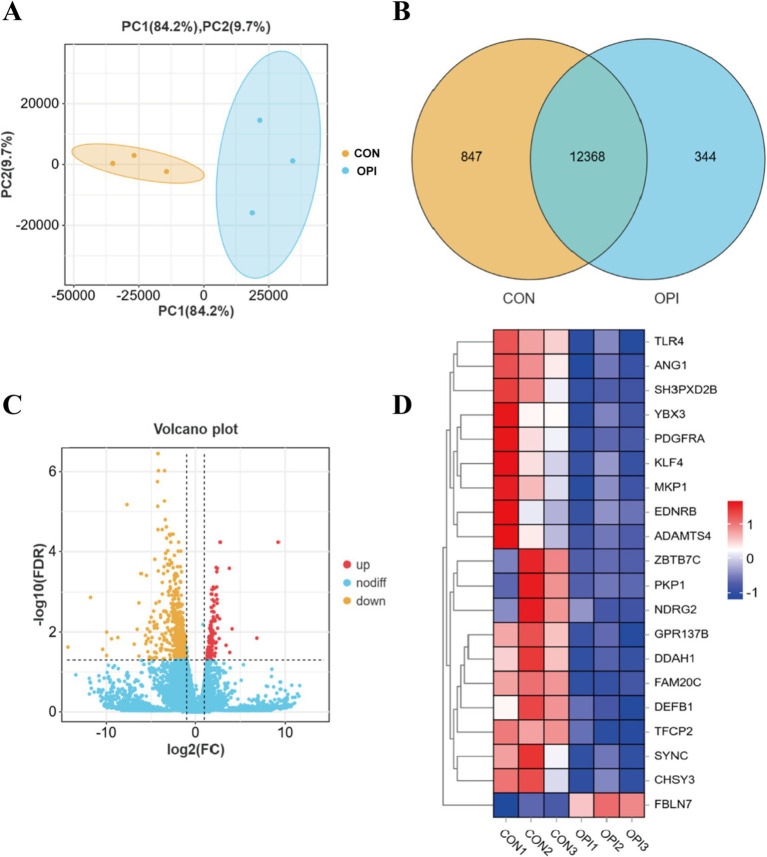 Figure 3
Figure 3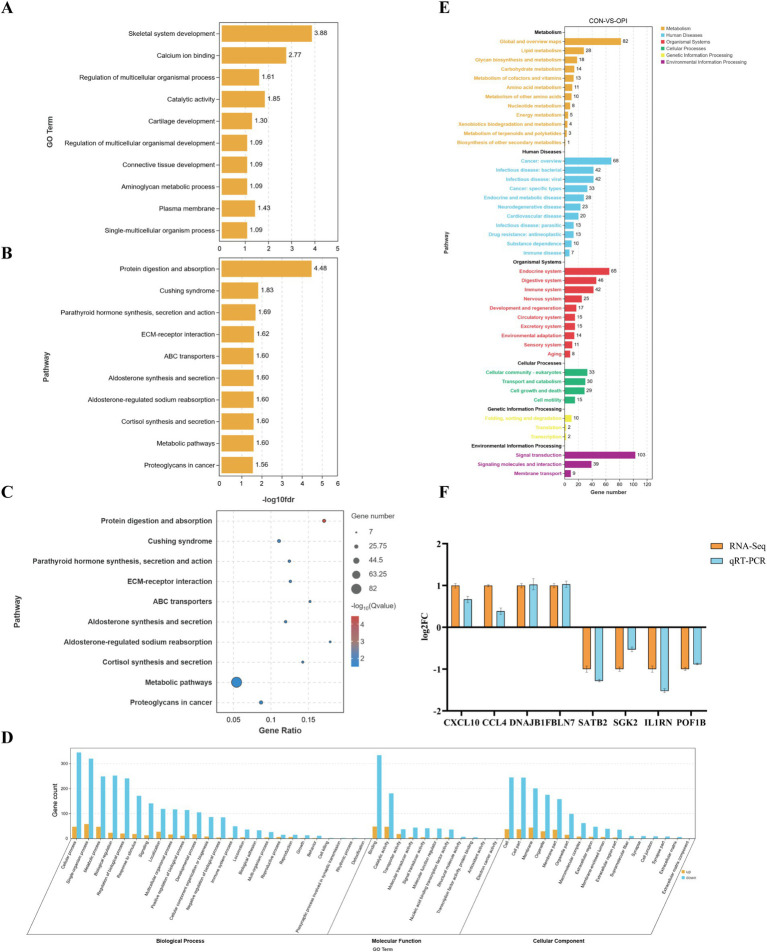 Figure 4
Figure 4| Basal diet for gestating sows | Value (%) | Basal diet for lactating sows | Value (%) |
|---|---|---|---|
| Ingredient composition (%) | |||
| Corn | 53 | Corn | 61.6 |
| Wheat bran | 12.5 | Low-protein soybean meal (43%) | 20.5 |
| Low-protein soybean meal (43%) | 10.8 | Soybean hulls | 4.3 |
| Secondary flour (ash 3.5%) | 10 | 4% lactating sow premix | 4 |
| Rice bran meal | 5 | Full-fat rice bran | 3 |
| 4% gestating sow premix | 4 | Rice bran meal | 2.4 |
| Soybean hulls | 3 | Soybean oil | 2 |
| Bentonite | 1 | Imported fishmeal (Peru) | 1 |
| Soybean oil | 0.5 | Bentonite | 1 |
| Imported mycotoxin adsorbent | 0.2 | Imported mycotoxin adsorbent | 0.2 |
| Nutrient composition | |||
| Crude protein (%) | 14.16 | Crude protein (%) | 16 |
| Crude fat (%) | 3.45 | Crude fat (%) | 5.12 |
| Crude fiber (%) | 5.57 | Crude fiber (%) | 4.5 |
| Digestible energy (kcal/kg) | 2947.87 | Digestible energy (kcal/kg) | 3278.00 |
| Net energy (kcal/kg) | 2122.19 | Net energy (kcal/kg) | 2332.01 |
| Lysine (%) | 0.80 | Lysine (%) | 1.04 |
| Calcium (%) | 0.78 | Calcium (%) | 0.96 |
| Phosphorus (%) | 0.35 | Phosphorus (%) | 0.41 |
| Primer name | Primer sequence (5′–3′) | Accession number |
|---|---|---|
|
| F: TGAAGGTCGGAGTGAACGGA | |
| R: TGGGTGGAATCATACTGGAACA | ||
|
| F: CCTGGGCCGATTGGTTATTG | |
| R: CAGAATCCCGATGACGATGC | ||
| SLC2A1 | F: GCTCCTGGTCCTGTTCTTCATCTTC | |
| R: CTCGGGTGTCTTGTCGCTTTGG | ||
|
| F: AACCCAGACATCTTTGCCGTGATC | |
| R: CCATTATGAAGCCCAGGACCAGAA | ||
|
| F: GCACTCGGACCTCCTTCACAAG | |
| R: AGGATGACAGCGACAATGACAGC | ||
|
| F: TACTTGGTTCTGCTGGTGTCC | |
| R: GTTGTGGGCTGTGTAAAGGTG | ||
|
| F: CACTGACTCCTCCGACCTAC | |
| R: TACTGCGTGACCAGGAAACT | ||
|
| F: CAGGTGCAGGTCCTCACTTCAATC | |
| R: GTCACATTGCCCAGGTCTCCAAC | ||
|
| F: TTCTGGACAAATCTGAGCCCTAACG | |
| R: CGACGGATACAGCGGTCAACTTC | ||
|
| F: TGCAACCAATTTGGACATCAG | |
| R: TTCACGTCACACTTCTGGATAAGG | ||
|
| F: AAACAGGAACCGGGAGACAA | |
| R: AGGACAGGCGTTCTTCAGGAA | ||
|
| F: TCCATGCACGAATTCTCAGCCAAG | |
| R: TCATTGAGAGGCCACATTGGTGAC | ||
|
| F: TGCCCATACTTCCCGTCC | |
| R: GGTCCAGGTTACCGTCAG | ||
|
| F: TCTGCCCTGTACCCCAACTG | |
| R: CCAGGAAGACGGGCTTTTG | ||
|
| F: TCAGTCCAGTCGCCTTCTCC | |
| R: GGCATTTGTGGTGGGGTTAG | ||
|
| F: AAATACGCATTCCACACCTTTCCAC | |
| R: TGCTGTTGTTGTTGCTTCTCAGTTC | ||
|
| F: ATGGGCGACTTGTTGCTGAC | |
| R: CACAGGGCAGAAATTGATGACA | ||
|
| F: CCAACATGACCAAGCCTACCACA | |
| R: ACAAGTTTGCCTCCATCCAGTGT | ||
|
| F: GGAGTAGAGGGCAAAGCAGG | |
| R: AGGTCTGGCGTGGGTCAAAG | ||
|
| F: AGGATTGTGGCCTTCTTTGAGTT | |
| R: CGGTTCAGGTACTCAGTCATCCA | ||
|
| F: GGTACCGGAGGGCATTCAG | |
| R: ACAATGCGACCCCAGTTCAC | ||
|
| F: GCTGCTGCTCCTGCTTCTAGTG | |
| R: AGGTGAATTCCTTGCACGGTCTG | ||
|
| F: CCTGCTGCTTCACATACA | |
| R: TTCAGTTCCAAGTCATCCAT | ||
|
| F: CCAGACCTCCAACAACATT | |
| R: CGGAAGAACCTGCTCAAG | ||
|
| F: CAGGATGTGAACGAATGC | |
| R: AGAGATAATGGAAGGAGATGG | ||
|
| F: GCCAACCAACTCTTCTGTA | |
| R: CGATTGAATGCCACTCTTG | ||
|
| F: GATGGAAGGCTCTGATACTG | |
| R: GGACCGAAGACCTCTGTT | ||
|
| F: ACTGGAAGAGAAGATAGATGTG | |
| R: GAGCGGATGAAGGTGAAG | ||
|
| F: GTAGTGTATGAGCGTGTGA | |
| R: GGTCTGTGGATATGAAGTGA | ||
|
| F: AAACAGGAACCGGGAGACAA | |
| R: AGGACAGGCGTTCTTCAGGAA |
| Group | CON Group ( | OPI Group ( | |
|---|---|---|---|
| Number of piglets born | 15.27 ± 3.87 | 15.81 ± 3.62 | 0.49 |
| Number of live piglets | 14.02 ± 3.54 | 14.23 ± 3.01 | 0.76 |
| Litter weight of live piglets | 17.42 ± 4.24 | 18.21 ± 3.87 | 0.35 |
| Average weight of live piglets | 1.27 ± 0.25 | 1.30 ± 0.22 | 0.61 |
| Number of stillborn piglets | 1.00 ± 1.41 | 1.23 ± 1.52 | 0.46 |
| Number of mummified fetuses | 0.27 ± 0.82 | 0.35 ± 0.64 | 0.59 |
| Number of weak piglets | 2.02 ± 2.31a | 1.23 ± 1.39b | 0.05 |
| Number of healthy piglets | 12.00 ± 2.94 | 13.00 ± 2.65 | 0.09 |
| Rate of healthy piglets/% | 87.10 ± 12.97a | 92.01 ± 8.79b | 0.04 |
| Group | CON Group (n = 44) | OPI Group (n = 48) | |
|---|---|---|---|
| Number of live piglets at 7 days | 11.30 ± 2.53 | 12.08 ± 2.20 | 0.10 |
| Litter weight of live piglets at 7 days | 29.45 ± 7.54a | 32.75 ± 6.77b | 0.03 |
| Average weight of live piglets at 7 days | 2.65 ± 0.62 | 2.71 ± 0.34 | 0.56 |
| Daily weight gain of live piglets at 7 days | 0.20 ± 0.07 | 0.20 ± 0.04 | 0.66 |
| Survival rate of piglets at 7 days | 82.33 ± 13.80 | 86.33 ± 10.57 | 0.13 |
| Group | CON group ( | OPI group ( | |
|---|---|---|---|
| Number of live piglets at 14 days | 10.61 ± 2.37a | 11.63 ± 1.85b | 0.03 |
| Litter weight of live piglets at 14 days | 43.98 ± 10.67a | 49.49 ± 10.57b | 0.02 |
| Average weight of live piglets at 14 days | 4.16 ± 0.52 | 4.26 ± 0.66 | 0.42 |
| Daily weight gain of live piglets at 14 days | 0.21 ± 0.03 | 0.21 ± 0.05 | 0.50 |
| Survival rate of piglets at 14 days | 77.81 ± 15.14a | 83.45 ± 11.91b | 0.05 |
| Group | CON Group ( | OPI Group ( | |
|---|---|---|---|
| Number of live piglets at 21 days | 10.32 ± 2.35a | 11.44 ± 1.77b | 0.01 |
| Litter weight of live piglets at 21 days | 59.22 ± 16.40a | 67.81 ± 14.46b | 0.01 |
| Average weight of live piglets at 21 days | 5.75 ± 0.88 | 5.95 ± 0.97 | 0.31 |
| Daily weight gain of live piglets at 21 days | 0.21 ± 0.04 | 0.22 ± 0.04 | 0.32 |
| Weaning survival rate at 21 days | 75.74 ± 15.69a | 82.22 ± 12.08b | 0.03 |
- —National Key R&D Program of China
- —Hebei Agriculture Research System
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Nutrition and Physiology · Aquaculture Nutrition and Growth · Insect Utilization and Effects
Introduction
1
The utilization of contemporary animal breeding and reproduction techniques has led to a substantial augmentation in the number of offspring per litter of sows. However, this development is concomitant with a series of challenges, including, but not limited to, low birth weight, a high incidence of weak piglets, a low piglet survival rate, high morbidity, and a decline in litter weight at weaning (1–3). The intake of colostrum and milk was lower in low-birth-weight piglets than in high-birth-weight piglets within the same litter (4). This had a detrimental effect on the growth, reproductive, and quality of life of the piglets (5). Researchers have confirmed that the material exchange between sows and fetuses intensifies during the late stages of gestation, and nutritional intake is crucial for fetal growth (6). Nutrient transfers of the conventional kind, such as amino acids and sugars, are inadequate in supplying fetal growth and development (7). Additionally, it has been shown that oxidative stress, a possible consequence of pregnancy, can lead to an increased incidence of low-birth-weight piglets (8). Therefore, adequate nutrition and antioxidant products supplement during the late gestation is essential for the health of the sow and the number of viable offspring (9).
Peptides have been demonstrated to be remarkably effective in promoting reproduction and enhancing litter performance, and they do not interfere with maternal protein uptake pathways, rendering them optimal nutritional supplements (10). Oyster peptide (OPI) has been demonstrated to possess functional activities that enhance immunity, antioxidant and anti-inflammatory properties (11). It has been reported that OPI exhibited high antioxidant activities and could promote cell growth (12–14). Furthermore, OPI effectively enhanced cell proliferation, differentiation, and mineralization in MC3T3-E1 cells (15). Other studies have indicated that OPI possesses anti-inflammatory properties and has the capacity to impede the expression of inflammatory cytokines, including TNF-α, IL-1β and IL-6, contributing to an enhancement in the body’s immune system (16, 17). We speculated that dietary OPI supplement to sows in the late gestation can reduce the oxidative stress level, enhance immunity, and strengthen the nutrient transport between sows and fetus. Therefore, the objective was to investigate the impact of incorporating OPI into the diets of sows during the late gestation and lactation phases on litter performance, immunological response of sows and the growth and development of piglets.
Materials and methods
2
Experimental materials
2.1
The OPI utilized in this experiment was obtained from the Biological Synthesis of Nutritional Resources Research Group, Tianjin Institute of Industrial Biotechnology, Chinese Academy of Sciences (18). During application, it was added at the recommended dosage to ensure the accuracy and standardization of the experimental conditions.
Experimental animals and design
2.2
This experiment was performed in September 2023 at Ruijia Agricultural Development Co., Ltd., Yongqing County, Langfang, Hebei Province, China. A total of 100 Canadian Landrace × Yorkshire sows with healthy hooves and healthy body conditions were selected on the 90th day of pregnancy. The selected sows were randomly divided into two groups: the control group (CON group) was fed a basic diet, while the experimental group received the basal diet supplemented with 2 mg/kg of OPI. The feeding period was from 90 days of pregnancy to 21 days post-farrowing, with 50 sows in each group. Base diets for sows in pregnancy and lactation, respectively, were selected according to the nutritional requirements of different periods (Table 1).
Feeding management
2.3
Prior to farrowing, the sows in each group were kept in pens. One week ante partum, the animals were moved into the farrowing unit and all allocated to individual farrowing pens. The environment was kept clean and dry, with regular ventilation, lighting and temperature control at 25 °C, and disinfection procedures carried out regularly. The pregnant sows were fed twice a day at 5:00 and 16:30, with each feeding event involving the provision of 1.5 kg of feed. On the day of farrowing, the pregnant sows were provided with a reduced amount of feed (1.2 kg/meal, twice a day), and feeding was then gradually resumed 1–2 days after giving birth (1.3 kg/meal, twice a day). The nursing sows were fed three times a day at 5:00, 10:30, and 16:30, with each feeding consisting of 2 kg. All sows had unrestricted access to clean drinking water to maintain their normal physiological functions.
Sample collection
2.4
During parturition, seven sows per group were randomly selected for sample collection. During sow farrowing, umbilical cords were tied with a short silk line and each piglet was marked with a numbered tag to match the individual piglets with their placentae. After placental expulsion and weight recording, the placentae were collected and snap-frozen in liquid nitrogen (3 to 4 cm from the cord insertion point) and then stored at −80 °C until analysis.
Colostrum samples were taken when the first piglet was expelled. Before colostrum sampling, the entire visible udder was cleaned with soap and water and dried with paper. Subsequently, colostrum was collected through manual milking by massaging evenly, and then stored at −80 °C until analysis (19).
Data measurement of sows and piglets
2.5
On delivery day, the litter size, numbers of born alive, stillborn piglets, mummified fetuses, and individual piglet weight of each sow were recorded. The weak and healthy piglets were identified by the birth weight. Birth weight of healthy piglet was more than 900 g, while birth weight of weak piglet was less than 900 g (20). On 7, 14, and 21 days in lactation period, the individual piglet weight was recorded. The survival rate was calculated by dividing the number of live piglets by the total number of piglets born.
Analysis of immunoglobulin levels
2.6
The immunoglobulin (IgA, IgE, IgG, IgM) concentrations in the colostrum of farrowing sows was detected by using an enzyme-linked immunosorbent assay (ELISA). The assay procedure was conducted according to the instructions provided in the relevant porcine ELISA kit manual (Jingmei, China). The specific operation was as follows: firstly, the standard sample gradient dilution treatment was conducted, and the standard sample and the sample to be tested were added to the enzyme standard coated plate. The plate was then incubated at 37 °C for 30 min. It was subsequently washed five times by washing solution. Subsequently, the enzyme reagent was added and the plate was incubated at 37 °C for a further 30 min. The plate was then washed on five separate occasions. Finally, the color development solution was added, and the plate was incubated at 37 °C for 10 min. The absorbance at 450 nm was read with an UV spectrophotometer.
RNA sequencing
2.7
Three samples of placentas from the same sow were randomly selected in each group for transcriptomic analysis. Firstly, the total RNA of the samples was extracted using a Trizol kit (Biomed, China) and the concentration and quality of the total RNA were detected using the NanoDrop 2000 (Thermo, Waltham, MA, United States). After enriched mRNA was subsequently reverse transcribed to cDNA, and cDNA ends were repaired, purified, and amplified. The resulting cDNA libraries were used with Novaseq 6000 (Illumina, United States) to obtain quantification and annotation information for all genes.
For the analysis of RNA sequencing, first, the clean reads were obtained by removing reads containing adapter ploy-N and low-quality reads from the raw data (raw reads). Next, TopHat2 software was used to map reads to the pig genome (Sscrofa 11.1). Cuffquant and cuffnorm (v2.2.1) were used to estimate the expression levels of all the transcripts and analyze expression levels for mRNAs by calculating FPKM. The various differentially expression genes (DEGs) were selected between the 2 groups by using the DESeq2 R package when the adjusted FDR <0.05 and the absolute value of the log2 (fold change) was larger than 1.3. The volcano plot and heatmap of DEGs were clustered using TBtools (version 1.046). Gene Ontology (GO, http://geneontology.org/) enrichment analysis of the DEGs was performed to assess their biological significance using the GOseq R package.
Real-time quantitative PCR (RT-qPCR)
2.8
RNA samples were the same with those used in transcriptomic analysis. QRT-PCR with SYBR fluorescent dye (Biotium, Bay Area, CA, United States) was used to detect the mRNA expression levels of the genes in the cells, and the PCR conditions were as follows: 95 °C for 2 min followed by 40 cycles of 95 °C for 5 s and 60 °C for 30 s. Each experiment was repeated four times. Finally, gene expression was quantified using the 2^−ΔΔCT^ method, and GAPDH was selected as housekeeping gene. Primers used to amplify each gene are listed in Table 2.
Statistical analysis
2.9
The data analysis of this experiment was conducted using SPSS (Version 25.0, IBM Corporation, Armonk, NY, United States). First, all data were detected normally distributed. Independent sample t-test was used for the comparison of number and weight of piglets between two groups, while the chi-square test for the comparison of rate of healthy piglets and survival rate of live piglets between two groups. All results were expressed as the mean ± standard deviation. A significance level of p ≤ 0.05 was established to indicate a statistically significant difference.
Results
3
Effects of dietary supplementation of OPI during late pregnancy on the farrowing performance of sows
3.1
As demonstrated in Table 3, the supplementation of OPI of sows during the late gestation period resulted significant decrease in the number of weak piglets (2.02 ± 2.31 vs. 1.23 ± 1.39, p ≤ 0.05). Rate of healthy piglets was significantly promoted in OPI group (92.01 ± 8.79 vs. 87.10 ± 12.97, p ≤ 0.05).
Effect of OPI on growth performance of piglets during the suckling period
3.2
Dietary OPI supplementation of sows could significantly increase litter weight of live piglets at 7 days (29.45 ± 7.54 vs. 32.75 ± 6.77, p ≤ 0.05) (Table 4). As can be observed from Table 5, the number of live piglets at 14 days of the OPI group (10.61 ± 2.37 vs. 11.63 ± 1.85), litter weight of live piglets (43.98 ± 10.67 vs. 49.49 ± 10.57) and the survival rate of piglets at 14 days (77.81 ± 15.14 vs. 83.45 ± 11.91) were all significantly increased compared to the CON group (p ≤ 0.05). It could be found from Table 6 that OPI supplementation could significantly increase the number of live piglets weaned at 21 days (10.32 ± 2.35 vs. 11.44 ± 1.77, p ≤ 0.05), litter weight of live piglets at 21 days (59.22 ± 16.40 vs. 67.81 ± 14.46, p ≤ 0.05) and weaning survival rate (75.74 ± 15.69 vs. 82.22 ± 12.08, p ≤ 0.05) compared to the CON group (p ≤ 0.05).
Effect of OPI on immunoglobulin levels in the colostrum of sows
3.3
As shown in Figure 1, the OPI supplementation could significantly increased the concentrations of IgA, IgG, and IgM in the colostrum of farrowing sows compared with the CON group (p ≤ 0.05).
*Effects of adding oyster peptide to the diet of sows in late pregnancy on the levels of immunoglobulin in sow’s colostrum. (A) The level of IgA in colostrum. (B) The level of IgE in colostrum. (C) The level of IgG in colostrum. (D) The level of IgM in colostrum. *Represents significant difference at p ≤ 0.01.
Expression of transporter carriers, antioxidant and immune genes in placenta of sows supplemented with OPI
3.4
The expression levels of glucose transporter carriers (GLUT4, SLC2A1), amino acid transporter carriers (SNAT1, LAT1), fatty acid transporter carriers (FABP3) and antioxidant genes (SOD1, SOD2, GPX2, CAT) in the placenta was significantly increased compared to the CON group (Figures 2A–D, p ≤ 0.05). Anti-apoptotic genes (BCL2, BCL2L1) were also significantly higher expressed in the OPI group (Figure 2E, p ≤ 0.05). As shown in Figure 2F, the OPI supplementation significantly decreased the expression level of pro-inflammatory factors (IL-1, IL-6, IL-8, AVPI1) and significantly increased the expression levels of anti-inflammatory factors (IL-10) in sow placenta (p ≤ 0.05).
*Effects of adding oyster peptide to the diet of sows in late pregnancy on the function of placenta. (A) The expression level of placental glucose related genes. (B) The expression level of placental amino-acid related genes. (C) The expression level of placental fatty -acid related genes. (D) The expression level of placental antioxidant-related genes. (E) The expression level of placental anti-apoptosis-related genes. (F) The expression levels of inflammatory cytokines within the placenta. *Represents significant difference at p ≤ 0.05. *Represents significant difference at p ≤ 0.01.
Transcriptomics analysis of placenta
3.5
Based on the PCA analyses of transcriptomics, the profiles in the OPI group were distinct from those in the CON group (Figure 3A). A total of 13,215 known genes were identified in the CON group of this experiment, of which 847 were uniquely expressed. A total of 12,712 known genes were identified in the OPI group, of which 344 were uniquely expressed (Figure 3B). There was a total of 730 differentially expressed genes (DEGs) between the two groups, including 111 up-regulated genes and 619 down-regulated genes (Figure 3C) in OPI group compared to CON group. The heatmap demonstrated the top 20 DEGs (Figure 3D).
Analysis of differentially expressed genes (DEGs) in RNA-seq. (A) Principal component confidence diagram analysis of OPI and CON groups. (B) mRNAs expression statistics. (C) Volcano plot of differentially expressed genes (DEGs) between OPI and CON groups. (D) Heatmap of top 20 DEGs.
In order to further explore the molecular mechanism in the function of OPI, GO and KEGG analysis of DEGs were performed. GO analysis demonstrated that the DEGs were predominantly associated with skeletal system development, regulation of multicellular organismal process, connective tissue development and plasma membrane (Figure 4A). The majority of the DEGs were enriched in cellular process, metabolic process, biological regulation (biological processes); binding, transporter activity, molecular transducer activity, molecular function regulator (molecular functions) and composition of cell, membrane and macromolecular complex (cellular components) (Figure 4D).
Enrichment analysis and verification of DEGs. (A) GO enrichment analysis of DEGs, value in X-axis represented the enrichment of GO terms. (B) KEGG enrichment analysis of DEGs, value in X-axis represented the enrichment of pathway. (C) KEGG significance bubble chart. (D) GO enrichment functional classification diagram. (E) KEGG quantity statistical chart. (F) Expression validation of 8 randomly selected DEGs by q-PCR. Y-axis represented the log2 FC (RNA-seq) or relative mRNA expression to GAPDH (q-PCR).
KEGG enrichment analysis demonstrated that the DEGs were predominantly associated with metabolism signaling and cortisol synthesis, secretion, including metabolic pathways, protein digestion, parathyroid hormone synthesis and ABC protein transport (Figures 4B,C,E).
As demonstrated in Figure 4F, randomly selected eight DEGs (CXCL10, CCL4, DNAJB1, FBLN7, SATB2, SGK2, IL1RN, POF1B) were subjected to qRT-PCR for validation in the CON and OPI groups. There were similar directions of the fold change of relative expression level in qRT-PCR and RNA-seq of the genes. This result verified the reliability of the sequencing data.
Discussion
4
Fetus grows and develops fastest in sows in the late gestation period, with significant enhancements in energy, protein and fat (21). Previous research has revealed bioactive ingredient can enhance the growth performance, by improving digestive function, inflammation resistance and the enhancement of antioxidant properties (22, 23). Another study revealed that the supplementation of N-carbamoyl-aspartic dipeptide into the diet of sows resulted in an augmentation of the litter weight of piglets, in addition to an increase in the survival rate of the piglets (18). The OPI produced from pichia pastoris was used in this study. It is a mixture, and its purity is not measured by the content of peptides, but by its antioxidant performance. The mixture is abundant in protein, taurine, glycogen and other nutrients. Since the dosage of OPI was 2 mg/kg, the protein and other nutrients in it had very little impact on the nutritional components of the feed, thus OPI is more regarded as an active additive.
However, there was no research about OPI supplementation on the litter performance, immunological response in sows, the addition of OPI resulted in a significant increase in the number of healthy litters (birth weight of healthy piglet ≥900 g), the number of live weaned piglets, the weaning litter weight and the weaning survival rate in this study as we expected. The function of OPI, enhancing immunity, antioxidant and anti-inflammatory properties, showed in this study was consistent with the previous research (12, 13).
The placenta, an organ that connects the fetus to the mother, plays a crucial role in facilitating fetal nutritional intake (24). Ren’s study demonstrated that the incorporation of the active peptide CC34 led to a substantial augmentation in the relative expression of sheep amino acid transporter carriers (25). The addition of the honeybee antimicrobial peptide Api-PR19 was indicated by Wang (26) to significantly increase the expression levels of glucose and fatty acid transporter carriers. The results in this study demonstrated that OPI could promoted the expression of glucose (GLUT4, SLC2A1), amino acids (SNAT1, LAT1), and fatty acid (FABP3) transporter carriers in the placenta of sows. Concurrently, the number of healthy piglets increased, while the number of weak piglets decreased. Therefore, OPI could help improve the production performance of sows through its nutritional components and improving placental transport function.
During the late pregnancy period, the maternal immune system is reduced and vulnerable to inflammatory factors. Inflammation is a basic pathological process that occurs in the body as a defense response to various damaging factors (such as infection, trauma, foreign bodies, etc.) (27, 28). It has been reported that the immune system of piglets was shaped by the placenta during gestation and further developed through colostrum after birth (29–31). Hwang et al. (17) found that OPI alleviated the inflammatory response induced by lipopolysaccharide and significantly reduced the expression level of pro-inflammatory factors by inhibiting the NF-κB pathway. Qian et al. (16) have reached similar conclusions, asserting that OPIs possess anti-inflammatory properties and are capable of alleviating the oxidative stress caused by lipopolysaccharides on macrophages. As demonstrated by reports such as the above, it can be concluded that OPI exerts some degree of anti-inflammatory effects. Our findings also showed a significant increase of anti-inflammatory factors (IL-10) in the placental with OPI supplementation. Furthermore, there is a substantial increase in immunoglobulins in the colostrum of sows, thereby enhancing the immunity of piglets and increasing survival rates. This might be due to the fact that the overall immune level of sows has been improved after OPI supplementation, which led to an increase in the level of immunoglobulin in colostrum.
Oxidative stress becomes more pronounced during late gestation and lactation of sows and piglets and did not recover until after weaning (32). It is directly associated with a rise of stillbirths and weak piglets (33). It has been established that elevated ROS could induce autophagy, dysfunction and apoptosis in vascular endothelial cells, leading to impaired development of the placental vasculature system and resulting in intrauterine fetal developmental restriction (34). Wang et al. (35) observed that administration of OPI to mice resulted in a significant decrease in ROS and a significant increase in SOD and GSH-Px, thereby significantly enhancing the antioxidant properties of the mice. Yan et al. (13) indicated that OPI may exert antioxidant effects by binding to ROS and interacting with antioxidant enzymes. In this study, the addition of OPI could increase SOD, GSH-Px and CAT expression levels, which was consistent with the previous research. This demonstrates OPI enhances the antioxidant properties of the placenta in sows, and this may be an important reason for the increase of healthy piglets.
In order to investigate the mechanism of OPI, q-PCR and transcriptome of placenta were analyzed. The pro-inflammatory factors (IL-1, IL-6, IL-8) were decreased while expression of anti-apoptotic genes (BCL2, BCL2L1) were increased in OPI group. In addition, key genes associated with placental transport and anti-inflammatory were identified. These results provided potential mechanisms of OPI on sow reproductive performance at the mRNA level.
Conclusion
5
Dietary OPI in sows during late gestation and lactation is a promising way to improve litter performance, immunological response of sows and growth performance of piglets. OPI supplementation could increase the expression of antioxidant and nutrient transport gene in placental, which may lead to better piglet birth weight and more healthy piglets. Moreover, it could also enhance immune function both in sows and colostrum, strengthening piglet passive immunity, and improving weaning litter weight and survival rate.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wu X Gao LM Liu YL Xie C Cai L Xu K. Maternal dietary uridine supplementation reduces diarrhea incidence in piglets by regulating the intestinal mucosal barrier and cytokine profiles. J Sci Food Agric. (2020) 100:3709–18. doi: 10.1002/jsfa.10410, PMID: 32248539 · doi ↗ · pubmed ↗
- 2Li M Chen L Zhao Y Sun H Zhao L. Research on the mechanism of HRP relieving IPEC-J 2 cells immunological stress based on transcriptome sequencing analysis. Front Nutr. (2022) 9:944390. doi: 10.3389/fnut.2022.944390, PMID: 35911118 PMC 9336541 · doi ↗ · pubmed ↗
- 3Zhao L Yu J Liu Y Liu Y Zhao Y Li MY. The major roles of intestinal microbiota and TRAF 6/NF-κB signaling pathway in acute intestinal inflammation in mice, and the improvement effect by Hippophae rhamnoides polysaccharide. Int J Biol Macromol. (2025) 296:139710. doi: 10.1016/j.ijbiomac.2025.139710, PMID: 39793780 · doi ↗ · pubmed ↗
- 4König NLWähner M Seeger J Sigmarsson HL Kauffold J. An investigation into uterine capacity based on litter and placental characteristics in two sow lines with different prolificacy (Danish Landrace × Danish Yorkshire versus German Saddleback). Reprod Domest Anim. (2021) 56:34–45. doi: 10.1111/rda.13847, PMID: 33098192 · doi ↗ · pubmed ↗
- 5Rutherford K Baxter ED’Eath R Turner S Arnott G Roehe R. The welfare implications of large litter size in the domestic pig i: biological factors. Anim Welf. (2013) 22:199–218. doi: 10.7120/09627286.22.2.199 · doi ↗
- 6Mc Gee M Bainbridge S Fontaine-Bisson B. A crucial role for maternal dietary methyl donor intake in epigenetic programming and fetal growth outcomes. Nutr Rev. (2018) 76:469–78. doi: 10.1093/nutrit/nuy 006, PMID: 29529267 · doi ↗ · pubmed ↗
- 7Gormley A Jang KB Garavito-Duarte Y Deng Z Kim SW. Impacts of maternal nutrition on sow performance and potential positive effects on piglet performance. Animals. (2024) 14:1858. doi: 10.3390/ani 14131858, PMID: 38997970 PMC 11240334 · doi ↗ · pubmed ↗
- 8Hu C Yang Y Deng M Yang L Shu G Jiang Q. Placentae for low birth weight piglets are vulnerable to oxidative stress, mitochondrial dysfunction, and impaired angiogenesis. Oxid Med Cell Longev. (2020) 2020:8715412. doi: 10.1155/2020/8715412, PMID: 32566107 PMC 7267862 · doi ↗ · pubmed ↗