Bacterial skin colonization with a specific Cutibacterium avidum clade as a risk factor for periprosthetic joint infections—a multicenter study
Llanos Salar Vidal, Julia Prinz, Pascal M. Frey, Tiziano A. Schweizer, Laura Böni, Silvio D. Brugger, Holger Brüggemann, Jaime Esteban, Yvonne Achermann

TL;DR
A specific group of Cutibacterium avidum bacteria found on the skin is linked to joint infections after surgery, suggesting these strains may be more likely to cause disease.
Contribution
This study identifies a distinct phylogenetic clade of Cutibacterium avidum associated with periprosthetic joint infections and reveals shared biofilm and antibiotic resistance traits.
Findings
All periprosthetic joint infection isolates belong to a single phylogenetic clade of Cutibacterium avidum.
PJI isolates show reduced bacterial fitness compared to skin isolates, potentially contributing to infection persistence.
Rifampin is the most effective antibiotic against Cutibacterium avidum biofilms and growth.
Abstract
Cutibacterium avidum is increasingly recognized as a causative agent of periprosthetic joint infections (PJIs), yet data on its pathogenic potential and distinguishing features from commensal strains remain limited. In this multicenter study, we compared 11 C. avidum isolates from PJIs with 32 isolates from healthy skin collected across four European hospitals. We investigated phylogenetic relationships, antibiotic susceptibility, biofilm formation, and bacterial fitness. Phylogenomic analysis revealed two main clades within the C. avidum population. All PJI isolates belonged exclusively to Clade 1, which also included skin isolates. Within Clade 1, gene content analysis showed no consistent genetic differences between PJI and skin isolates. All isolates exhibited moderate to strong biofilm formation, with no significant differences in either data set. Minimal inhibitory concentration…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
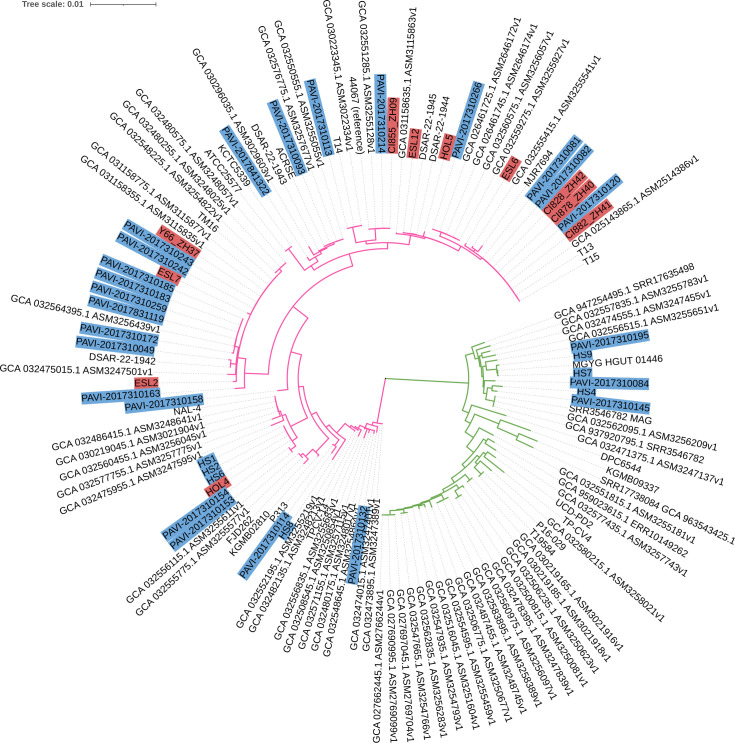 Fig 1
Fig 1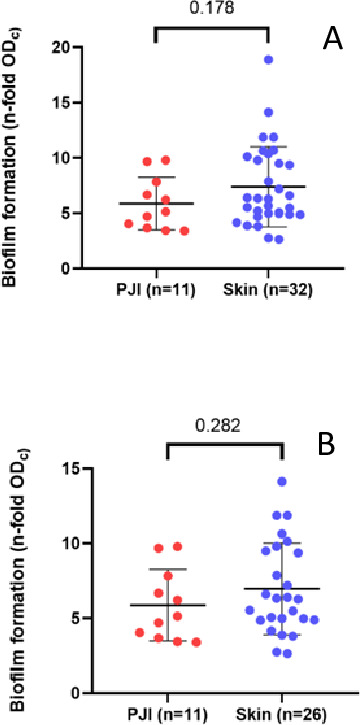 Fig 2
Fig 2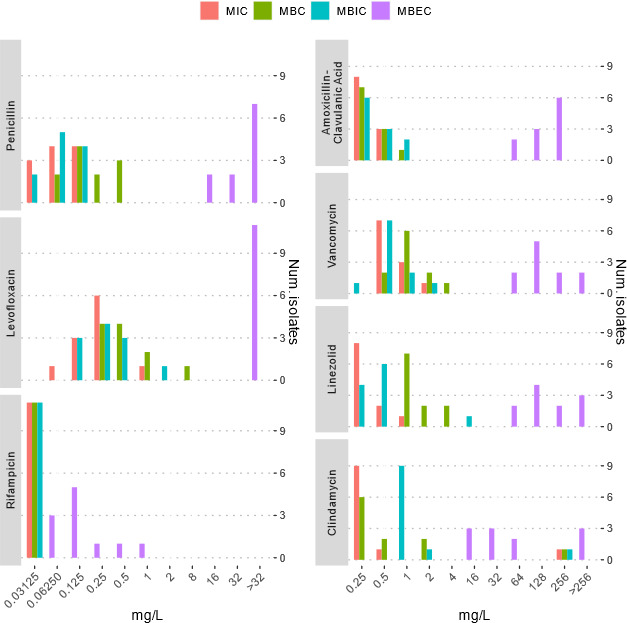 Fig 3
Fig 3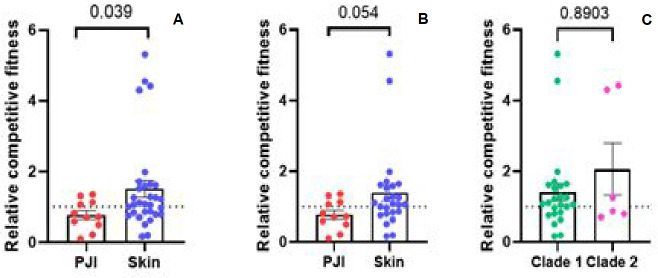 Fig 4
Fig 4- —Jubiläumsstiftung von Swiss Lifehttp://dx.doi.org/10.13039/501100022796
- —Centro de Investigación Biotecnológica en Red de Enfermedades Infecciosas (CIBERINFEC)
- —Fabrikant Vilhelm Pedersen og Hustrus Legat (VPL)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsOrthopedic Infections and Treatments · Bacterial Identification and Susceptibility Testing · Antimicrobial Resistance in Staphylococcus
INTRODUCTION
Infections related to arthroplasty cause high morbidity and also impose an increased cost to the healthcare system. The most common bacteria isolated from prosthetic joint infections (PJIs) are staphylococci, followed by streptococci, enterococci, gram-negative bacilli, and anaerobes (1, 2).
Even though anaerobic bacteria make up only approximately 3–6% of PJI (3), they can cause more than 50% of infections in shoulder implants (4). Among them, more than 70% are gram-positive bacteria belonging to Cutibacterium spp. (5) with Cutibacterium acnes monopolizing the attention (6). However, another species from this genus, Cutibacterium avidum, is also gaining importance (6–8).
C. avidum is a member of the human skin microbiota. It tends to reside in wet areas such as the axilla, nares, groin, and rectum (7). It is considered a skin commensal with low virulence potential; however, it can act as an opportunistic pathogen in superficial and deep/invasive infections such as skin abscesses, abdominal infections, breast infections, infective endocarditis, prostate infections, and bone and joint infections (8–11). C. avidum is an infrequent etiological agent of PJI, predominantly in late chronic infections. It preferentially colonizes moist areas; hence, mainly obese individuals who underwent primary hip arthroplasty were found to be affected (8, 12, 13).
Many different virulence factors may be involved in PJI, including biofilm formation (7). However, very little information is currently available about the role of C. avidum biofilms in PJI. Therefore, we investigated antibiotic susceptibility and virulence properties such as biofilm formation and fitness of C. avidum isolates recovered from deep tissues in PJI patients and compared them to isolates from healthy skin. We performed whole-genome sequence analysis of the core genome to examine the phylogenetic relationship between the cohort.
MATERIALS AND METHODS
Strain isolation and identification
The study (research project involving biological material and health-related personal data, clinical trial number: not applicable) includes 43 C. avidum isolates, either recovered from PJI (n = 11) from four European hospitals as part of a multicenter study supported by the European Study Group for Implant-Associated Infections of the European Society of Clinical Microbiology and Infectious Diseases or taken from healthy skin (HS) volunteers (n = 32) by scraping the skin with sterile blades. Skin scrapings were removed from blades and transferred into ESwab culture swabs (Copan, Brescia, Italy) (13). Swabs were cultured onto Schaedler-5% sheep blood agar plates (BioMérieux, Marcy l’Étoile, France) for 48 h at 37°C under anaerobic conditions and isolates were identified by MALDI-TOF MS (Vitek MS, BioMérieux, Marcy l’Étoile, France). Clinical data of C. avidum isolates recovered from PJI are shown in Table S1.
Biofilm formation
Biofilm formation of all PJI and skin isolates was evaluated using a modified method of Stepanović et al. (14) as a static biofilm assay. Briefly, brain heart infusion (BHI) broth with 2% glucose was used with bacterial inocula of 10^7^ CFU/mL and incubated at 37°C under anaerobic conditions in 96-well plates. After 72 h of incubation, BHI broth was removed, and the wells were rinsed two times with methanol, and crystal violet was used for staining. The optical density (OD) of each well was measured at 570 nm using a microtiter plate reader. Each isolate was tested in triplicate. Isolates were divided into different categories based on OD values and a cut-off value (ODc) was established. The results were interpreted as: no biofilm producer (OD ≤ ODc), weak biofilm producer (ODc < OD ≤ 2 × ODc), moderate biofilm producer (2 × ODc < OD ≤ 4 × ODc), or strong biofilm producer (4 × ODc < OD).
Susceptibility testing
Antibiotic susceptibility testing of 11 PJI-causing C. avidum isolates was performed. Antibiotics tested were amoxicillin-clavulanate, clindamycin, levofloxacin, linezolid, penicillin, rifampin, and vancomycin. The minimal inhibitory concentration (MIC) and the minimal bactericidal concentration (MBC) were determined with the broth microdilution method according to the protocols of the European Committee on Antimicrobial Susceptibility Testing (EUCAST) (15). Colonies of the isolates were suspended in Mueller Hinton cation-adjusted broth. Sterile round-bottomed 96-well plates were inoculated with 100 µL of Mueller-Hinton cation-adjusted broth containing the antimicrobial agent plus 100 µL of the bacterial suspension for obtaining a final inoculum of 10^4^ CFU per well and incubated under anaerobic conditions. After 48 h of incubation, the MIC was determined (the first antibiotic concentration where there was no turbidity). Subsequently, 50 µL of each well was transferred into a new plate containing 150 µL of Mueller Hinton cation-adjusted broth by using a modified flash microbiocide method (16). Plates were incubated for 2 days at 37°C under anaerobic conditions, and the MBC was determined (the first antibiotic concentration where there was no bacterial growth).
The minimal biofilm inhibitory concentration (MBIC) and the minimal biofilm eradication concentration (MBEC) were assessed following the protocols previously described by Coenye et al. (17). Briefly, colonies of the isolates were transferred into sterile phosphate-buffered saline (PBS), and the supernatant was adjusted to a turbidity of 0.5 ± 0.02 McFarland. A volume of 200 µL of the bacterial suspension was transferred into each well of a sterile, flat-bottomed, polystyrene 96-well plate tissue cultured. Plates were incubated in anaerobiosis jars (Oxoid Ltd., Thermo Fisher Scientific, Boston, MA, USA) to allow bacterial adhesion to the well bottom. Following 4 h of adhesion, the supernatant, containing the planktonic bacteria, was carefully removed. Each well was washed with 200 µL of sterile PBS to remove non-adherent cells. Then, 200 µL of clostridial nutrient medium (Sigma-Aldrich, Missouri, USA) was added, and the plates were incubated for 24 h at 37°C under anaerobic conditions to allow biofilm maturation. After incubation, the supernatant was again removed, and the plates were washed with 200 µL of sterile PBS. Each well was then filled with 100 µL of clostridial nutrient medium plus 100 µL of clostridial nutrient medium containing serial dilutions of the antimicrobial agent, and plates were further incubated. After 48 h of incubation in anaerobic conditions, the MBIC was determined (the first antibiotic concentration where there was no turbidity). After the MBIC assessment, the wells were washed with 200 µL of sterile PBS and each well was filled with 200 µL of fresh clostridial nutrient medium. The biofilm was mechanically disrupted by vigorously scraping the well bottom using sterile pipette tips, followed by homogenization. The MBEC was determined after 48 h of incubation (the first antibiotic concentration, where there was no bacterial growth).
The EUCAST resistance breakpoints for C. acnes were used to interpret antimicrobial susceptibility results (18). Levofloxacin and rifampin breakpoints have not been determined for anaerobic gram-positive bacteria.
Statistical analysis was performed using GraphPad Prism 8.4.3 (GraphPad Software, San Diego, CA, USA). Data were evaluated using a Wilcoxon nonparametric test to compare two groups. Statistical significance was set at P values ≤ 0.05. Figure 3 was generated using R (19).
Bacterial quantitative fitness analysis (BaQFA)
To assess reproductive fitness variations among C. avidum isolates, the BaQFA method was used (20). This approach involved spotting 96 bacterial cultures on an agar plate and capturing their growth over time through time-lapse photography with a BaQFA robot using an Arduino platform (21).
Eleven invasive isolates recovered from PJI and 31 superficial skin isolates were tested against one superficial skin strain as reference strain to analyze the difference in fitness. C. avidum strain PAVI-2017310081 was used as reference strain for all BaQFA experiments. C. avidum isolates were streaked out on Columbia sheep blood agar plates and incubated at 37°C under anaerobic conditions for 3 days. Fresh bacterial colonies were scraped from the agar plates and diluted in PBS to an OD_600nm_ of 0.10 (±0.01). The bacterial solution was diluted 1:10 in PBS, and 3 µL spots was spotted onto a rectangular single-well BHI agar plate with 20 mL agar medium in a grid pattern (each tested strain was grown in direct neighborhood of the competing reference strain). The agar plate was transferred into the BaQFA setup and incubated at 37°C under anaerobic conditions. Automated image capturing (every 30 min) was performed.
These images were analyzed using the BaColonyzer software to extract each colony’s growth over time for detailed growth curves (21). Fitness was then derived from the parameters of a Gompertz growth model fitted to these curves as previously described (20). Taking into account the variability inherent in experimental runs, each of the runs of 96 culture spots per plate of the same strain comparisons was considered as a separate study with a potentially random difference in fitness estimate. Therefore, meta-analysis with a random effects model was used for the computation and plotting of relative fitness estimates and their confidence intervals (22), allowing for a comprehensive assessment of strain-specific fitness differences with the reference strain (PAVI-2017310081) in pair-wise comparisons.
Genomic analyses
DNA isolation and genome sequencing
For genomic DNA extraction, the Master Pure Gram-Positive DNA Purification Kit (Lucigen) was used as per the manufacturer’s instructions. Concentration and purity of the isolated DNA were first checked with a NanoDrop ND-1000 (Peqlab, Erlangen, Germany); concentrations were determined using the Qubit dsDNA HS Assay Kit as recommended by the manufacturer (Life Technologies GmbH, Darmstadt, Germany). Illumina shotgun libraries were prepared using the Nextera XT DNA Sample Preparation Kit and subsequently sequenced on a MiSeq system using the v3 reagent kit with 600 cycles (Illumina, San Diego, CA, USA) as recommended by the manufacturer. Quality filtering was done with version 0.36 of Trimmomatic (23). Assembly was performed with version 3.13.0 of the SPAdes genome assembler software (24). Version 2.2.1 of Qualimap was used to validate the assembly and determine the sequence coverage (25). All genome sequences were deposited in GenBank, and the accession numbers are listed in Table S2.
Bioinformatics tools and analyses
For phylogenomic analyses, the core genome was identified and aligned with the Parsnp program from the Harvest software package (26). Reliable core-genome SNPs identified by Parsnp were used for reconstruction of whole-genome phylogeny. Phylogenetic trees were visualized using the Interactive Tree Of Life (iTOL; https://itol.embl.de/). ResFinder (27) was used to identify (acquired) genes mediating antimicrobial resistance.
Proteinortho was used to identify intra- and interclade-specific gene content, applying a bidirectional blast approach (28). Orthologous proteins were identified with the following blast settings: coverage > 50% and identity > 50%. The following genomes were used to identify Clade 1- versus Clade 2-specific gene content differences: CI828_clade 1; CI878_clade1; CI882_clade1; PAVI-2017310081_clade1; PAVI-2017310082_clade1; PAVI-2017310120_clade1; HS4_clade2; HS7_clade2; HS9_clade2; PAVI-2017310084_clade2; PAVI-2017310145_clade2; and PAVI-2017310195_clade2. To identify potential differences between skin isolates and PJI isolates within Clade 1 the following genomes were compared: CI828_clade 1_PJI; CI878_clade1_PJI; CI882_clade1_ PJI; PAVI-2017310081_clade1_healthy skin; PAVI-2017310082_clade1_healthy skin; and PAVI-2017310120_clade1_healthy skin. Annotations were done with BV-BRC (29) and the KEGG tool BlastKOALA (30).
RESULTS
Genomic analysis
Eleven PJI isolates and 32 HS isolates were sequenced. The genome size of the isolates ranged from 2,471 to 2,728 kb, thus differing by a maximum of 257 kb. Annotation predicted coding sequences (CDS), ranging from 2,362 to 2,654, thus differing by a maximum of 292 CDS. Next, a core genome comparison was carried out. Two main phylogenetic clades (Clade 1 and Clade 2) within the C. avidum population could be detected (Fig. 1). Interestingly, all PJI isolates belonged to the same clade (Clade 1). This clade also contained isolates from other PJI cases, including isolates T13, T14, and T15 (8, 31) and FMS2275 and FMS4815 (32), as well as isolates associated with other diseases such as strain TM16, isolated from radical prostatectomy specimens (33). In contrast, HS isolates could be found in both clades, Clade 1 and Clade 2. We noticed that the genomes of Clade 2 isolates (n = 6) were on average 129 kb larger than the genomes of Clade 1 isolates (n = 37). Within Clade 1, genomes of skin isolates (n = 26) were on average 42 kb larger than genomes of PJI isolates (n = 11). We searched for consistent gene content differences between skin isolates and PJI isolates within Clade 1 (Table S3A); only very few CDS or CDS fragments were found to differ (Table S3B). Next, we searched for consistent gene content differences between Clade 1 and Clade 2 isolates (Table S4A). Here, 209 and 272 CDS (or CDS fragments) were found to be Clade 1- or Clade 2-specific, respectively (Table S4B and C).
Phylogenetic comparison of PJI and healthy skin isolates of C. avidum based on the core genome. Core genome-based single-nucleotide variant (SNV) analysis and phylogenetic reconstruction were done with Parsnp. The isolates studied here are highlighted in color; PJI isolates in red; isolates from healthy skin in blue. The other genomes were taken from GenBank (NCBI) (status February 2024). Two large clades can be distinguished. Clade 1 (pink) harbors all PJI isolates. Healthy skin isolates are distributed among Clade 1 (pink) and Clade 2 (green).
Biofilm
We compared biofilm formation of 11 isolates recovered from PJI with 32 isolates from HS (Fig. 2A). No statistically significant differences were found between the two groups. Furthermore, in the sub-analysis performed for Clade 1, the differences between the HS group and the PJI group were also not significant (Fig. 2B). According to the methodology used using a static biofilm assay, all isolates were biofilm producers. In both groups, most of the isolates were strong biofilm producers (93.7% skin group; 72.7% PJI group), and the rest of the isolates were moderate biofilm producers. No statistically significant differences in the proportions of strong and moderate biofilm producers in the two different groups were found (P-value 0.27).
Biofilm formation of healthy skin colonizers and PJI isolates of C. avidum using a modified method of Stepanović et al. (14) as a static biofilm assay. (A) Biofilm formation of all isolates (healthy skin colonizers, n = 32; PJI isolates, n = 11) was compared. (B) Biofilm formation of Clade 1 isolates was compared (healthy skin colonizers, n = 26; PJI isolates, n = 11). Statistical significance was assessed using the Mann-Whitney test following evaluation of data normality.
Antibiotic susceptibility
C. avidum isolates from PJIs displayed generally low MIC values for most antibiotics tested (Fig. 3). MIC and MBIC distributions were nearly identical for the majority of agents, with MBICs being slightly higher in some isolates, indicating limited biofilm tolerance. Rifampin showed the most potent activity against both planktonic and biofilm-embedded cells, with MIC, MBC, and MBIC values ≤0.125 mg/L in all isolates. Notably, MBEC values for rifampin remained low, with most isolates eradicated at ≤0.5 mg/L, contrasting sharply with other agents. For penicillin and levofloxacin, MBC and MBEC values were markedly higher than MIC/MBIC, indicating a limited bactericidal effect and poor biofilm eradication. Amoxicillin-clavulanic acid, linezolid, vancomycin, and clindamycin exhibited favorable MIC and MBC profiles; however, MBEC values were >32 mg/L in most cases, suggesting reduced efficacy in eradicating mature biofilms. One isolate (HOL 4) demonstrated clindamycin elevated MIC, MBC, MBIC, and MBEC values, consistent with the presence of the erm(X) resistance gene. Overall, rifampin was the only antibiotic to consistently exhibit low MIC, MBC, MBIC, and MBEC values, highlighting its superior activity against C. avidum in both planktonic and biofilm states.
Antibiotic susceptibility profiles of Cutibacterium avidum isolates (n = 11) from prosthetic joint infections. Minimal inhibitory concentrations (MICs), minimal bactericidal concentrations (MBCs), minimal biofilm inhibitory concentrations (MBICs), and minimal biofilm eradication concentrations (MBECs) were determined for penicillin, amoxicillin-clavulanic acid, clindamycin, linezolid, vancomycin, levofloxacin, and rifampin. Bars represent the number of isolates exhibiting a given concentration (mg/L) for each antibiotic and susceptibility parameter.
BaQFA
Since reproductive fitness of bacteria plays a major role in the evolution of antimicrobial resistance as well as persistence, which has implications in chronic infections, we quantitatively assessed fitness differences between bacterial isolates. To do so, we used the BaQFA method (20), which allows us to define the relative competitive fitness (RCF) between two given isolates. We chose to assess the RCF of all isolates compared to one randomly selected reference isolate from HS. When including all strains (Clade 1 and Clade 2 strains), we detected significantly higher RCF for HS isolates as compared to isolates which caused PJI (P value 0.039) (Fig. 4A). However, when restricting the analysis to Clade 1 isolates only (the clade that contains PJI and HS isolates), this difference no longer reached statistical significance, although a similar trend was observed (P = 0.054) (Fig. 4B). RCF values within the skin-derived group were notably heterogeneous. Upon exclusion of outlier isolates within the Clade 1 subset, the difference in RCF between PJI and skin isolates further diminished and was not statistically significant (P = 0.099). To explore potential clade-specific differences in bacterial fitness, we compared RCF between C. avidum isolates belonging to Clade 1 and Clade 2. No significant difference in RCF was observed between Clade 1 and Clade 2 isolates (median RCF: Clade 1 = X, Clade 2 = Y; P = 0.8903) (Fig. 4C).
Relative competitive fitness (RCF) of C. avidum isolates assessed using the BaQFA method. (A) RCF values were compared between isolates from all clades obtained from healthy skin (n = 32) and PJIs (n = 11). (B) RCF values were compared between Clade 1 isolates only. No statistical significance, but a trend was seen (P = 0.054). (C) RCF values were compared between Clade 1 and Clade 2 isolates. No significant difference was observed. The dashed horizontal line indicates the reference isolate set at RCF = 1. Plotted values are means from two independent biological repeats. Statistical significance was determined by Mann-Whitney test after testing for normal distribution.
DISCUSSION
To our knowledge, this is the most comprehensive comparative study of C. avidum isolates derived from PJIs and from HS, integrating analyses of phylogeny, biofilm formation, antibiotic susceptibility, and bacterial fitness.
Phylogenomic analysis revealed two distinct phylogenetic clades, Clade 1 and Clade 2, which may represent subspecies of C. avidum. Notably, all PJI isolates belonged exclusively to Clade 1, while skin isolates were distributed across both clades. This suggests that only Clade 1 isolates possess the potential to cause implant-associated infections.
Gene content comparison between Clade 1 and Clade 2 isolates revealed 209 and 272 clade-specific genes, respectively. The majority of genes could not be functionally annotated. Clade 1 specific functions were primarily associated with signaling and transport processes (Fig. S1). In contrast, Clade 2 isolates harbored genes enabling carbohydrate metabolism (e.g., sorbitol, galactitol, and myo-inositol) and de novo fatty acid biosynthesis via the type II fatty acid synthesis (FASII) pathway, which is absent in Clade 1. These metabolic differences may reflect niche-specific adaptation, suggesting that Clade 2 strains are better equipped to survive in nutrient-limited environments, whereas Clade 1 strains may be more dependent on host-derived nutrients such as fatty acids.
All C. avidum isolates in our study exhibited strong biofilm-forming capacity, irrespective of clinical origin or clade affiliation. This stands in contrast to previous findings in C. acnes, where most isolates formed only moderate or weak biofilms (34). The strong biofilm phenotype observed in C. avidum may be linked to a species-specific exopolysaccharide-like structure, which is absent in other Cutibacterium species (33) and may facilitate both surface adherence and antibiotic tolerance (31, 33).
MIC values observed in our study were consistent with previous data (8, 31, 35) and comparable to those reported for C. acnes in orthopedic implant-associated infections (36). One isolate (HOL 4) demonstrated high clindamycin MICs, associated with the erm(X) gene, a known resistance determinant in Cutibacterium spp (37). Importantly, no species-specific EUCAST or CLSI clinical breakpoints exist for C. avidum, and we therefore present our MIC data descriptively without categorizing isolates as “susceptible” or “resistant.” Rifampin showed the lowest MIC, MBC, MBIC, and MBEC values, suggesting high in vitro activity against both planktonic and biofilm-associated bacteria. Nevertheless, the clinical efficacy of rifampin remains debated (38), and its use should be carefully balanced against the risk of adverse effects (39) and the lack of clear evidence from well-controlled studies.
Using BaQFA, we found that PJI isolates showed reduced relative competitive fitness compared to commensal skin isolates, when all isolates are taken into consideration (Clade 1 and Clade 2). A sub-analysis of fitness restricted to Clade 1 showed that the difference between PJI and skin isolates was no longer statistically significant (P = 0.053), although the trend persisted. Given the known role of bacterial fitness in persistence and antimicrobial tolerance, this may represent an adaptive phenotype favoring chronic infection. Reduced fitness is consistent with a more dormant, slow-growing state, potentially resembling bacterial persisters (40). Such states are known to reduce susceptibility to antibiotics and immune clearance (41), thus promoting chronic infection. Our findings support the notion that biofilm-related persistence may be driven not only by extracellular matrix protection, but also by shifts in bacterial growth dynamics and metabolic activity (42–44). Whether reduced RCF is a cause or consequence of biofilm formation in C. avidum remains unclear. One hypothesis is that an adaptation favoring invasiveness may involve a trade-off in proliferative capacity. Alternatively, reduced growth could itself promote biofilm formation and persistence under the hostile conditions of the prosthetic joint microenvironment.
A major limitation of this study is the absence of species-specific clinical breakpoints for C. avidum. Our interpretations rely on descriptive MIC values and cannot inform treatment decisions directly. Additionally, the broth microdilution method used for susceptibility testing, though standard, may underestimate MICs for certain agents such as amoxicillin-clavulanic acid, as EUCAST has noted for C. acnes. Furthermore, while our isolate set represents the largest cohort studied to date, the sample size remains relatively small, limiting statistical power and generalizability.
In summary, C. avidum isolates from PJIs and HS show strong in vitro biofilm formation regardless of clinical origin. All PJI isolates belonged to Clade 1, suggesting this clade harbors the potential for pathogenicity. While antibiotic resistance was rare, biofilm-related persistence and reduced bacterial fitness may contribute to treatment challenges. Our findings underscore the need for species-specific clinical breakpoints, better functional understanding of phylogenetic clades, and further investigation into biofilm-driven phenotypes in C. avidum-associated infections.
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zimmerli W, Trampuz A, Ochsner PE. 2004. Prosthetic-joint infections. N Engl J Med 351:1645–1654. doi:10.1056/NEJ Mra 04018115483283 · doi ↗ · pubmed ↗
- 2Tande AJ, Patel R. 2014. Prosthetic joint infection. Clin Microbiol Rev 27:302–345. doi:10.1128/CMR.00111-1324696437 PMC 3993098 · doi ↗ · pubmed ↗
- 3Shah NB, Tande AJ, Patel R, Berbari EF. 2015. Anaerobic prosthetic joint infection. Anaerobe 36:1–8. doi:10.1016/j.anaerobe.2015.08.00326341272 · doi ↗ · pubmed ↗
- 4Fink B, Sevelda F. 2017. Periprosthetic joint infection of shoulder arthroplasties: diagnostic and treatment options. Biomed Res Int 2017:4582756. doi:10.1155/2017/458275629423407 PMC 5750516 · doi ↗ · pubmed ↗
- 5Benito N, Franco M, Ribera A, Soriano A, Rodriguez-Pardo D, Sorlí L, Fresco G, Fernández-Sampedro M, Dolores del Toro M, Guío L, et al.. 2016. Time trends in the aetiology of prosthetic joint infections: a multicentre cohort study. Clin Microbiol Infect 22:732. doi:10.1016/j.cmi.2016.05.00427181408 · doi ↗ · pubmed ↗
- 6Portillo ME, Corvec S, Borens O, Trampuz A. 2013. Propionibacterium acnes: an underestimated pathogen in implant-associated infections. Biomed Res Int 2013:804391. doi:10.1155/2013/80439124308006 PMC 3838805 · doi ↗ · pubmed ↗
- 7Corvec S. 2018. Clinical and biological features of Cutibacterium (formerly Propionibacterium) avidum, an underrecognized microorganism. Clin Microbiol Rev 31:e 00064-17. doi:10.1128/CMR.00064-1729848774 PMC 6056840 · doi ↗ · pubmed ↗
- 8Achermann Y, Liu J, Zbinden R, Zingg PO, Anagnostopoulos A, Barnard E, Sutter R, Li H, Mc Dowell A, Zinkernagel AS. 2018. Propionibacterium avidum: a virulent pathogen causing hip periprosthetic joint infection. Clin Infect Dis 66:54–63. doi:10.1093/cid/cix 66529020266 PMC 5850018 · doi ↗ · pubmed ↗