ArcA modulates multidrug resistance and compound susceptibility in Klebsiella pneumoniae through ArcB-independent regulation of the SMR efflux pump kpnEF
Tongtong Fu, Zheng Fan, Yuchen Chen, Zhoufei Li, Hongbo Liu, Bing Du, Xiaohu Cui, Yanling Feng, Hanqing Zhao, Guanhua Xue, Jinghua Cui, Chao Yan, Lin Gan, Junxia Feng, Ziying Xu, Yang Yang, Zihui Yu, Yuehua Ke, Jing Yuan

TL;DR
This study shows that ArcA in Klebsiella pneumoniae increases antibiotic resistance by regulating the KpnEF efflux pump, independent of ArcB and through the AckA-Pta pathway.
Contribution
The study reveals ArcA's direct regulation of KpnEF efflux pump and its phosphorylation via the AckA-Pta pathway, independent of ArcB.
Findings
Deletion of arcA increases resistance to antibiotics and disinfectants independently of arcB.
ArcA directly regulates kpnEF transcription by binding to its promoter region.
ArcA phosphorylation in ΔarcB is mediated by the AckA-Pta pathway.
Abstract
Klebsiella pneumoniae has become a major clinical and public health threat due to the increasing prevalence of healthcare-associated infections caused by multidrug-resistant strains. In this study, we demonstrated that the deletion of arcA of ArcAB two-component system diminished the susceptibility of K. pneumoniae to antibiotics, osmotic agents, disinfectants, and structural compounds, which was independent of arcB. RNA-seq analysis revealed a marked upregulation of SMR efflux pump genes kpnEF in the ΔarcA strain compared to the wild-type strain, while the ΔarcB strain exhibited no significant changes. Notably, the deletion of kpnEF in both ΔarcA and wild-type strains abolished their differential susceptibility to antibiotics, osmotic agents, disinfectants, and structural compounds. The EMSA experiments showed that ArcA-P regulated the kpnEF transcriptional expression by directly…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
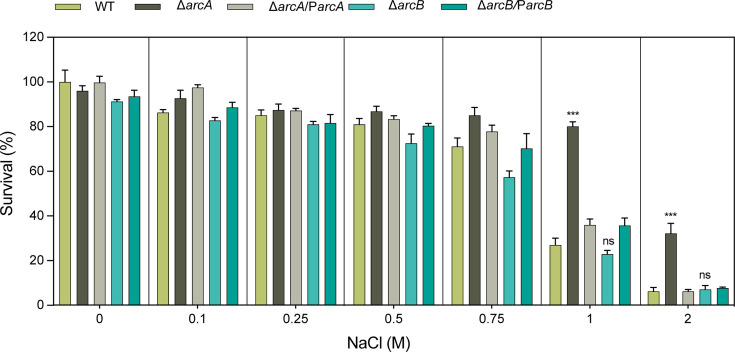 Fig 1
Fig 1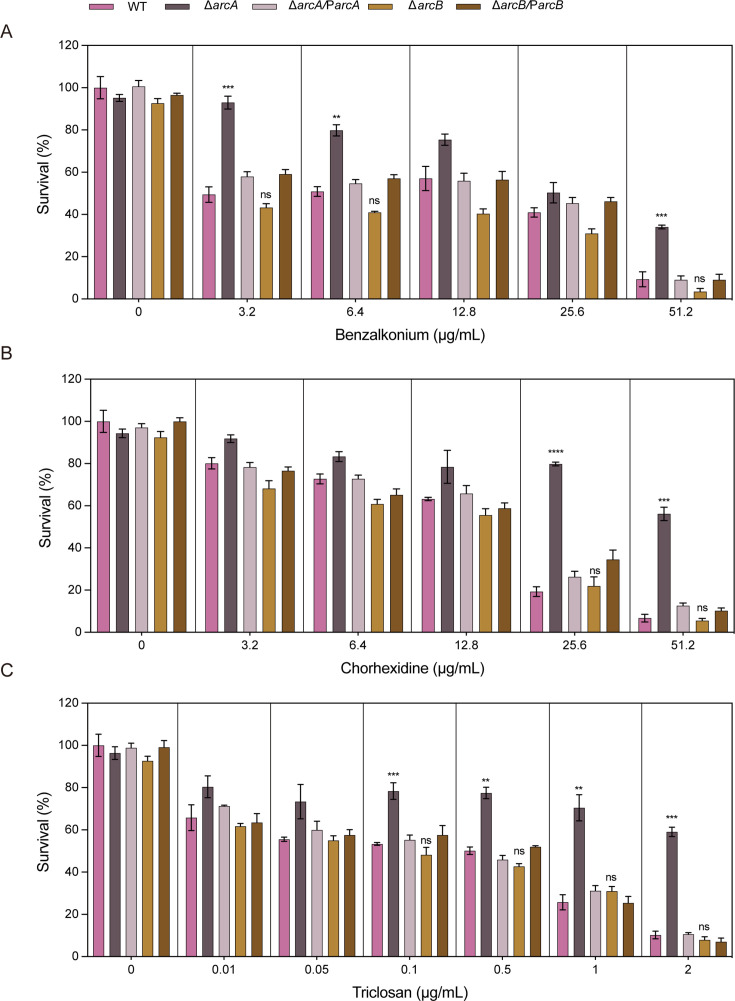 Fig 2
Fig 2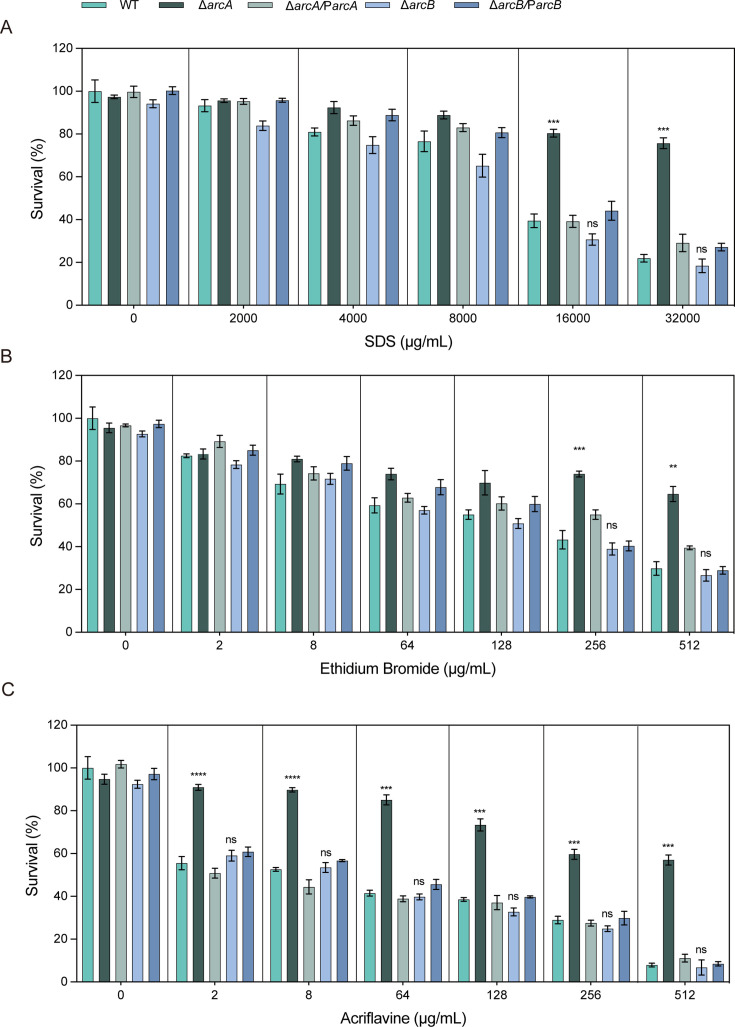 Fig 3
Fig 3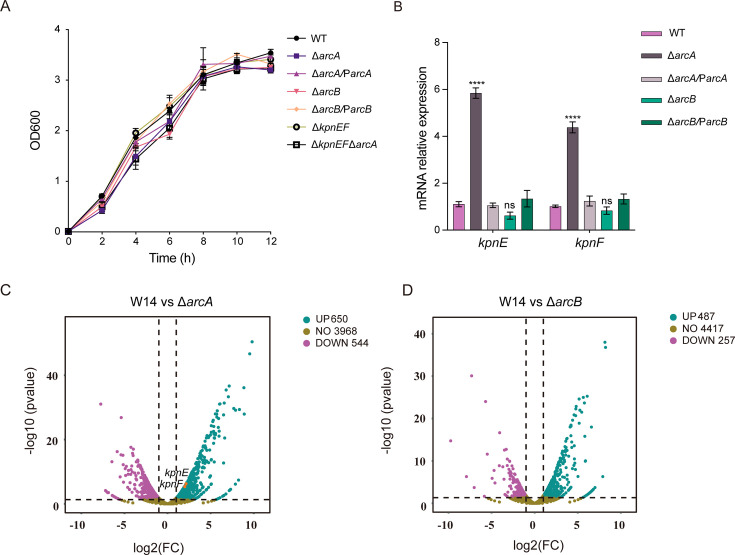 Fig 4
Fig 4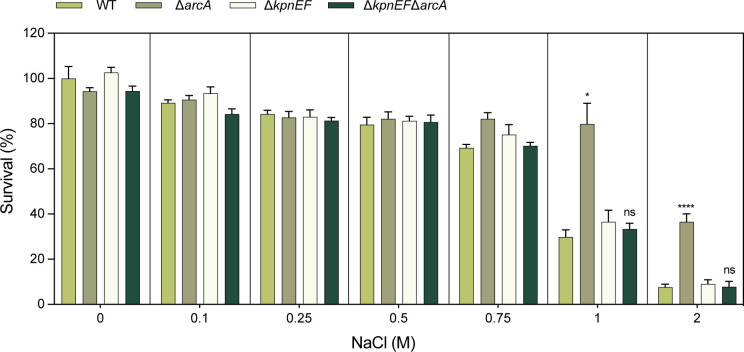 Fig 5
Fig 5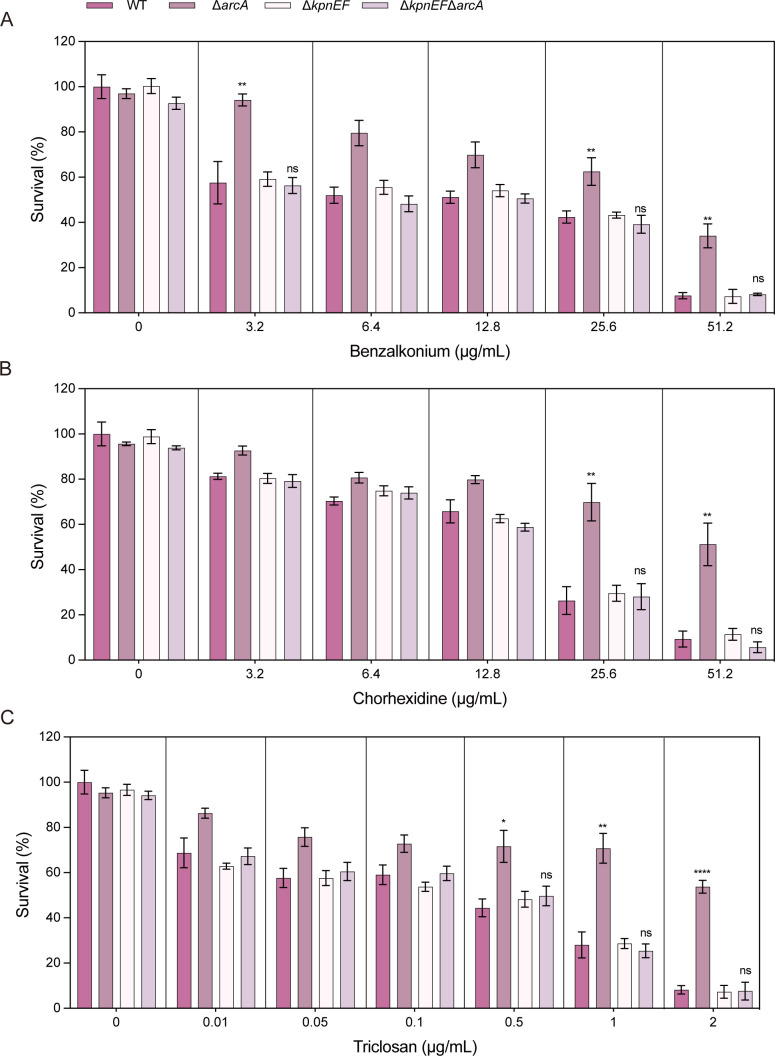 Fig 6
Fig 6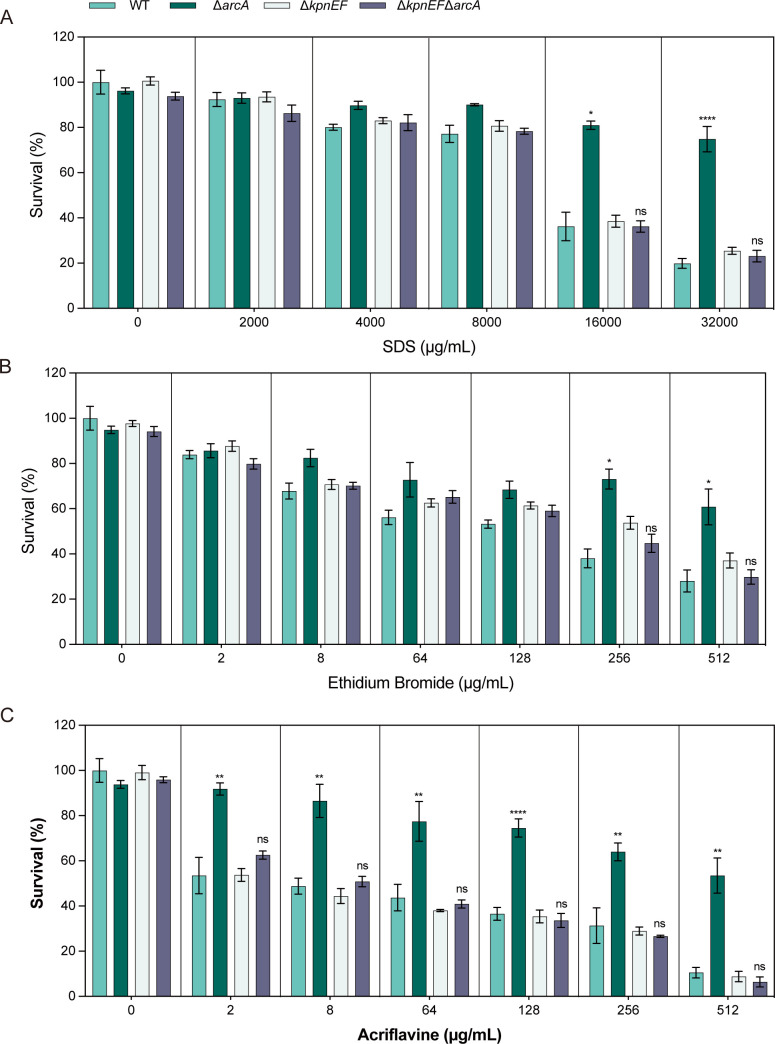 Fig 7
Fig 7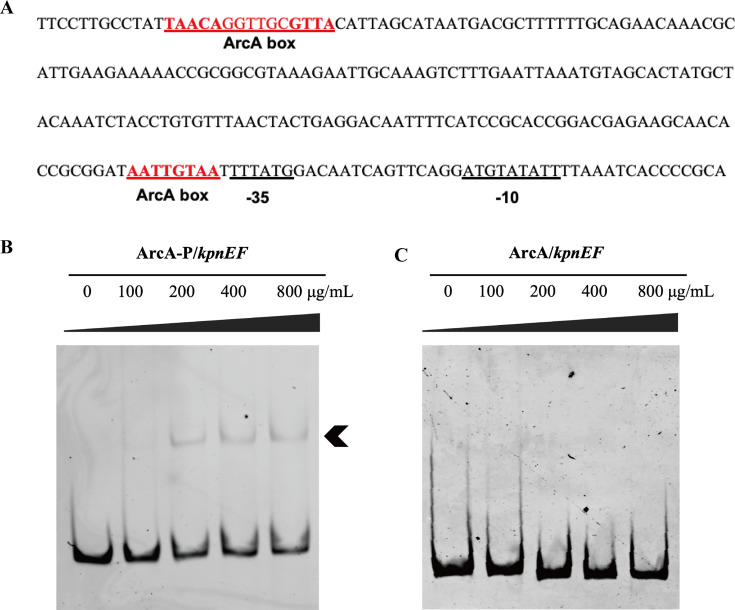 Fig 8
Fig 8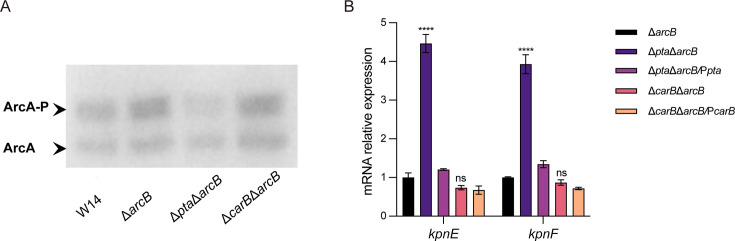 Fig 9
Fig 9| MIC (µg/mL) | |||||
|---|---|---|---|---|---|
| Antibiotics | WT strain | Δ | Δ | Δ | Δ |
| Cefepime | 0.125 | 0.25 | 0.125 | 0.125 | 0.125 |
| Ceftazidime | 0.063 | 0.125 | 0.063 | 0.063 | 0.063 |
| Kanamycin | 0.625 | 2.5 | 0.625 | 0.625 | 0.625 |
| Streptomycin | >62.5 | >62.5 | >62.5 | >62.5 | >62.5 |
| Ciprofloxacin | 0.001 | 0.002 | 0.001 | 0.001 | 0.001 |
| Colistin | 1 | 4 | 1 | 1 | 1 |
| Erythromycin | 1.25 | 1.25 | 1.25 | 1.25 | 1.25 |
| Polymyxin B | 1 | 4 | 1 | 1 | 1 |
| Rifampin | 0.625 | 0.625 | 0.625 | 0.625 | 0.625 |
| Tetracycline | 3.125 | 6.25 | 3.125 | 3.125 | 3.125 |
| MIC (µg/mL) | ||||
|---|---|---|---|---|
| Antibiotics | WT | Δ | Δ | Δ |
| Cefepime | 0.125 | 0.25 | 0.063 | 0.063 |
| Ceftazidime | 0.063 | 0.125 | 0.016 | 0.016 |
| Kanamycin | 0.625 | 2.5 | 0.031 | 0.031 |
| Streptomycin | >62.5 | >62.5 | >62.5 | >62.5 |
| Ciprofloxacin | 0.001 | 0.002 | 0.0005 | 0.0005 |
| Colistin | 1 | 4 | 0.5 | 0.5 |
| Erythromycin | 1.25 | 1.25 | 1.25 | 1.25 |
| Polymyxin B | 1 | 4 | 0.5 | 0.5 |
| Rifampin | 0.625 | 0.625 | 0.625 | 0.625 |
| Tetracycline | 3.125 | 6.25 | 1.563 | 1.563 |
- —Natural Science Foundation of Beijing Municipalityhttp://dx.doi.org/10.13039/501100004826
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Bacterial Genetics and Biotechnology · Antibiotics Pharmacokinetics and Efficacy
INTRODUCTION
Klebsiella pneumoniae is an important opportunistic bacterial pathogen and a leading cause of both nosocomial and community-acquired infections (1). This pathogen exhibits an exceptional ability to acquire a diverse range of antimicrobial resistance (AMR) genes, primarily through the uptake of plasmids and mobile genetic elements (2). This genetic adaptability has led to the emergence of multidrug-resistant (MDR) and extensively drug-resistant (XDR) strains, posing significant challenges to public health. Among these, the rise of carbapenem-resistant K. pneumoniae (CRKP) has particularly complicated clinical treatment, as it limits therapeutic options (3). In such cases, polymyxin B and colistin are often employed as the last line of defense against CRKP infections (4). Bacteria, including K. pneumoniae, have evolved multiple mechanisms to resist antibiotics. These mechanisms commonly include enzymatic modification and inactivation of antibiotics, modification of antibiotic targets, efflux of antibiotics, and the development of bypass mechanisms that can replace or circumvent drug targets (5).
Antibiotic efflux pumps are key to bacterial resistance and are divided into several families: ATP-binding cassette (ABC), major facilitator superfamily (MFS), resistance/nodulation/cell division (RND), multidrug and toxic compound extrusion (MATE), proteobacterial antimicrobial compound efflux (PACE), and small multidrug resistance (SMR). These pumps use proton motive force (PMF) or ATP to expel antibiotics and toxins from cells (6–9). The SMR family is unique for its small size (100–150 amino acids) and four transmembrane helices (TM1–TM4) (10, 11). The SMR protein family is categorized into three distinct subclasses: small multidrug proteins (SMP), suppressor of groEL mutations, and paired small multidrug resistance (PSMR) proteins. EmrE in E. coli and P. aeruginosa, SsmE in S. marcescens, and AbeS in A. baumannii belong to SMP subclass (12–15). The experimentally characterized pairs MdtI/MdtJ, YkkC/YkkD, and EbrA/EbrB are classified within the PSMR subclass (16, 17). The SMR efflux pump KpnEF, a member of the PSMR subclass, functions as a homolog of EbrAB and plays a role in both physiological processes and multidrug resistance within the K. pneumoniae (18). SMR pumps are valuable for studying bacterial resistance due to their simple structure and specific function. They also offer potential targets for new antibiotics and resistance-fighting strategies.
The anoxic redox control (or aerobic respiration control) two-component system, “ArcAB system,” consists of regulator ArcA and the sensor kinase ArcB. The “ArcAB system” is involved in the metabolic transition from aerobic respiration to fermentation and anaerobic respiration when bacteria enter the micro-oxygen and anaerobic environment (19). When sensing oxygen consumption, sensor kinase ArcB auto-phosphorylated and trans-phosphorylated regulator ArcA via the ArcBHis292 - ArcBAsp576 - ArcBHis717- ArcAAsp54 process (20–22). ArcA remains capable of phosphorylation through other pathways in the absence of ArcB. In Shewanella oneidensis MR-1, ArcS likely functions as the cognate sensor kinase of the Arc system, a role previously attributed to ArcB in Escherichia coli (23). The AckA-Pta pathway could modulate the expression of ArcA-P target genes in the absence of ArcB in E. coli (24). In vitro, the ArcA protein could be phosphorylated by carbamoyl phosphate (25, 26). Phosphorylated ArcA could promote fermentation pathway and inhibit pathways associated with aerobic respiration (27–29). ArcA has been characterized and could regulate a multitude of processes, including response to growth limitation, response to reactive oxygen species and even pathogenesis (19). As a broad regulatory factor, the regulatory functions of ArcA remain to be further explored.
Our previous study found that high-alcohol-producing K. pneumoniae (W14), obtained from a patient with auto-brewery syndrome, is a primary causative agent of NAFLD (30). To investigate the mechanism of high ethanol production in W14 and screen for potential therapeutic targets, we constructed knockout strains of the global regulatory factors Fnr, IhfAB, ArcAB, and Crp separately. Subsequently, the ethanol production capacity and antibiotic resistance in these mutant strains were measured. We found that ArcA played a crucial role in the sensitivity of K. pneumoniae to antibiotics, osmotic agents, disinfectants, and structural compounds, which was independent of ArcB. The expression of the SMR efflux pump KpnEF was significantly upregulated in the ΔarcA, so further deletion of kpnEF restored the sensitivity of the ΔarcA. Additional experiments demonstrated that ArcA directly binds to the promoter region of kpnEF, regulating its expression and thereby influencing the bacterial sensitivity to multiple compounds. These findings enhanced our understanding of the regulatory mechanisms underlying drug resistance and enriched the regulatory network of the ArcAB system in K. pneumoniae.
MATERIALS AND METHODS
Bacterial strains, plasmids, and primers
The K30 (clinical Klebsiella pneumoniae) cKp W14 was utilized as the wild-type strain (WT) in this study. All strains employed for both culture and phenotypic analysis were incubated at a temperature of 37°C in a shaking incubator with a speed of 180–200 rpm. And the culture media used was Luria-Bertani (LB) broth (5 g/L yeast extract, 10 g/L sodium chloride, and 10 g/L tryptone). The addition of 50 µg/mL kanamycin was used for the construction of gene knockout strains and complemented strains. The related strains and plasmids were listed in Table S1. The primers in this study were listed in Table S2.
Mutant and complementation strains construction
To obtain the ΔarcA, ΔarcB, ΔkpnEF, and ΔkpnEFΔarcA mutant strains and related complementation strains, methods by Link et al. and Pan et al. were used (31, 32). Briefly, the flanking regions of arcA, arcB, and kpnEF cluster were ligated into the plasmid pKO3-Km (digested by Not I) and the resulting plasmids were transformed into the wild-type strains W14 or the ΔarcA strains by electroporation. At 43°C and 37°C, respectively, plasmid was first inserted into the bacterial chromosome and then shed and screened for the correct mutant strain by 5% sucrose plate. For the construction of the arcA and arcB complementation strain, these genes coding region and their ribosomal binding site were cloned into the plasmid pGEM-T Easy (digested by Sal I and Sph I). Then these resulting plasmids were transformed into their related mutant strains respectively by electroporation.
Bacterial growth curves
Growth curves of W14, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains were drawn by subculturing in LB broth overnight. Briefly, the overnight cultures of wild type (W14), deletion strains and complementary strains were diluted 1:100 into 20 mL of fresh LB broth and then grown by shaking at 200 rpm at 37°C. OD600 measurements were detected per hour to represent the cell density. Three independent cultures were used for each assay.
RNA extraction, reverse transcription, RNA-seq, and quantitative real-time PCR
The bacterial suspension was subjected to centrifugation at a speed of 11,340 × g, and then bacterial precipitate was resuspended in 100 µL of 1× RNase-free TAE and subsequent extraction of total RNA was performed utilizing RNAprep Pure Cell/Bacteria Kit (Tiangen Biotech, Beijing, China). A quantity of RNA ranging from 0.5 to 1 µg was employed for the process of reverse transcription, wherein cDNA was synthesized utilizing PrimeScript Reverse Transcriptase (TaKaRa, Dalian, China). The qRT-PCR experiment was conducted by employing a 20 µL mixture comprising the SYBR Premix Ex TaqTM II (TaKaRa), cDNA templates, and specific forward and reverse primers. The qRT-PCR reactions were performed on the QuantStudio 5 Real-Time PCR System (Applied Biosystems, USA). The Ct values of the target genes were normalized to the reference gene rpoB, and the fold-change in gene expression was calculated using the classic 2^(−ΔΔCt) method. RNA sequencing and analysis services were performed by NoVo gene (Beijing, China).
Minimal inhibit concentration assay
The MICs were determined following the guidelines outlined in the Clinical and Laboratory Standards Institute (CLSI) M07 11th edition and M100 32nd edition. E. coli ATCC 25922 was employed as the quality control strain. Overnight cultures were diluted 1:100 into fresh medium and incubated at 37°C with shaking at 220 rpm until the OD600 reached 0.8–1.0. The bacterial strains used for MIC determination showed no significant differences in colony counts. The bacterial suspension was then appropriately diluted and dispensed into 96-well plates. Antibiotics, including cefepime, ceftazidime, kanamycin, ciprofloxacin, streptomycin, erythromycin, rifampicin, colistin E, polymyxin B, and tetracycline, were serially diluted using the twofold broth microdilution method and added to the 96-well plates. For the MICs testing of the above-mentioned antibiotics, cation-adjusted Mueller-Hinton broth (CAMHB) medium was utilized, with each well receiving an inoculum of 5 × 10⁵ CFU. The plates were incubated at 37°C for 20 h, after which the MIC was determined as the lowest antibiotic concentration that completely inhibited visible bacterial growth.
Structurally related compounds, hospital-based disinfectants, osmotic compounds challenge assays
Various stress assays were conducted as previously described (33). Briefly, mutant, complemented, and WT strains were cultured to mid-exponential phase, and the cultures were then spread onto LB agar plates containing different concentrations of the following compounds: osmotic stress agent NaCl (0.075 M, 0.15 M, 0.25 M, 0.5 M, 0.75 M, 1 M, and 2 M); disinfectants include benzalkonium chloride, chlorhexidine (3.2 µg/mL, 6.4 µg/mL, 12.8 µg/mL, 25.6 µg/mL, 51.2 µg/mL) and triclosan (0.01 µg/mL, 0.05 µg/mL, 0.1 µg/mL, 0.5 µg/mL, 1 µg/mL, 2 µg/mL); structure-related compounds include SDS (1,024 µg/mL, 2,048 µg/mL, 4,096 µg/mL, 8,192 µg/mL, 16,384 µg/mL), ethidium bromide, and acriflavine (2 µg/mL, 8 µg/mL, 64 µg/mL, 128 µg/mL, 256 µg/mL, 512 µg/mL). The results were expressed as the ratio of the number of colony-forming units (CFU) obtained from LB plates containing different concentrations of NaCl, disinfectants, and structure-related compounds to the number of CFU obtained from control plates (LB agar alone). Each experiment was repeated at least three times.
Protein expression and purification
To construct the ArcA expression plasmid, the arcA gene fragment was amplified from the W14 genome and ligated into the Nco I and Hind III digested pET28a vector. Transformants were selected on kanamycin plates, and the correct pET28A-His6-ArcA expression plasmid was verified by PCR and sequencing. The confirmed pET28a-His6-ArcA plasmid was then transformed into E. coli BL21(DE3). The transformants were cultured in LB medium supplemented with 50 µg/mL kanamycin at 37°C until the OD600 reached 0.6, followed by induction with 0.5 mM IPTG at 37°C for 3 hs. The cells were harvested and lysed by sonication in lysis buffer (25 mM HEPES, 5 mM β-mercaptoethanol, 500 mM NaCl, and 10% glycerol, pH 7.8). The His-tagged fusion protein was purified using nickel-nitrilotriacetic acid (Ni-NTA) agarose resin (Qiagen, Beijing, China) and eluted with elution buffer (25 mM HEPES, 5 mM β-mercaptoethanol, 500 mM NaCl, 10% glycerol, and 300 mM imidazole, pH 7.8). The target protein was stored at −80°C until further use. The concentration of the purified protein was determined by SDS-PAGE and quantified using the bicinchoninic acid (BCA) assay with a standard protein as the reference.
Electrophoretic mobility shift assay
The EMSA was performed following a previously described method with slight modifications (34). We amplified the DNA fragment of the kpnEF promoter region using W14 as the template. The kpnEF promoter (50 ng) DNA fragment was pre-incubated with increasing concentrations of purified ArcA protein in a binding buffer (20 µL) containing 10 mM Tris–HCl (pH 8.0), 1 mM EDTA, 1 mM DTT, 1.5 mg/mL poly IC, 50 mM KCl, 50 µg/mL BSA, and 10% glycerol, supplemented with potassium lithium acetyl phosphate (20 nM), at ambient temperature for 30 min. An 8% polyacrylamide gel was pre-electrophoresed in 1 × Tris-borate-EDTA buffer (0.044 M Tris, 0.044 M boric acid, and 0.001 M EDTA, pH 8.0) for 1 h to remove impurities. After loading the samples, electrophoresis was conducted on ice for 1 h and 40 min. Following electrophoresis, the gel was stained with 0.5 µg/mL ethidium bromide. Imaging was performed using a gel documentation system (Bio-Rad, Hercules, CA, USA).
Phos-assay western blotting
The phos assay reagent (Vazyme, Nanjing, China) was added to the 8.5% SDS-PAGE separating gel for phosphorylation level detection. Cells were lysed in SDS-loading buffer (2% β-mercaptoethanol) and boiled for 10 min. Protein concentrations were determined by BCA assay, and 20 µg of protein was separated by Phos-assay SDS-PAGE. After transfer to PVDF membranes, blots were blocked with 5% BSA/TBST and probed with anti-His (1:2,000, Sigma, USA) at 4°C overnight, followed by HRP-conjugated secondary antibody anti-Mousse IgG (1:2,000, Promega, USA). Signals were detected by Immobilon Western HRP.
Co-immunoprecipitation/MS
The ΔarcB mutant strain complemented with His-tagged ArcA was lysed in NP-40 buffer (50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1% NP-40, protease inhibitors). Lysates were split into two aliquots: one incubated with anti-His beads (Sigma, USA), the other with mouse IgG1 isotype control (Thermo, USA) for 4 h at 4°C. Both groups used Protein G magnetic beads and identical wash conditions. After washing five times with NP-40 buffer, bound proteins were eluted with 2 × Laemmli buffer. IgG controls were processed in parallel. Samples were trypsin-digested (Promega, USA) and analyzed by LC-MS/MS (Q Exactive HF-X). Data were searched against the UniProt Escherichia coli database (2023 release) using PEAKS Studio with a 1% FDR threshold. Specific interactors required ≥2 unique peptides and Fold Change (His/IgG) ≥2.
Statistical analysis
Three biological replicates were performed for each experiment. Statistical analysis was conducted using GraphPad Prism (version 5.0, USA), and the data were presented as mean ± standard deviation (SD). Statistical significance was determined by one-way ANOVA, with P < 0.05, P < 0.01, and P < 0.001, P < 0.0001 indicating significant differences. Following one-way analysis of variance (ANOVA) which revealed significant differences, we performed all pairwise comparisons using Tukey’s Honest Significant Difference (HSD) test. Growth curve statistical analyses were performed using SPSS software Statistics 27.0.1 with repeated-measures ANOVA.
RESULTS
ArcA influenced the susceptibility of K. pneumoniae to multiple antibiotics, and this effect was independent of arcB
To investigate whether the two-component system ArcA-ArcB affected the antibiotic resistance of K. pneumoniae, the minimum inhibitory concentrations (MICs) of WT, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains were determined against cefepime, ceftazidime, kanamycin, streptomycin, ciprofloxacin, colistin E, erythromycin, polymyxin B, rifampicin, and tetracycline. The MICs of the ΔarcA strains for the above antibiotics were two-, four-, four-, one-, two-, four-, one-, four-, one-, and twofold higher than those of the wild-type strains, respectively (Table 1). The antibiotic susceptibility of the ΔarcA strains was restored in the ΔarcA/ParcA strains. In contrast, the MICs of the ΔarcB strains were consistent with those of the WT strains (Table 1).
ArcA decreased the susceptibility of K. pneumoniae to osmotic stress compound, hospital disinfectants, and structurally related compounds by an arcB-independent manner
In this study, the susceptibilities of the WT, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains to different concentrations of osmotic stress compound, disinfectants, and structurally related compounds were evaluated. Under NaCl conditions (physiological concentration, 150 mM) (33, 35), the viability of the ΔarcA was significantly higher than that of the wild type. Specifically, in 1 M NaCl, the growth capacity of the ΔarcA was approximately 2.65-fold greater than that of the wild-type, and in 2 M NaCl, it was approximately 4.38-fold greater. This difference was independent of the inoculum size (Fig. 1).
*The susceptibility of the WT, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains to osmotic stress agent. Sensitivities of WT, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains to different concentrations (0.075, 0.15, 0.25, 0.5, 0.75, 1.0, and 2.0 M) of NaCl. The survival ability of the ΔarcA in 1 M NaCl was 2.65-fold greater, and in 2 M NaCl, it was 4.38-fold greater compared to the WT strain. The resistance percentages to different concentrations of NaCl were calculated by comparing the numbers of surviving cells to those in LB medium alone. Asterisks indicate statistically significant differences compared to the WT strain. Each experiment was performed in triplicate. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001, analyzed with one-way ANOVA.
When cells were exposed to varying concentrations of benzalkonium chloride, it was observed that at 51.2 µg/mL, the total CFU count of the ΔarcA was 3.64-fold higher than that of the wild type; at 6.4 µg/mL, the total CFU count of the ΔarcA was 1.57-fold higher than that of the wild type, and at 3.2 µg/mL, the total CFU count of the ΔarcA was 1.72-fold higher than that of the wild type (Fig. 2A). When cells were exposed to varying concentrations of chlorhexidine, it was observed that at 51.2 µg/mL, the total CFU count of the ΔarcA was 8.35-fold higher than that of the wild type, and at 25.6 µg/mL, the total CFU count of the ΔarcA was 3.80-fold higher than that of the wild-type strain (Fig. 2B). Similarly, when cells were exposed to 0.1, 0.5, 1, and 2 µg/mL triclosan, the total CFU count of the ΔarcA was 1.47-, 1.50-, 2.58-, and 5.26-fold higher than that of the wild type (Fig. 2C).
*The susceptibility of the WT, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains to hospital-based disinfectants. (A) Sensitivities of the WT, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains to different concentrations (3.2 µg/mL, 6.4 µg/mL, 12.8 µg/mL, 25.6 µg/mL, 51.2 µg/mL) of benzalkonium chloride. The survival ability of the ΔarcA in benzalkonium chloride at 51.2 µg/mL was 3.64-fold greater, at 6.4 µg/mL, it was 1.57-fold greater, and at 3.2 µg/mL, it was 1.72-fold greater compared to the WT strain. (B) Sensitivities of WT, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains to different concentrations (3.2 µg/mL, 6.4 µg/mL, 12.8 µg/mL, 25.6 µg/mL, 51.2 µg/mL) of chlorhexidine. The survival ability of the ΔarcA in chlorhexidine at 51.2 µg/mL was 8.35-fold greater, and at 25.6 µg/mL was 3.80-fold greater compared to the WT strain. (C) Sensitivities of WT, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains to different concentrations (0.01 µg/mL, 0.05 µg/mL, 0.1 µg/mL, 0.5 µg/mL, 1 µg/mL, 2 µg/mL) of triclosan. The survival ability of the ΔarcA in triclosan at 0.1 µg/mL was 1.47-fold, at 0.5 µg/mL was 1.50-fold, at 1 µg/mL was 2.58-fold, and at 2 µg/mL was 5.26-fold greater compared to the WT strain. The resistance percentages to different concentrations of benzalkonium chloride, chlorhexidine, and triclosan were calculated by comparing the numbers of surviving cells to those in LB medium alone. Asterisks indicate statistically significant differences compared to the WT strain. Each experiment was performed in triplicate. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001, analyzed with one-way ANOVA.
When cells were exposed to varying concentrations of sodium dodecyl sulfate (SDS), it was observed that at 16,000 µg/mL, the CFU count of the ΔarcA was 1.88-fold higher than that of the wild type, and at 3,200 µg/mL, the CFU count of the ΔarcA was 3.45-fold higher than that of the wild type (Fig. 3A). Additionally, the susceptibility of the ΔarcA to ethidium bromide (EtBr) at 256 µg/mL and at 512 µg/mL (Fig. 3B) was decreased by 1.56-fold and 1.54-fold, respectively. Similarly, when cells were exposed to 2-, 8-, 64-, 128-, 256-, and 512 µg/mL acriflavine, the total CFU count of the ΔarcA was 1.50-, 1.60-, 1.84-, 1.90-, 1.92-, and 7.22-fold higher than that of the wild type (Fig. 3C). These results indicate that the deletion of arcA broadly affects the susceptibility of K. pneumoniae to various substances.
*The susceptibility of the WT, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains to structurally related compounds. (A) Sensitivities of WT, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains to different concentrations (1,024 µg/mL, 2,048 µg/mL, 4,096 µg/mL, 8,192 µg/mL, 16,384 µg/mL) of SDS. The survival ability of the ΔarcA in SDS at 16,000 µg/mL was 1.88-fold and at 32,000 µg/mL was 3.45-fold higher compared to the WT strain. (B) Sensitivities of WT, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains to different concentrations (2 µg/mL, 8 µg/mL, 64 µg/mL, 128 µg/mL, 256 µg/mL, 512 µg/mL) of ethidium bromide. The survival ability of the ΔarcA in ethidium bromide at 256 µg/mL was 1.56-fold, and at 512 µg/mL was 1.54-fold compared to the WT strain. (C) Sensitivities of WT, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains to different concentrations (2 µg/mL, 8 µg/mL, 64 µg/mL, 128 µg/mL, 256 µg/mL, 512 µg/mL) of acriflavine. The survival ability of the ΔarcA in acriflavine at 2-, 8-, 64-, 128-, 256-, and 512 µg/mL was 1.50-, 1.60-, 1.84-, 1.90-, 1.92-, and 7.22-fold higher compared to the WT strain. The resistance percentages to different concentrations of SDS, ethidium bromide, and acriflavine were calculated by comparing the numbers of surviving cells to those in LB medium alone. Asterisks indicate statistically significant differences compared to the WT strain. Each experiment was performed in triplicate. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001, analyzed with one-way ANOVA.
Transcriptome analysis revealed that the upregulation of the gene of SMR kpnEF in the ΔarcA strain might be responsible for the reduced susceptibility
The growth curves of the WT, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains were determined to investigate whether the altered susceptibility of the ΔarcA to various compounds was growth-related. As shown in Fig. 4A, significant growth impairment in the ΔarcA and ΔarcB vs the WT (P < 0.05), no significant difference between the ΔarcA and ΔarcB strains (P > 0.05). To explore the regulatory mechanisms mediated by ArcA on bacterial responses to antibiotics, osmotic agents, disinfectants, and structurally related compounds, the global gene expression profiles of the WT, ΔarcA, and ΔarcB strains were analyzed using RNA-seq. Differentially expressed genes were identified based on an adjusted with P < 0.05 and |log2 (fold change) | >1. The results revealed that, among 5,406 genes, 1,194 were differentially expressed between the WT and ΔarcA strains and 744 between the WT and ΔarcB strains. Volcano plot analysis of the RNA-seq data showed that 544 genes were downregulated and 650 genes were upregulated in the ΔarcA (Fig. 4C), while 257 genes were downregulated and 487 genes were upregulated in the ΔarcB (Fig. 4D). Notably, the expression of the small multidrug efflux pump gene kpnE and kpnF was upregulated 4- and 3.71-fold in the ΔarcA, but no significant change was observed in the ΔarcB. The top 20 most upregulated and top 20 most downregulated genes in the ΔarcA compared to the wild-type and in the ΔarcB compared to the wild-type were showed in Tables S3 and S4. The qRT-PCR experiments further confirmed that the expression of kpnE and kpnF in the ΔarcA was 5.28- and 4.29-fold higher than that in the wild type, while the expression level in the ΔarcA/ParcA was restored to that of the wild type (Fig. 4B). In contrast, no significant changes in kpnEF expression were observed in the ΔarcB or the arcB/ΔarcB compared to the wild type (Fig. 4B). These findings suggested that ArcA regulates the expression of kpnEF independently of ArcB.
*The regulation of ArcAB in K. pneumoniae. The growth curves of WT, ΔarcA, ΔarcA/ParcA, ΔarcB, ΔarcB/ParcB, ΔkpnEF, and ΔkpnEF/ΔarcA strains (A). (B)The transcript levels of kpnE and kpnF in the WT, ΔarcA, ΔarcB, ΔarcA/ParcA, and ΔarcB/ParcB strains were detected by qRT-PCR. Volcano plot depicting gene transcripts to be differentially expressed between the WT and ΔarcA strains (C), WT and ΔarcB strains (D). Red and green dots represent significantly upregulated and downregulated proteins, respectively. ArcA negatively regulates the transcription of kpnEF. Error bars represent the standard errors of the means of three biological replicates. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001, analyzed with one-way ANOVA.
The deletion of the kpnEF gene restored the antibiotic susceptibility of the ΔarcA strain
To investigate whether ArcA affected the antibiotic resistance of K. pneumoniae through the small multidrug efflux pump KpnEF, the mutant strains ΔkpnEF and ΔkpnEFΔarcA were constructed by deleting kpnEF in the WT and ΔarcA background. Prior to MIC determination, growth curves were established for the WT, ΔarcA, ΔkpnEF, and ΔkpnEFΔarcA strains. No significant differences were observed between the WT and ΔkpnEF strains, nor between the ΔarcA and ΔkpnEFΔarcA strains (Fig. 4A). MIC assays revealed that the differential antibiotic susceptibility between the ΔarcA and WT strains was dependent on KpnEF, as deleting kpnEF in both backgrounds abolished this difference (Table 2). Additionally, ΔkpnEF alone showed significantly reduced resistance compared to WT, confirming the role of KpnEF as a key efflux pump contributing to intrinsic antibiotic resistance in this K. pneumoniae.
The deletion of kpnEF also restored the sensitivity of ΔarcA strain to osmotic stress, disinfectants, and structurally related compounds
To explore whether ArcA affected the susceptibility of K. pneumoniae to osmotic agents, disinfectants, and structurally related compounds through the small multidrug efflux pump KpnEF, the susceptibilities of the WT, ΔarcA, ΔkpnEF, and ΔkpnEFΔarcA strains to varying concentrations of these compounds were determined. Under conditions of different concentrations of NaCl, benzalkonium chloride, chlorhexidine, triclosan, SDS, deoxycholate, EtBr, and acriflavine, the sensitivity of the ΔkpnEF to these compounds was not significantly different from that of the ΔkpnEFΔarcA, while was significantly lower than that of wild-type strains. The results show that KpnEF influences the susceptibility of W14 to these compounds, and ArcA can influence the susceptibility of W14 to these compounds by mediating the expression of KpnEF (Fig. 5 to 7).
*The osmotic stress agent susceptibility in the WT, ΔarcA, ΔkpnEF, and ΔkpnEFΔarcAstrains. Sensitivities of the WT, ΔarcA, ΔkpnEF, and ΔkpnEFΔarcA strains to different concentrations (0.075, 0.15, 0.25, 0.5, 0.75, 1.0, and 2.0 M) of NaCl. The MICs of the tested antibiotics showed no significant difference between ΔkpnEFΔarcA and ΔkpnEF strains. ΔkpnEF exhibited significantly reduced antibiotic susceptibility compared to the WT strain. The tolerant percentages to different concentrations of NaCl were calculated by comparing the numbers of surviving cells to those in LB medium alone. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001, analyzed with one-way ANOVA.
*The hospital-based disinfectants susceptibility in the WT, ΔarcA, ΔkpnEF, and ΔkpnEFΔarcA strains. (A) Sensitivities of WT, ΔarcA, ΔkpnEF, and ΔkpnEFΔarcA strains to different concentrations (3.2 µg/mL, 6.4 µg/mL, 12.8 µg/mL, 25.6 µg/mL, 51.2 µg/mL) of benzalkonium chloride. (B) Sensitivities of the WT, ΔarcA, ΔkpnEF, and ΔkpnEFΔarcA strains to different concentrations (3.2 µg/mL, 6.4 µg/mL, 12.8 µg/mL, 25.6 µg/mL, 51.2 µg/mL) of chlorhexidine. (C) Sensitivities of the WT, ΔarcA, ΔkpnEF, and ΔkpnEFΔarcA strains to different concentrations (0.01 µg/mL, 0.05 µg/mL, 0.1 µg/mL, 0.5 µg/mL, 1 µg/mL, 2 µg/mL) of triclosan. The sensitivity of ΔkpnEF mutant to these hospital-based disinfectants was not significantly different from that of ΔkpnEFΔarcA mutant, while was significantly lower than that of WT strains. The tolerant percentages to different concentrations of benzalkonium chloride, chlorhexidine, and triclosan were calculated by comparing the numbers of surviving cells to those in LB medium alone. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001 analyzed with one-way ANOVA.
*The structurally related compounds susceptibility in the WT, ΔarcA, ΔkpnEF, and ΔkpnEFΔarcA strains. (A) Sensitivities of WT, ΔarcA, ΔkpnEF, and ΔkpnEFΔarcA strains to different concentrations (1,024 µg/mL, 2,048 µg/mL, 4,096 µg/mL, 8,192 µg/mL, 16,384 µg/mL) of SDS. (B) Sensitivities of the WT, ΔarcA, ΔkpnEF, and ΔkpnEFΔarcA strains to different concentrations (2 µg/mL, 8 µg/mL, 64 µg/mL, 128 µg/mL, 256 µg/mL, 512 µg/mL) of ethidium bromide. (C) Sensitivities of the WT, ΔarcA, ΔkpnEF, and ΔkpnEFΔarcA strains to different concentrations (2 µg/mL, 8 µg/mL, 64 µg/mL, 128 µg/mL, 256 µg/mL, 512 µg/mL) of acriflavine. The sensitivity of ΔkpnEF mutant to these structurally related compounds was not significantly different from that of ΔkpnEFΔarcA mutant, while was significantly lower than that of WT strains. The tolerant percentages to different concentrations of SDS, ethidium bromide, and acriflavine were calculated by comparing the numbers of surviving cells to those in LB medium alone. *P < 0.05, *P < 0.01, *** P < 0.001, **** P < 0.0001, analyzed with one-way ANOVA.
ArcA ~P could specifically bind to the promoter region of kpnEF and repressed their expression, independently of ArcB
Through upregulating SMR efflux pump KpnEF, the ΔarcA enhanced multidrug resistance and decreased susceptibility to osmotic agents, disinfectants, and structurally related compounds in K. pneumoniae. To determine whether ArcA regulated KpnEF by direct binding and whether phosphorylation of ArcA promotes its binding to the KpnEF promoter region, we conducted in vitro phosphorylation and the EMSA experiments. Based on previous reports, ArcA is known to bind to a conserved sequence (5′-TGTTA-3′) in E. coli (36). Subsequently, we used published consensus sequences and the Softberry bioinformatics platform to search for conserved ArcA binding sites in the kpnEF promoter region (37). Then the similar sequences (TAACA-GGTTGC-GTTA) and (AATTGTAA) located in the kpnEF promoter region were identified (Fig. 8A). The ArcA protein was expressed in E. coli BL21 using the pET-28a vector. EMSA was performed to detect the binding of ArcA and phosphorylated ArcA (ArcA-P) to a 574 bp DNA fragment of the kpnEF promoter region. As the amount of ArcA-P increased, the amount of free DNA gradually decreased (Fig. 8B). In contrast, the DNA mobility was unaffected when non-phosphorylated ArcA protein was used (Fig. 8C). These results indicate that ArcA requires phosphorylation to bind to the promoter region, and this process is independent of ArcB-mediated phosphorylation of ArcA, suggesting the existence of other proteins capable of phosphorylating ArcA.
ArcA regulated the expression of the kpnEF operon by directly binding to their promoter regions. The promoter sequence of the kpnEF and the locations of the ArcA boxes were marked (A). The interactions between ArcA and the DNA fragments of their promoter regions were detected with EMSA. ArcA-P was incubated with the kpnEF promoter region (B), and ArcA was incubated with the kpnEF promoter region (C) at 37°C for 30 min. The mixtures were electrophoresed on 8% native polyacrylamide gels and the bands were imaged under UV light after EB staining.
ArcA phosphorylation was mediated by the AckA-Pta pathway when ArcB was absent in K. pneumoniae
Phosphorylation level assays demonstrated that the phosphorylation level of ArcA in the ΔarcB showed no significant difference compared to the wild-type, indicating the existence of alternative pathways mediating ArcA phosphorylation (Fig. 9A). The Co-IP experiment was performed to investigate whether other kinases could phosphorylate ArcA through direct binding. As shown in Table S5, the results were identified by adjusted −10 lgP ≥30 and unique ≥2. The kinase capable of directly binding and phosphorylating ArcA was not identified under these experimental conditions. Subsequently, the knockout strains of pta and carB genes were constructed in the ΔarcB background, respectively. Expression analysis of the kpnEF operon in these mutants demonstrated that deletion of pta significantly increased kpnEF expression (Fig. 9B). Phosphorylation assays further confirmed that the deletion of pta markedly decreased ArcA phosphorylation levels (Fig. 9A). These results demonstrate that in K. pneumoniae, ArcA could be phosphorylated via the AckA-Pta pathway under the absence of ArcB condition, enabling its regulatory function.
*ArcA phosphorylation was mediated by the AckA-Pta pathway in the ΔarcB strain. (A) The phosphorylation levels of ArcA in the W14, ΔarcB, ΔptaΔarcB, and ΔcarBΔarcB were performed by phos-assay. (B) The transcript levels of kpnE and kpnF in the ΔarcB, ΔptaΔarcB, ΔcarBΔarcB, ΔptaΔarcB/pta, and ΔcarBΔarcB/carB strains were detected by qRT-PCR. Error bars represent the standard errors of the means of three biological replicates. ***P < 0.0001 analyzed with one-way ANOVA.
DISCUSSION
The ArcAB system, also known as the Arc system, is a member of the bacterial two-component regulatory family, consisting of the sensor kinase ArcB and the response regulator ArcA. The ArcAB system exhibits broad regulatory capabilities, participating not only in central metabolism but also in a range of cellular processes, including bacterial conjugation, biofilm formation, stress response, acid tolerance, and even bioluminescence (38–42). ArcAB is predicted to orchestrate the sophisticated integration of these cellular processes with cognate metabolic pathways, thereby furnishing the energy and metabolic precursors essential for the execution of these functions. The regulatory role of ArcAB is particularly evident during infection, likely because pathogens must balance metabolic demands with survival in the host environment (36, 43, 44). In this study, we found that ArcA influenced sensitivity of K. pneumoniae to antibiotics, osmotic agents, disinfectants, and structural compounds by regulating the expression of the KpnEF multidrug efflux pump. This discovery enriched the regulatory network of the ArcAB system.
There is growing evidence that ArcA and ArcB do not always function physiologically as the “ArcAB system.” In Serratia marcescens, ArcB could regulate bacterial motility by activating the synthesis of flagella without regulator ArcA (45). In Salmonella enterica, ArcA was required in bacterial response to oxidative stress independent of its cognate sensor ArcB (46). And in this study, we found that the ArcA could regulate kpnEF independent of ArcB in K. pneumoniae. Thus, ArcA may act alone or in conjunction with other sensor protein to form another two-component system, while ArcB may also function with other regulator. Compared with other two-component systems, genes arcA and arcB are not found together in the same operon (47–49), which may explain the functional differences shown by ArcA and ArcB, since the expression and regulation of these two genes are relatively independent. Hence, the understanding of the function of ArcA and ArcB is far from enough.
Studies on the ArcA-DNA complex have revealed that ArcA typically bound to DNA fragments following dimerization or oligomerization (36, 50). The formation of higher-order ArcA oligomers is dependent on its phosphorylation status. Phosphorylated ArcA has been demonstrated to bind promoter regions at the −35 and −10 elements, as well as positions up to 500 bp upstream. The canonical ArcA-binding motif spans approximately 15 bp, with the most frequently observed sequence GTTAATTAAATGTTA being conserved across E. coli, Salmonella, and Shewanella oneidensis. Within this sequence are two direct repeats (GTTA) (51–54). In the current study, P-ArcA was found to bind to the promoter sequence of kpnEF, while non-phosphorylated ArcA showed no binding activity. This suggested that ArcA interacted with this fragment exclusively in its phosphorylated form. However, it remained unclear whether this binding occurred in a dimerized state. The phosphorylation level of ArcA primarily depended on the AckA-Pta pathway. Notably, the AckA-Pta pathway regulated acetyl phosphate (AcP) levels, which, in turn, influenced global protein acetylation, capsular polysaccharide (CPS) production, serum resistance, and type 3 fimbriae expression ultimately modulating virulence and playing a critical role in K. pneumoniae pathogenesis. In future studies, it will be essential to examine whether AcP levels are altered in the ΔarcA or ΔarcB and whether such changes affect the pathogenicity of K. pneumoniae.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wyres KL, Lam MMC, Holt KE. 2020. Population genomics of Klebsiella pneumoniae. Nat Rev Microbiol 18:344–359. doi:10.1038/s 41579-019-0315-132055025 · doi ↗ · pubmed ↗
- 2Wyres KL, Holt KE. 2018. Klebsiella pneumoniae as a key trafficker of drug resistance genes from environmental to clinically important bacteria. Curr Opin Microbiol 45:131–139. doi:10.1016/j.mib.2018.04.00429723841 · doi ↗ · pubmed ↗
- 3Yang X, Dong N, Chan EW-C, Zhang R, Chen S. 2021. Carbapenem resistance-encoding and virulence-encoding conjugative plasmids in Klebsiella pneumoniae. Trends Microbiol 29:65–83. doi:10.1016/j.tim.2020.04.01232448764 · doi ↗ · pubmed ↗
- 4Aye SM, Galani I, Yu H, Wang J, Chen K, Wickremasinghe H, Karaiskos I, Bergen PJ, Zhao J, Velkov T, Giamarellou H, Lin YW, Tsuji BT, Li J. 2020. Polymyxin triple combinations against polymyxin-resistant, multidrug-resistant, KPC-producing Klebsiella pneumoniae. Antimicrob Agents Chemother 64:e 00246-20. doi:10.1128/AAC.00246-2032393492 PMC 7526826 · doi ↗ · pubmed ↗
- 5Foster TJ. 2017. Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiol Rev 41:430–449. doi:10.1093/femsre/fux 00728419231 · doi ↗ · pubmed ↗
- 6Piddock LJV. 2006. Multidrug-resistance efflux pumps - not just for resistance. Nat Rev Microbiol 4:629–636. doi:10.1038/nrmicro 146416845433 · doi ↗ · pubmed ↗
- 7Piddock LJV. 2006. Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin Microbiol Rev 19:382–402. doi:10.1128/CMR.19.2.382-402.200616614254 PMC 1471989 · doi ↗ · pubmed ↗
- 8Hassan KA, Maher C, Elbourne LD, Henderson PJ, Paulsen IT. 2021. Increasing the PACE of characterising novel transporters by functional genomics. Curr Opin Microbiol 64:1–8. doi:10.1016/j.mib.2021.08.00534492595 · doi ↗ · pubmed ↗