First report of Anaplasma marginale in the European bison Bison bonasus
Anna W. Myczka, Stanisław Kaczor, Elwira Plis-Kuprianowicz, Joanna Werszko, Aleksandra Kawińska, Arkadiusz Juszczyk, Anna Bajer, Zdzisław Laskowski

TL;DR
This study reports the first detection of Anaplasma marginale in European bison in Poland, raising concerns about its impact on this protected species.
Contribution
The first report of Anaplasma marginale in European bison (Bison bonasus) in Europe.
Findings
Anaplasma spp. was detected in 40% of European bison tissue samples.
Anaplasma marginale DNA was identified in eight samples, marking its first report in Poland and in European bison.
Anaplasma phagocytophilum was also detected in 10 samples.
Abstract
European bison, Bison bonasus, is a strictly protected species of large mammal, with 25% of the world’s population living in Poland. The most numerous populations of European bison live in the Białowieża Primeval Forest, northeastern Poland, and in the Bieszczady Mountains, southeastern Poland. The purpose of this study was to investigate the occurrence of Anaplasma spp. in B. bonasus from Poland. Tissue samples were collected from 45 European bisons between 2021 and 2024 in the Białowieża and Bieszczady areas. Two genetic markers, 16S ribosomal DNA (rDNA) and msp4, were used for the detection, genotyping, and phylogenetic analysis of bacteria from the genus Anaplasma. The prevalence of Anaplasma spp. was 40% (18/45) in the examined samples. Anaplasma phagocytophilum was detected in 10 samples, and eight samples were found to be positive for the presence of Anaplasma marginale DNA.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
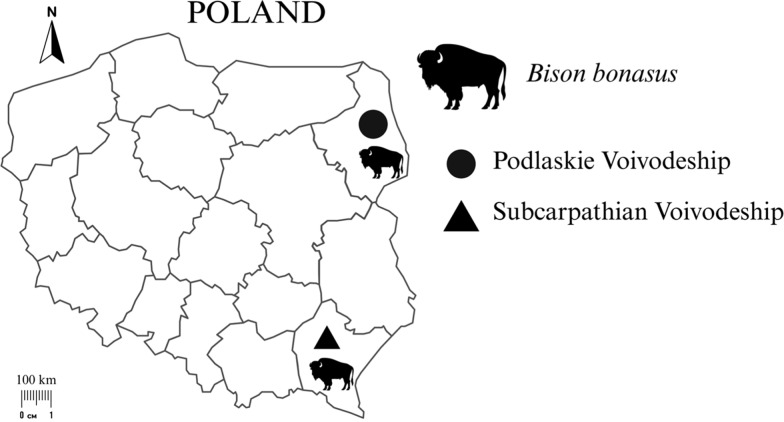 Figure 1
Figure 1 Figure 2
Figure 2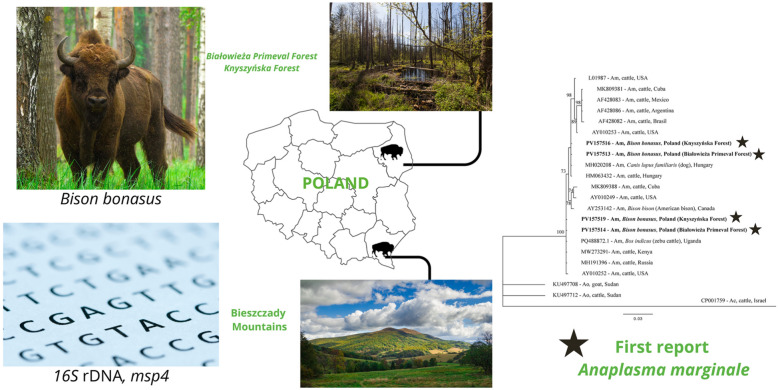 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Aquaculture disease management and microbiota · Bartonella species infections research
Background
The current global population of European bison (Bison bonasus Linnaeus, 1758) consists of 11,180 individuals. A significant share (25%) of this population lives in Poland, mainly in two areas: the Bieszczady Mountains and the Białowieża Primeval Forest [1]. This strictly protected species was nearly extinct at the beginning of the twentieth century, but since 1919, the population has been restored with great success, including in Poland [2]. Health monitoring is currently the key aspect of species conservation and research [3, 4]. Because of a limited gene pool of European bisons, the species can be prone/susceptible to various infectious diseases [5]. The recent rapid spread of Thelazia spp. infection in the two largest free-living populations may be an example of such susceptibility [6].
Anaplasma marginale (family Anaplasmaceae, order Rickettsiales) is an obligate intracellular bacterium that invades erythrocytes, similar to its close relatives, Anaplasma ovis and Anaplasma centrale [7–9]. It is the most pathogenic Anaplasma species for cattle, but it does not infect humans [10, 11]. It is the causative agent of bovine anaplasmosis, a livestock disease with a great economic impact on dairy and cattle production [10]. The most frequently reported clinical signs are fever, anorexia, anemia, rumen atony, fetus loss, changes in urine color (dark yellow to brown), weakness, a reduction in milk production, and death in acute infection [7, 11–14].
In the literature, the endemic occurrence of A. marginale is strongly associated with tropical and subtropical climates [7]. However, pathogens are widely reported worldwide in North America (the USA [15], Canada [16], Mexico [17]), South America (Argentina [18], Brazil [19]), Africa (Egypt [20], Uganda [21], Zimbabwe [22]) and Asia (Malaysia [23], India [24], and Iran [25]). In Europe, probably due to climate change, the northern border of the geographical range of vectors which can transmit A. marginale, is moving north annually [26]. The first reports concerning European bovine anaplasmosis and the occurrence of A. marginale were from Mediterranean areas, such as France [27], Italy [28], Portugal [29], and Spain [30]. Cases have since been described in Central European countries such as Austria [31], Switzerland [32], and Hungary [8]. To the best of our knowledge, in Europe, there are no reports on A. marginale occurrence further north than the three-country (Hungary, Austria, Switzerland) line.
Anaplasma marginale can be transmitted between hosts by three routes: by vectors (ticks and tabanids), mechanical transfer of live infected erythrocytes (by needle pokes or surgical equipment), and through the placenta (vertical transmission) [10, 33, 34]. However, the wide distribution and great success of the spread of A. marginale in various habitats is due primarily to vector transmission, mainly by hard ticks from the family Ixodidae [34]. Over 20 species of ticks from three genera, Dermacentor, Hyalomma, and Rhipicephalus, can transmit this pathogen [9, 10, 34]. Dermacentor reticulatus ticks occur widely, whereas Hyalomma spp. occur sporadically in Poland [35, 36]; however, their role in A. marginale transmission in Poland is unknown. Moreover, within Dermacentor spp., D. reticulatus is not listed as a vector of A. marginale [34]. A few species from the family Tabanidae are also reported as vectors of A. marginale. This intracellular pathogen was detected in Tabanus bovinus, Tabanus tergestinus, and Poeciloderas lindneri in Europe and South America [37, 38]. A few of the species also occur in Poland, but A. marginale was not detected in them [39].
The aim of this study was to investigate the occurrence of Anaplasma spp. in B. bonasus from Poland. We determined the prevalence of infection, and genotyping and phylogenetic analyses were conducted to characterize the obtained Anaplasma spp. species and strains.
Methods
Material collection and study area
Spleen samples were collected from 45 free-living European bison individuals. Tissue samples were collected between 2021 and 2024 at three study sites in Poland: the Białowieża Primeval Forest and the Knyszyńska Forest in Podlaskie Voivodeship in Poland (n = 19) and the Bieszczady Mountains in Subcarpathian Voivodeship in Poland (n = 26) (Fig. 1). Samples from the Podlaskie Voivodeship were taken from bisons found dead for unknown reasons and road-killed individuals (n = 11) or from individuals culled during selective shooting (n = 8) conducted by employees of the Białowieża National Park and the Knyszyńska Forest District (collection permit decisions: PN.51.10.2021 AK, DZP-WG.6401.121.2024.EB). Tissue samples from the Subcarpathian Voivodeship were collected from 26 individuals culled at the discretion of the General Directorate for Environmental Protection (collection permit decisions: DZP-WG.6401.2.2021.EB) and with the approval of the Regional Directorate for Environmental Protection in Rzeszów (collection permit decisions: WPN6401.1.194.2021.RN.2).Fig. 1. Geographical distribution of the collected Bison bonasus tissue samples from Podlaskie Voivodeship (NE) and Subcarpathian Voivodeship (SE)
Anaplasma spp. molecular detection
Genetic material was isolated from the spleen samples using a commercial kit for DNA isolation (DNA Mini Kit Syngen, Wrocław, Poland; and Genomic Mini AX Tissue Spin, A&A Biotechnology, Gdańsk, Poland) according to the manufacturers’ protocols.
The DNA of Anaplasma spp. was detected in samples by nested polymerase chain reaction (PCR) amplification and sequencing of a 400-base-pair (bp) fragment of 16S ribosomal DNA (rDNA) according to Szewczyk et al. [40]. In samples positive for Anaplasma spp. DNA, partial msp4 (major surface protein 4) gene fragments were amplified to characterize the obtained Anaplasma spp. An 850-bp fragment of the msp4 gene was amplified according to de la Fuente et al. [15]. The nested PCR (for amplification of 16S rDNA) and PCR (for amplification of msp4) were performed in a volume of 20 µl (2 µl of 10× PCR Dream Taq Green buffer, 0.1 µl of 5U Dream Taq, 0.2 µl of 10 mM dNTPs [Thermo Fisher Scientific, Waltham, MA, USA], 1 µl of 10 mM of each primer, and 2 µl of template DNA and filled with nuclease-free water up to 20 µl). The DNA of Anaplasma phagocytophilum from wild boar (GenBank accession number: MT510541) was used as a positive control [41], whereas nuclease-free water was used as a negative control.
The PCR products were visualized on a 1.5% agarose gel (EURx, Gdańsk, Poland) stained with Midori Green stain (Nippon Genetics, Düren, Germany) in the presence of a size-marked DNA marker of 100–1000 bp (A&A Biotechnology, Gdańsk, Poland). Visualization was performed by GelDoc MP Lab software (Imagine, Bio-Rad, Hercules, CA, USA). The obtained PCR products were sequenced using the Sanger method by Eurofins Genomics (Ebersberg, Germany). The sequences were aligned in CodonCode Aligner v. 11.0.1 software (CodonCode Corporation, Centerville, MA, USA). The obtained sequences were compared with the GenBank database (Basic Local Alignment Search Tool [BLAST], National Center for Biotechnology Information [NCBI], USA) and then submitted to GenBank. Phylogenetic trees of the A. marginale msp4 partial gene (786 bp) were constructed in Bayesian inference (BI), as implemented in MrBayes version 3.2.0 software [42]. The SYM + I model was selected for the msp4 sequences as the best-fitting nucleotide substitution model by jModelTest version 2.1.10 software [43, 44]. The analysis was run for 2,000,000 generations, with 1,000,000 generations discarded as burn-in. The phylogenetic trees were visualized in TreeView software (S&N Genealogy Supplies, UK).
Results
Eighteen of the 45 examined samples were positive for Anaplasma spp., resulting in a prevalence of 40%. Of these 18 positive samples, 13 samples were from the Podlaskie Voivodeship and five from the Podkarpackie Voivodeship. Some findings regarding the detection of Anaplasma spp. by molecular methods have been published previously [45]. Among the 18 obtained 16S rDNA sequences, two species were identified: A. marginale (n = 8) on the basis of 99.76–100% identity to A. marginale from cattle from different countries (i.e., Uganda [KU686793], Kazakhstan [PQ133434], Republic of South Africa [AF414873]), and A. phagocytophilum (n = 10) on the basis of 99.76–100% identity to A. phagocytophilum from Poland [ON025953], Russia [HM366579], and Estonia [MW922756]). Data concerning A. phagocytophilum from this study will be published elsewhere. This study is the first confirmed report of A. marginale in Poland and the first report of A. marginale in B. bonasus in Europe. The general prevalence of A. marginale was 17.8% (8/45); however, this pathogen was detected only in Podlaskie Voivodeship, and the prevalence in this area was 42.1% (8/19). Anaplasma phagocytophilum was detected in both study areas, and its prevalence was 22.2% (10/45).
Among the eight A. marginale-positive samples, two sequence variants of 16S rDNA and msp4 gene fragments were detected (Table 1). There was a two-nucleotide difference in the 795-bp sequence of the msp4 gene between the two groups, namely, genotype 1 (five sequences) and genotype 2 (three sequences), resulting in 99.75% similarity between the genotypes. Five sequences of msp4, which formed A. marginale “genotype 1” (this study), showed 100% identity with the sequence of A. marginale from cattle, Hungary (HM063432). This Hungarian genotype was described in 2012 by Hornok et al. [8] and was associated with fatal and anaemic cases of anaplasmosis in cows in Hungary. Three msp4 sequences that constituted “genotype 2” showed 100% identity for several A. marginale strains from cattle from different countries: Kenya (MW273291), Uganda (PQ488872), Russia (MH191396), and Israel (AY786993). There was only one single-nucleotide polymorphism (SNP) in the short sequences of 16S rDNA between the two groups of sequences, resulting in 99.75% similarity between these groups.Table 1. Results of the molecular analysis of Anaplasma marginale strains, isolated from European bison (Bison bonasus)European bison sample numberGenBank accession numbersStudy siteGenotypemsp4**16S rDNAZ22PV157513^a^PV151354Białowieża Primeval forest1Z38PV157515PV151355Białowieża Primeval ForestZ41PV157516^a^PV151356Knyszyńska ForestZ43PV157518PV151357Knyszyńska ForestZ45PV223869PV151358Knyszyńska ForestZ36PV157514^a^PV151359Białowieża Primeval Forest2Z42PV157517PV151360Knyszyńska ForestZ44PV157519^a^PV151361Knyszyńska Forest^a^Nucleotide sequences selected as representatives of genotype groups in sequence and phylogenetic analysis
For phylogenetic analysis, sequences of msp4 were analysed, with 17 sequences of A. marginale and two root sequences of A. ovis and A. centrale from GenBank (Table 2, Fig. 2). Phylogenetic analysis of the msp4 sequences revealed that the sequences could be divided into two clades. These two clades represented genotypes 1 and 2 and were closely related to each other. Genotypes 1 and 2 were grouped with A. marginale from Europe (Hungary, Russia, Moscow Region) and Africa (Uganda, Kenya), respectively (Fig. 2).Table 2. Selected literature sequences (GenBank) for msp4 partial gene phylogenetic analysisPathogen speciesStrain name, sample originAccession numberHostReferenceAnaplasma marginaleOklahoma, USAAY010252CattleDe la Fuente et al. [46]St. Maries, USAAY010249CattleDe la Fuente et al. [46]Florida, USALO1987CattleWickwire et al. [47]; Allred et al. [48]Okeechobee, USAAY010253CattleDe la Fuente et al. [47]Minas Gerais, BrazilAF428082CattleDe la Fuente et al. [46]Salta, ArgentinaAF428086CattleDe la Fuente et al. [46]Texcoco, MexicoAF428083CattleGarcia Ortiz et al. [49]; Garcia Ortiz et al. [50]; Rodriguez et al. [51]Oriente CuBov140, CubaMK809381Bos taurus (cattle)Díaz-Sánchez et al. [52]CuBov132, CubaMK809388Bos taurus (cattle)Díaz-Sánchez et al. [52]Moscow Region, RussiaMH191396CattleNo dataKilifi County, KenyaMW273291CattleNo dataUgandaPQ488872Bos indicus (zebu cattle)Etiang et al. [53]CanadaAY253142Bison bison (American bison)De la Fuente et al. [54]HungaryHM063432CattleHornok et al. [8]HungaryMH020208DogNo dataA. ovisSudanKU497708GoatNo dataSudanKU497712CattleNo dataA. centraleIsraelCP001759CattleHerndon et al. [55]Fig. 2. Phylogenetic tree of the msp4 partial gene (786 bp) of Anaplasma marginale haplotypes constructed in Bayesian inference (BI) analysis with MrBayes version 3.2 [39]. The SYM + I model was chosen as the best-fitting nucleotide substitution model by JModelTest version 2.1.10 software [40, 41]. The analysis was run for 2,000,000 generations, with 1,000,000 generations discarded as burn-in. Nodal support is indicated as Bayesian posterior probabilities. Sequences from A. centrale (CP001759) and A. ovis (KU497708, KU497712) were used as outgroups. GenBank accession numbers, hosts, and countries of origin are shown. The sequences from this study are in bold. Am = A. marginale, Ao = A. ovis, Ac = A. centrale
Discussion
To the best of our knowledge, this is the first confirmed report of A. marginale in Poland, the first report of A. marginale in B. bonasus, and the first report of this pathogen occurring further north than Hungary, in Europe [8].
There is one report about the potential occurrence of A. marginale in Poland. In 2011, Anaplasma spp. were tested in Warmian–Masurian Voivodeship cattle by serological methods (enzyme-linked immunosorbent assay [ELISA]), and the seroprevalence of A. marginale was 1.7% (23/1326) [56]. However, the commercial test used in this study, the Anaplasma Antibody Test Kit, cELISA (VMRD, Inc., Pullman, WA, USA), which is still available on the market, enables the detection of several Anaplasma species in addition to A. marginale, as stated in the manufacturer’s description. Different species of Anaplasma can infect cattle, including A. phagocytophilum, A. centrale, and A. bovis, in addition to A. marginale [57]. This previous report was based solely on ELISA, and the findings were not confirmed by molecular or microscopic (smear) methods. In addition, at that time in Europe, A. marginale was not reported in cattle, even in Hungary, until 2009 [8].
Bison bonasus populations in Poland are affected by numerous parasites and bacteria, including Ashworthius sidemi, Haemonchus contortus, Ostertagia kolchida, Thelazia gulosa, Toxoplasma gondii, Chlamydia spp., and A. phagocytophilum [4, 6, 45, 58]. Our study has added A. marginale to the list of European bison pathogens. This pathogen can constitute a serious threat to the health of cattle and European bisons, which are representative of the family Bovidae. Bovine anaplasmosis is responsible for annual losses in livestock of up to 9 million euros worldwide [59]. Additionally, many fatal cases of bovine anaplasmosis are reported every year [8, 60, 61], which highlights the real threat of A. marginale to the B. bonasus population. Notably, the detection of A. marginale among the Polish population of the European bison is most likely an example of autochthonous infection, because the Polish population of B. bonasus was restored as the first one in Europe [1]. In fact, in many European countries where this species was reintroduced, Polish individuals were used to establish a population [62]. Therefore, the research we present sheds new light on aspects of A. marginale epidemiology, considering the transport of individual bison for reintroduction purposes, theoretically from an area where A. marginale was not previously present or endemic. This may indicate a potential epidemiological threat to wild and farm animals in new areas where reintroduced B. bonasus individuals may serve as reservoirs of A. marginale. In contrast to the cattle trade, which facilitates the transport of A. marginale-infected livestock from endemic to non-endemic regions [7], this is not the case in the Polish population of B. bonasus.
So far, there are no data about the prevalence of A. marginale in B. bonasus. To date, only A. phagocytophilum, related to A. marginale, has been detected in Polish and European populations of European bison [46, 63–67], with quite marked prevalence rates ranging from 18% to 67%. In the present study, the overall prevalence of Anaplasma spp. was 40%, 22.2%, and 17.8% for A. phagocytophilum and A. marginale, respectively, which is in line with previous reports [46, 63–67]. Interestingly, the prevalence of A. marginale in American bison (Bison bison) in US and Canadian populations is much higher, ranging from 88% to 100% [54].
An interesting aspect of our study is the occurrence of A. marginale in only one population of examined European bison, Podlaskie Voivodeship (Białowieża Primeval Forest and Knyszyńska Forest), and the lack of detection of this pathogen in samples from the Bieszczady Mountains. The presence or absence of A. marginale can be associated with the occurrence and/or availability of its vectors in the examined areas. A similar situation was described in the Argentinian population of cattle [68], where Rhipicephalus spp. are associated with the transmission of A. marginale in the environment. Rhipicephalus microplus occurs in northern Argentina, the area with optimal temperature and humidity for reproduction of this tick species, but it was not found in southern Argentina. A similar pattern was observed for A. marginale occurrence: infection was detected in the northern area but not in the southern area [68].
The vector of A. marginale in Poland is unknown. Three genera of ticks and flies are referred to as possible vectors: Dermacentor, Hyalomma, and Rhipicephalus [9, 10, 34], and T. bovinus, T. tergestinus, and P. lindneri [37, 38]. Dermacentor reticulatus (ornate dog tick), Hyalomma spp., and T. bovinus occur in Poland [35, 36, 38]. It is highly unlikely that Hyalomma spp. ticks are responsible for the transmission of A. marginale in Poland, as the first report of the occurrence of these ticks was from 2024 (unpublished data, [36]), and according to our results, this bacterium was detected in samples collected in 2021. From the family Tabanidae, only A. phagocytophilum was reported in examined specimens in Poland [39]. It seems more likely that A. marginale can be transmitted by ornate dog ticks. The eastern metapopulation of this hard tick is reported in the northwestern part of Poland (Podlaskie Voivodeship), the same region where A. marginale is prevalent in the examined animals. The opposite situation occurs in the southwestern region, where D. reticulatus is not present (Subcarpathian Voivodeship), and no positive samples for A. marginale were detected there [35]. In the Central European B. bonasus population, two species of ticks feed on bisons, Ixodes ricinus and D. reticulatus [67], but the first hard tick is not identified as an A. marginale vector, and in the second, A. marginale has not yet been detected, in either Poland or Europe. Additionally, because A. marginale was detected in two different areas (Białowieża National Park and Knyszyńska Forest District), there is a low probability of transmission of these bacterial pathogens other than by vector transmission. However, the occurrence of A. marginale in Poland has not yet been confirmed in any of the hard tick species, and the vector of transmission remains unknown.
For phylogenetic and phylogeographical analysis of A. marginale isolates from European bisons, fragments of the msp4 gene were used. This genetic marker is highly conserved among A. marginale and is more useful and informative for this type of analysis than the 16S rDNA gene. In addition, the msp4 gene is frequently used in phylogenetic analysis [8, 15]. Our msp4 sequences of genotype 1 clustered with the European strain of A. marginale from cattle and dogs in Hungary. The sequences of genotype 2 clustered with A. marginale sequences from Africa and Russia (Moscow Region). None of the genotypes obtained from this study were strongly associated with A. marginale strains from North and South America, which were isolated from naturally infected hosts. Our results of phylogenetic analysis seem to confirm that tropical and subtropical bacterial parasites, such as A. marginale, are expanding their geographical range northwards from Africa to Europe, likely in response to climate change [26].
Conclusions
Our study provides new data about the occurrence, genetic diversity, and changes in the geographical range of A. marginale. Infection by the pathogenic A. marginale in strictly protected species such as the European bison may have an impact on the health, ecology, and conservation of this endangered species. The tick vectors and routes of A. marginale transmission in Poland remain unknown. The influence of climate change on the distribution of this pathogen and its vector must be investigated in the near future to understand the short- and long-term consequences for humans, wild animals, and livestock.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Raczyński J, Bołbot M, Księga Rodowodowa Żubrów [In Polish]. 2023. www.bpn.gov.pl/ksiega-rodowodowa-zubrow.
- 2Krasiński ZA. Żubr Puszcz Imperator [In Polish]. Wydawnictwo BPN: Białowieża, Poland. 2005. https://bpn.gov.pl/zubr-puszcz-imperator.
- 3Romanek W, Bajer A. Narodowe Kleszczobranie—Nauka Obywatelska. [in Polish]. (2025). the promising start of a citizen science project. Parasites Vectors 18:383. 10.1186/s 13071-025-07022-4www.narodowekleszczobranie.pl.10.1186/s 13071-025-07022-4PMC 1246219040993823 · doi ↗ · pubmed ↗