Rapid acquisition of daptomycin resistance in a Corynebacterium striatum osteoarticular infection: case report and discussion on antimicrobial resistance
Ines Rezzoug, Emmanuel Zamparini, Charles Court, Assaf Mizrahi, Laurent Dortet, Cécile Emeraud

TL;DR
A patient with a bone infection caused by Corynebacterium striatum developed resistance to daptomycin after treatment, highlighting the need for caution in using this antibiotic for long-term infections.
Contribution
This study reports a rare case of rapid daptomycin resistance in C. striatum and identifies potential genetic mutations linked to resistance.
Findings
A daptomycin-susceptible C. striatum isolate became resistant after 14 days of monotherapy.
Whole-genome sequencing identified a nonsense mutation in pgsA likely responsible for daptomycin resistance.
Mutations in mltG and ftsI genes may explain increased β-lactam resistance in the resistant strain.
Abstract
Corynebacterium striatum, a commensal of the skin microbiota, can cause invasive osteoarticular infections that are challenging to diagnose and treat due to biofilm formation. Daptomycin, owing to its potent activity against gram-positive bacteria and favorable toxicity profile, is frequently used as empirical therapy in the management of postoperative bone and joint infections. However, cases of acquired resistance remain rare and poorly understood. We describe a case of chronic T11–T12 spondylodiscitis in a paraplegic patient. During the first surgical intervention, a daptomycin-susceptible C. striatum isolate (CORY-1) was recovered. The patient received 14 days of daptomycin monotherapy (10 mg/kg). At the time of the second surgery, a daptomycin-resistant C. striatum strain (CORY-2) was isolated. Whole-genome sequencing revealed only 13 single nucleotide polymorphisms between the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
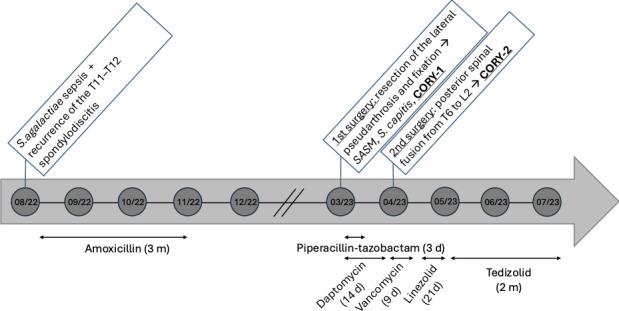 Fig 1
Fig 1| Antibiotic | MIC (mg/L) | EUCAST interpretation | |
|---|---|---|---|
| March 2023 CORY-1 | April 2023 CORY-2 | ||
| Amoxicillin | 6 | 6 | R |
| Meropenem | 0.25 | 0.75 | No data |
| Imipenem | 0.75 | 1.5 | No data |
| Piperacillin-tazobactam | >256 | >256 | No data |
| Vancomycin | 0.25 | 0.5 | S |
| Teicoplanin | 0.12 | 0.12 | No data |
| Daptomycin | 0.12 | >32 | No data |
| Dalbavancin | 0.015 | 0.015 | No data |
| Linezolid | 0.25 | 0.25 | S |
| Tedizolid | 0.12 | 0.12 | S |
- —French National Reference Center for Antimicrobial Resistance
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDiphtheria, Corynebacterium, and Tetanus · Orthopedic Infections and Treatments · Bacterial Identification and Susceptibility Testing
INTRODUCTION
Corynebacterium striatum is a gram-positive bacillus and a commensal organism of the skin flora, but it can also cause invasive infections (1). This species has been implicated in various osteoarticular infections, including osteomyelitis, septic arthritis, and prosthetic joint infections (2–4). True C. striatum infections are difficult to diagnose, as several positive samples are necessary to rule out the possibility of contamination. Furthermore, treatment is often difficult due to the ability of C. striatum to form biofilms that protect the bacteria and contribute to its persistence (5). Vancomycin is often the antibiotic of choice to treat C. striatum infections. However, other antibiotics, such as linezolid, tetracyclines, or daptomycin, have shown low minimum inhibitory concentrations (MICs), suggesting potential effectiveness against this pathogen (2).
Daptomycin is a cyclic lipopeptide antibiotic that is bactericidal and active against gram-positive bacilli. It possesses efficacy comparable to that of vancomycin but with a lower toxicity profile, making it a preferred treatment option (1). Its bactericidal activity relies on a unique mechanism of action involving membrane disruption. In the presence of calcium ions (Ca^2+^), daptomycin binds to the bacterial plasma membrane, composed mainly of phospholipids, leading to the formation of an ion channel. This causes membrane depolarization and potassium efflux, ultimately resulting in cell death.
Daptomycin resistance has been described in Staphylococcus, Enterococcus, and Corynebacterium species (6). In Staphylococcus, resistance to daptomycin is due to the accumulation of negatively charged phospholipids on the cell membrane surface, leading to electrostatic repulsion of the daptomycin molecule. This phenomenon results from an accumulation of mutations in several genes, including mprF, cls2, and pgsA (7, 8). In contrast, in Enterococcus species, daptomycin resistance is primarily driven by mutations in cls and gdpD genes, which alter membrane composition and may lead to cell wall thickening, or by mutations affecting the LiaFSR regulatory system, which governs the cell envelope stress response (9, 10). As with Staphylococcus, the development of daptomycin resistance in Enterococcus typically requires an accumulation of mutations rather than a single genetic change.
Here, we report the rapid acquisition of high-level daptomycin resistance in a C. striatum strain isolated from a patient with an osteoarticular infection, who was initially treated with daptomycin following orthopedic surgery.
CASE PRESENTATION
A 49-year-old man was hospitalized in August 2022 for surgical management of chronic T11–T12 spondylodiscitis, associated with multiple rib lesions and bilateral pleural involvement. His medical history included paraplegia following a road traffic accident in 1993, with a subsequent development of a Charcot spine at T12–L1, complicated by septic pseudarthrosis secondary to multiple pressure ulcers. The initial spinal instrumentation placed after the 1993 accident was removed in 2002 due to infection. In 2014, the patient was treated for costal osteitis related to a subscapular pressure ulcer, with S. aureus and Proteus mirabilis identified from intraoperative samples. Management included surgical debridement and a 6 week course of antibiotic therapy with rifampin and levofloxacin. In 2019, the patient developed T11–T12 spondylodiscitis in the context of a new ischial pressure ulcer. S. epidermidis was isolated and treated with rifampin and doxycycline for 3 months. In the summer of 2022, the patient presented with fever. Blood cultures were positive for S. agalactiae, and imaging confirmed a recurrence of the T11–T12 spondylodiscitis, originating from the ischial pressure ulcer and complicated by associated rib and pleural involvement, treated by a 3 months course of antimicrobial therapy with amoxicillin (Fig. 1). Due to the need for both anterior and posterior surgical approaches as well as thoracic intervention, the patient was referred for the first time to our teaching hospital. In the spring of 2023, the first stage involved resection of the lateral pseudarthrosis and fixation. One month later, the second stage consisted of a posterior spinal fusion from T6 to L2, using bone graft harvested from the right iliac crest. Between the two surgical stages, the patient received intravenous antibiotic therapy consisting of piperacillin–tazobactam (4g × 3 /day IV) and high-dose daptomycin (10 mg/kg in a single daily infusion).
Chronological representation of antibiotic therapy and orthopedic surgical interventions from August 2022 to July 2023.
All first perioperative samples grew methicillin-susceptible S. aureus, methicillin-resistant S. capitis, and C. striatum susceptible to daptomycin (named CORY-1). Based on these results, piperacillin-tazobactam was shortly discontinued, and daptomycin was maintained in monotherapy until the second-stage surgery. After the second procedure, all five perioperative bone samples were positive for C. striatum resistant to daptomycin (named CORY-2). The antimicrobial therapy was immediately modified following the second intervention, and vancomycin was initiated. In light of the microbiological results and the favorable clinical and biological evolution, treatment was switched to oral linezolid after 10 days of intravenous vancomycin. After 3 weeks of linezolid therapy, the treatment was discontinued due to anemia and replaced with tedizolid. Under tedizolid, the anemia regressed, and the treatment was carried out to completion, for a total duration of 3 months.
Antimicrobial susceptibility testing
MICs for amoxicillin, meropenem, and imipenem were determined using E-test strips (BioMérieux) (Table 1), while MICs for vancomycin, teicoplanin, daptomycin, and dalbavancin were assessed by broth microdilution using Sensititre plates (Thermofisher, Dardilly, France), according to manufacturer’s recommendations. All MIC determinations were performed using a 0.5 McFarland standard inoculum. These tests were performed for both clinical C. striatum isolates (CORY-1 and CORY-2). Comparing the two isolates, we observed notable increases in MIC values: meropenem rose from 0.19 mg/L in CORY-1 to 0.75 mg/L in CORY-2, imipenem from 0.75 mg/L to 1.5 mg/L and vancomycin from 0.25 mg/L to 0.5 mg/L. Most strikingly, the MIC for daptomycin increased dramatically, from 0.12 mg/L to >32 mg/L (Table 1).
Whole genome sequencing
To understand the mechanism involved in daptomycin resistance in these C. striatum isolates, the two strains CORY-1 and CORY-2 were compared using genomic analysis through whole genome sequencing (WGS). The genomes of the two isolates were sequenced using Illumina short-read sequencing (NextSeq500 system, Illumina technology). Reads were assembled using Shovill v1.1.0 and SPAdes v3.14.0. To assess whether this recurrence came from the same bacterial strain or a completely independent infection, we analyzed the number of single nucleotide polymorphisms (SNPs) between the two strains using SNIppy v6.4.0. and found only 13 SNPs between the two strains. Among these 13 mutations, only three resulted in amino acid changes, each affecting three different genes (mltG, ftsl, and pgsA), while the remaining mutations were silent. The pgsA gene, which encodes phosphatidylglycerophosphate synthase involved in the synthesis of phospholipid acids in the bacterial membrane, harbored a G147D mutation. The mltG gene, which encodes an endolytic murein transglycosylase involved in peptidoglycan synthesis by acting as an elongation terminator, carried the T102A substitution. The ftsl gene, encoding penicillin-binding protein PBP3, a key player in cell division and peptidoglycan synthesis, carried an A370D substitution.
DISCUSSION
Acquired resistance to daptomycin in Corynebacterium species has been increasingly reported in recent years, particularly in C. striatum. Several clinical case reports and in vitro studies have described the ability of this organism to develop high-level resistance during antimicrobial therapy (11–13). Both clinical and experimental data have shown that resistance can emerge in 30–100% of isolates, depending on the strain and duration of treatment (6, 13), highlighting the capacity of this species to adapt under antibiotic pressure.
Our case provides additional evidence of this phenomenon. A patient treated for a chronic low-grade osteoarticular infection received 4 weeks of daptomycin therapy before a second isolate of C. striatum was recovered. Two clonally related isolates obtained 4 months apart exhibited an important increase in daptomycin MICs, from 0.12 mg/L to >32 mg/L. WGS confirmed that the two isolates, CORY-1 and CORY-2, were highly related, differing by only 13 SNPs, supporting the hypothesis of in vivo selection of resistance rather than reinfection by a distinct strain.
Three nonsynonymous mutations were identified between these two isolates. Among them, a nonsense mutation in pgsA, a gene encoding phosphatidylglycerol synthase, has previously been shown to confer high-level daptomycin resistance in Staphylococcus, Enterococcus, and Corynebacterium species (6, 14). Unlike Staphylococcus or Enterococcus, which typically require multiple mutations to achieve resistance (14), a single mutation in pgsA is sufficient in C. striatum. The G147D mutation identified in our case introduces a premature stop codon in the active site of the PgsA enzyme, abolishing phosphatidylglycerophosphate synthase activity. This results in a loss of phosphatidylglycerol, the primary target of daptomycin, thereby conferring resistance that has already been characterized (13). This mutation is consistent with previous reports: a recent systematic review investigating high-level daptomycin resistance in C. striatum showed that resistance typically emerged after 8–41 days of daptomycin therapy, and when explored, mutations in the pgsA2 gene were systematically involved (15). To date, no specific mutations have been consistently associated with daptomycin resistance in other Corynebacterium species.. However, mutations in the pgsA2 gene, previously linked to high-level resistance in C. striatum, have recently been described in other Corynebacterium spp., suggesting a potentially shared mechanism (16).
Two additional mutations were detected in mltG and ftsI, genes involved in peptidoglycan metabolism and cell division, respectively. These may contribute to the modest increase in β-lactam MICs observed in CORY-2, although their functional impact remains to be experimentally validated. Their presence may reflect compensatory mechanisms or adaptive cell envelope remodeling under selective pressure.
Together, these findings illustrate how a commonly used empiric regimen, although initially appropriate, may favor the selection of daptomycin-resistant subpopulations, especially in the context of chronic infections requiring prolonged empirical coverage and delayed microbiological diagnosis. This case underscores the need to reassess the use of daptomycin monotherapy in suspected C. striatum infections and to consider alternative agents or combination therapies early in the course of treatment.
To conclude, this case highlights the fact that, contrary to Staphylococcus spp. related infections, daptomycin should not be the antibiotic for C. striatum infections, especially in the case of infections requiring long-term treatment. Alternative agents, such as vancomycin or oxazolidinones, may represent a more reliable therapeutic alternative.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kalt F, Schulthess B, Sidler F, Herren S, Fucentese SF, Zingg PO, Berli M, Zinkernagel AS, Zbinden R, Achermann Y. 2018. Corynebacterium species rarely cause orthopedic infections. J Clin Microbiol 56:e 01200-18. doi:10.1128/JCM.01200-1830305384 PMC 6258853 · doi ↗ · pubmed ↗
- 2Trivedi GR, Merchant SS. 2024. Corynebacterium striatum: a true pathogen in chronic contiguous osteomyelitis. Canadian Journal of Infectious Diseases and Medical Microbiology 2024:1–8. doi:10.1155/2024/5020721 PMC 1085787638344026 · doi ↗ · pubmed ↗
- 3Verma R, Kravitz GR. 2016. Corynebacterium striatum empyema and osteomyelitis in a patient with advanced rheumatoid arthritis. BMJ Case Rep 2016:bcr 2016214691. doi:10.1136/bcr-2016-214691 PMC 478542626944378 · doi ↗ · pubmed ↗
- 4Söderquist B, Henningsson T, Stegger M. 2023. Corynebacterium striatum prosthetic joint infection successfully treated with long-term dalbavancin. Microorganisms 11:550. doi:10.3390/microorganisms 1103055036985124 PMC 10051730 · doi ↗ · pubmed ↗
- 5Zimmerli W, Trampuz A, Ochsner PE. 2004. Prosthetic-joint infections. N Engl J Med 351:1645–1654. doi:10.1056/NEJ Mra 04018115483283 · doi ↗ · pubmed ↗
- 6Mitchell KF, Mc Elvania E, Wallace MA, Droske LE, Robertson AE, Westblade LF, Burnham CA. 2021. Evaluating the rapid emergence of daptomycin resistance in Corynebacterium: a multicenter study. J Clin Microbiol 59:e 02052-20. doi:10.1128/JCM.02052-2033472898 PMC 8092723 · doi ↗ · pubmed ↗
- 7Peleg AY, Miyakis S, Ward DV, Earl AM, Rubio A, Cameron DR, Pillai S, Moellering RC, Eliopoulos GM. 2012. Whole genome characterization of the mechanisms of daptomycin resistance in clinical and laboratory derived isolates of Staphylococcus aureus. P Lo S One 7:e 28316. doi:10.1371/journal.pone.002831622238576 PMC 3253072 · doi ↗ · pubmed ↗
- 8Bayer AS, Mishra NN, Cheung AL, Rubio A, Yang S-J. 2016. Dysregulation of mpr F and dlt ABCD expression among daptomycin-non-susceptible MRSA clinical isolates. J Antimicrob Chemother 71:2100–2104. doi:10.1093/jac/dkw 14227121398 PMC 4954926 · doi ↗ · pubmed ↗