Cestode parasite accumulation in Octopus maya: Insights from an opportunistic sampling during the 2022 red tide event
Linda Yacsiri G. Marmolejo-Guzmán, Jhonny G. García-Teh, Karen Ascenet Arjona-Cambranes, Guadalupe Anai May-Sosa, M. Leopoldina Aguirre-Macedo

TL;DR
This study found that larger octopuses in the Yucatán Peninsula have more parasites, suggesting parasites accumulate as the octopuses grow.
Contribution
The study provides new insights into the host-parasite dynamics of Prochristianella sp. in Octopus maya during a red tide event.
Findings
Larger octopuses had significantly higher parasite counts (r = 0.85).
Parasite accumulation aligns with general host-parasite interaction patterns.
Octopus maya plays an ecological role as an intermediate host in its marine ecosystem.
Abstract
The infection dynamics of Prochristianella sp., a metacestode parasitizing the Yucatán Peninsula-endemic octopus, Octopus maya, were examined to explore the relationship between host size and parasitic infection parameters. During a red tide event, forty-nine octopuses were sampled from a single locality in the Yucatán Peninsula, México, and classified into three size classes. Infection metrics were assessed, including prevalence, mean intensity, abundance, and total parasite count. The results revealed a significant positive association (r = 0.85) between host size and the number of Prochristianella sp., with larger octopuses exhibiting higher infection parameters. The findings suggest that Prochristianella sp. progressively accumulates in O. maya throughout its very early life, aligning with the general patterns observed in host-parasite interactions. This finding is consistent with…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
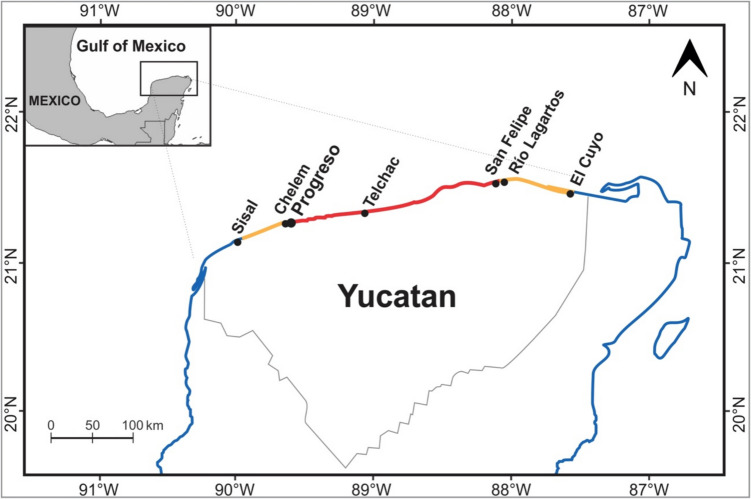 Figure 1
Figure 1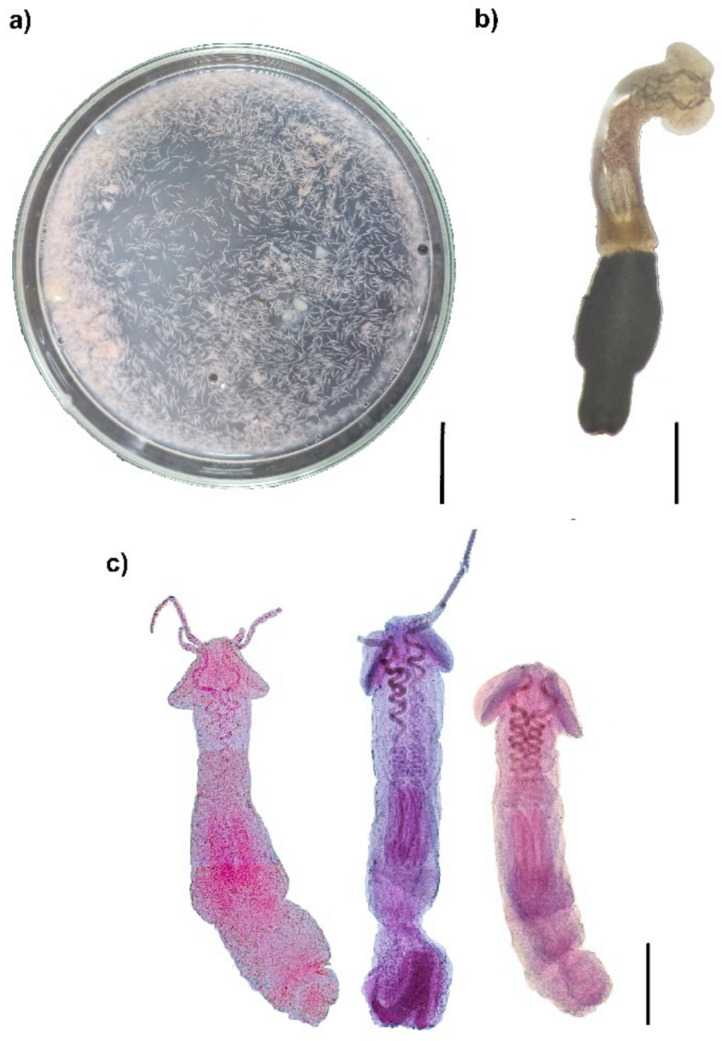 Figure 2
Figure 2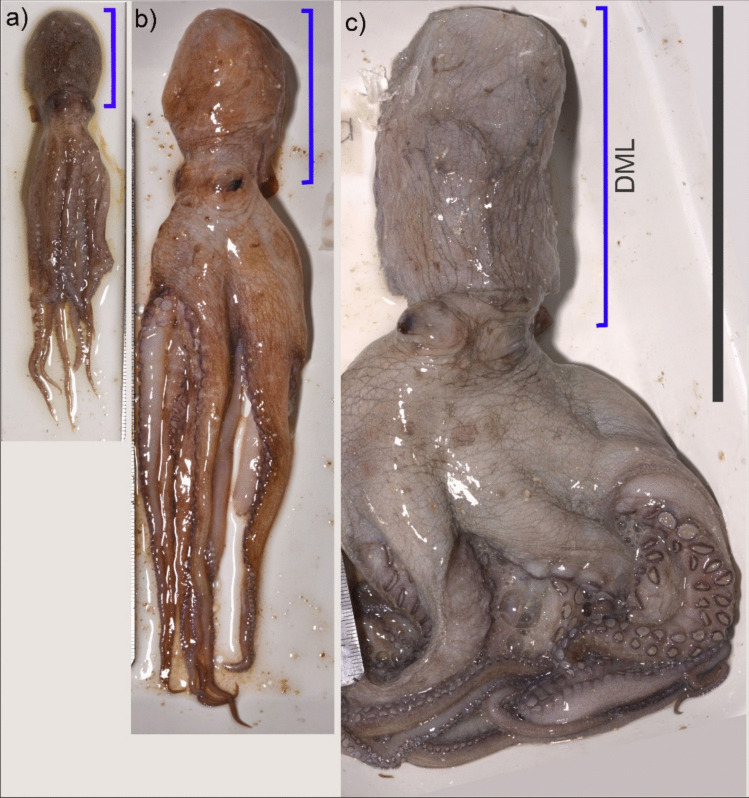 Figure 3
Figure 3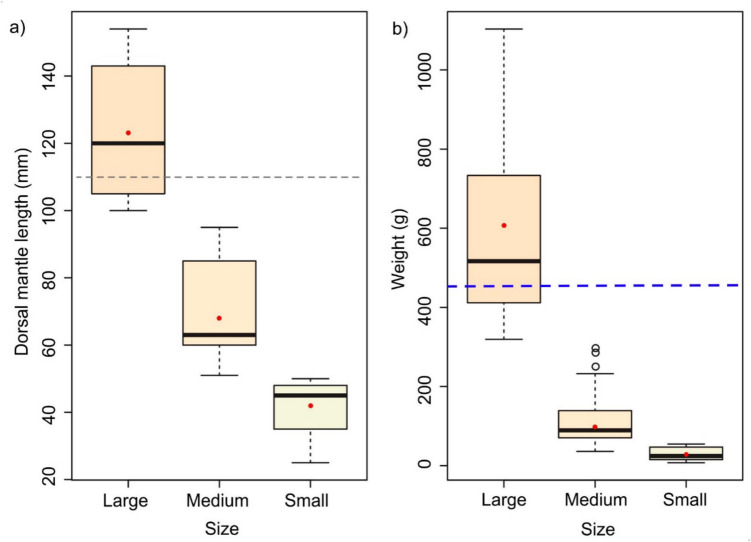 Figure 4
Figure 4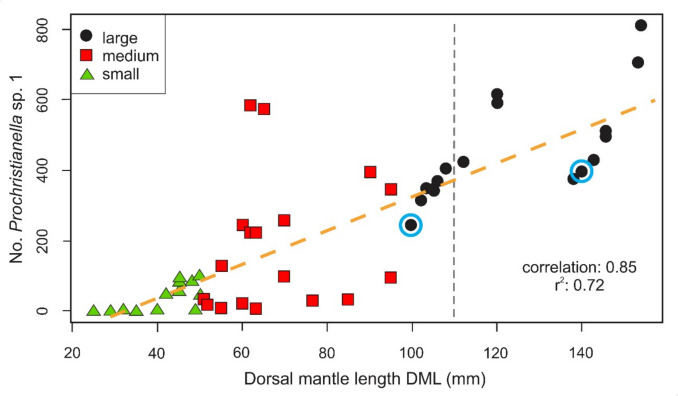 Figure 5
Figure 5- —Consejo Nacional de Humanidades, Ciencias y Tecnologías , México
- —https://doi.org/10.13039/501100008688Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional
- —https://doi.org/10.13039/501100003141Consejo Nacional de Humanidades, Ciencias y Tecnologías
- —MLA-M
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCephalopods and Marine Biology · Parasite Biology and Host Interactions · Marine Invertebrate Physiology and Ecology
Introduction
Endemic to the Yucatán Peninsula, México, Octopus maya is one of the most ecologically and economically important species in the region. It plays a crucial role in the marine ecosystem, sustaining the local fishing industry. Indeed, it is estimated to contribute over 60% of the total octopus catch (Avendaño et al. 2019; Coronado et al. 2020). The Yucatán octopus fishery is the largest in the Americas and is regulated by the National Fisheries Charter and Mexican Official Standards NOM-008-SAG/PESC-2015 (2015a) and NOM-009-SAG/PESC-2015 (2015b). These regulations ensure sustainable practices by setting a minimum capture size of 110 mm mantle length, prohibiting the use of hooks and spears, and enforcing a closed season from December 16 to July 31 (Rosas et al. 2014; Guarneros-Narváez et al. (2016). However, these regulations also limit scientific research opportunities, especially for collecting specimens under regulated sizes. The parasite fauna of O. maya comprises 20 parasite taxa, including seven cestode species, with Prochristianella sp. exhibiting the highest prevalence and abundance, affecting the octopus's health by causing tissue alterations and fibrosis in the anterior salivary glands (Guillén-Hernández et al. 2018a, b; Marmolejo-Guzmán et al. 2022).
Harmful algal blooms (HABs), or red tides, are spontaneous natural phenomena that appear on the coasts and significantly affect the marine ecosystem; their duration varies from days to months; their appearance is not limited to one time of the year; however, they are more common in summer (Rodríguez-Gil et al. 2007). These events result from the massive growth of phytoplankton due to favorable marine conditions, such as nutrient availability and specific currents and winds (Aguilar-Medrano et al. 2023; Hallegraeff 2023). Recently, non-toxic but harmful algal blooms have been observed along the Yucatán coast, causing significant fish and marine organism deaths (Rodríguez-Gil et al. 2007; Zetina-Ríos et al. 2009). The 2022 red tide, dominated by the dinoflagellate Cylindrotheca closterium, was a notable ecological disturbance that began at the port of San Felipe and progressed westward to Chelem, lasting approximately a week and resulting in widespread mortality of marine organisms, including specimens of O. maya (Aguilar-Medrano et al. 2023).
The 2022 red tide event offered an opportunity for O. maya sampling across a broad size range, overcoming the restrictions of Mexican Official Standards (NOM). This event resulted in numerous moribund and freshly deceased octopuses washing ashore, allowing us to collect specimens that span different developmental stages. This unprecedented sampling opportunity enabled a comprehensive study on the progression of infection of Trypanorhyncha cestodes (marine tapeworms with elasmobranch definitive hosts) in O. maya. By examining octopuses of a size gradient, we gain valuable insights into the prevalence, intensity, and pathological effects of these infections across different life stages of O. maya. These baseline data contribute to the ecological understanding of octopus-parasite systems, though direct fishery impacts remain unquantified.
Materials and methods
Specimen recollection
During the "red tide" phenomenon on the coasts of the state of Yucatán in August 2022, 34 moribund and visibly fresh dead specimens of the mayan octopus (O. maya) were recollected from the coasts of the municipality of Progreso, Yucatán, México (Fig. 1). The specimens were transported in cool boxes to the Aquatic Pathology Laboratory at Cinvestav Mérida, where they were preserved in freezers at −20 °C for subsequent examinations. Once in the laboratory, morphometric data were collected for each specimen, including dorsal mantle length (DML) in millimeters and weight in grams.Fig. 1. Main fishing ports in Yucatán affected by the 2022 red tide event. The red line represents the highly affected area. The orange line represents the medium-affected area. Map created by the authors, based on COBI (2022)
Helminthological examination
For the helminthological examination, the octopuses were dissected. Their organs (buccal mass, renal sacs, ink sac, stomach, siphon, spiral cecum, crop, heart, and gills) were examined on glass slides under a stereoscopic microscope (Motic SMZ-168) (Fig. 2a). Prochristianella sp. plerocercoids were encapsulated in a single structure (presumably elicited as a host response to Prochristianella sp. infection), primarily on the superior mandibular muscle of the buccal mass. At the same time, some free individuals were found in other digestive system organs (Marmolejo-Guzmán et al. unpublished data). All Prochristianella sp. individuals were separated, counted, and used for further analysis in this study.Fig. 2. Morphological confirmation of Prochristianella sp. cestodes. a) A Petri dish with Prochristianella sp. individual extracted from a single octopus, b) a detailed view of a single Prochristianella sp., c) individuals of Prochristianella sp. at different developmental stages within the plerocercoid phase. Bar scale: a) 2000 µm, b) 200 µm, and c) 200 µm
Some Prochristianella sp. individuals were fixed in vials with 4% formalin for staining purposes (others were preserved differently for additional analyses or discarded). From these, five randomly selected specimens per octopus (n = 34 hosts; total 170 individuals) were stained with Mayer-Schuberg's carmine technique, mounted in Canada balsam (Palm 2004), and identified as Prochristianella sp. according to Palm (2004) (Fig. 2b). All collected individuals were confirmed as Prochristianella sp. despite the varied developmental stages of the plerocercoid phase (Fig. 2c). The best-preserved specimens were deposited in the Helminthological Collection of Cinvestav Unit Mérida (CHCM) with accession codes CHCM-705, CHCM-705.2, and CHCM-705.3.
Size class categorization
Dorsal mantle length (DML) was measured for all specimens, with data presented as mean ± standard deviation (SD) for each size class. Octopus maya specimens were categorized into three size classes based on dorsal mantle length (DML): small (< 50 mm), medium (51–100 mm), and large (101–150 mm). The small size class corresponds to juvenile individuals, consistent with the developmental definition proposed by Rosas et al. (2014) beginning at 14 days post-hatching (DPH) and supported by wild growth rates of 0.425 mm/day (Solís and Chávez 1986), indicating these sizes represent immature individuals below sexual maturity thresholds (450 g ≈ 110 mm DML) (Rosas et al. 2014). Medium and large classes represent progressively mature stages, where the 110 mm DML minimum catch size (NOM-008-SAG/PESC-2015 2015a) aligns with both the modal size in wild populations (Nepita Villanueva and Defeo 2001) and documented reproductive maturity (Rosas et al. 2014).
Due to the sample limitation, only two of the 34 individuals collected during the red tide event belonged to the largest size category (101–150 mm DML). Consequently, morphometric and parasitological data of 15 additional O. maya specimens in the largest category from the fishing season of 2021 (August–December) were incorporated for the analyses. This inclusion was intended to enhance the representation of larger specimens.
Data analysis
To analyze the progression of Prochristianella sp. infections, the prevalence and abundance were calculated following the recommendations of Bush et al. (1997). Data on the prevalence and abundance of cestode infections across different classes of O. maya were statistically analyzed to understand the infection dynamics.
A nonparametric Kruskal–Wallis rank sum test was applied to detect differences between host size classes and the number of Prochristianella sp. individuals. Subsequently, the Dunn post-hoc test with Holm's adjustment method was applied to account for multiple comparisons. Additionally, Spearman's rank correlation was used to determine a possible significant association between octopus size (DML) (independent variable) and the number of Prochristianella sp. individuals (dependent variable). Finally, the coefficient of variation was calculated to determine the percentage of the total variation in the number of Prochristianella sp. individuals that was due to variation in octopus DML size (Sokal and Rohlf 1995). All statistical analyses were performed in R (R Development Core Team, http://www.R-project-com) using cort.test function and library "dunn.test". Statistical significance was set at α < 0.05 unless otherwise stated.
Results
Forty-nine specimens of O. maya were examined (Fig. 3). The dorsal mantle length (DML) ranged from 25 to 154 mm, with a mean of 81.1 ± 36.9 mm, and weight from 7.2 g to 1103.4 g, with a mean of 269.7 ± 298.1 g (Table 1). The dorsal mantle length and weights by size classes are presented in Fig. 4.Fig. 3. Specimens of Octopus maya collected during the 2022 red tide event in Yucatán, México. a) Small size specimen; b) Medium size specimen; c) Large size specimen. Scale bar = 110 mm; blue line = Dorsal Mantle Length (DML)Table 1. Infection parameters of Prochristianella sp. in Octopus maya are categorized by size classes based on dorsal mantle length (DML) from Progreso, Yucatán PeninsulaParametersOverallSmallMediumLarge(49)(13)(20)(16)Range DML (mm)25–15425–5051–100102–154DML (mm) \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\overline{X }$$\end{document} ±SD81.1 ± 36.941.1 ± 8.471.9 ± 16.7125.1 ± 19.7Weight (g) ± SD269.7 ± 298.129.1 ± 16.7145.3 ± 103.7620.8 ± 261.1Prevalence (%)9685100100Abundance \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\overline{X }$$\end{document} ±SD237 ± 22239 ± 38182 ± 180467 ± 144Intensity1–8122–971–585316–812No. Prochristianella sp.11,61950936357475 \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\overline{X }$$\end{document} -mean; SD-standard deviation; %-percentage; the numbers in parentheses correspond to the octopus examinedFig. 4Dorsal mantle length (a) and body weight (b) of Octopus maya across size classes. The red circle represents the mean values. Dashed lines indicate reference thresholds: grey for the minimum legal catch size (110 mm DML, NOM-008-SAG/PESC-2015 2015a), and blue for the average weight at sexual maturity (450 g, NOM-008-SAG/PESC-2015 2015a; Rosas et al. 2014)
While other cestodes (Onchoproteocephalidae and Trypanorhyncha species) were registered during the parasitological examination, this study focused exclusively on Prochristianella sp. All examined cestodes of this genus were confirmed to belong to Prochristianella sp., through morphological analysis (Fig. 2c), consistently identified across all size classes and developmental stages. While previous molecular analysis (28S rDNA) showed genetic similarity to Prochristianella sp. 1 (GenBank: DQ642769) (Marmolejo-Guzmán et al. 2022), formal morphological identification (particularly of tentacle armature patterns following Palm 2004) could not be completed due to suboptimal preservation conditions of the cestode specimens. We therefore cautiously designate these larvae as Prochristianella sp. (= Prochristianella sp. 1 from Marmolejo-Guzmán et al. 2022) until comprehensive morphological and molecular characterization can be performed.
Prochristianella sp. plerocercoid reached 96% prevalence, with an average abundance of 237 ± 222 individuals per examined O. maya. In total, 11,619 Prochristianella sp. plerocercoid individuals were recorded in this parasitological study (Table 1).
Infection progression was analyzed across the three previously defined octopus size classes: 13 specimens were classified in the small-size class (< 50 mm DML), 20 in the medium-size class (51–100 mm DML), and 16 in the large-size class (101–150 mm DML). A prevalence of 85% Prochristianella sp. infection was observed for the small-sized octopus group, with a mean abundance of 39 ± 38 individuals and an infection intensity ranging from 2 to 97 cestodes per infected host (Table 1). The medium-sized class exhibited a 100% prevalence, with a mean abundance of 182 ± 180 cestodes per octopus and an infection intensity ranging from one to 585 cestodes per infected host (Table 1). The large-sized class individuals were the most heavily infected, with a 100% prevalence, the highest mean abundance (467 ± 144 cestodes per octopus), and infection intensities ranging from 316 to 812 cestodes per infected host (Table 1).
Statistically significant differences were found between octopus size classes and the number of Prochristianella sp. individuals (Kruskal–Wallis, H (2,46) = 29.02; p = 4.98e-07). Dunn's multiple comparison post-hoc test revealed significant differences among all size classes (p < 0.05 for the comparisons). Additionally, a positive association (r = 0.85) was found between octopus size and the number of Prochristianella sp. individuals. Finally, it was found that 72% of the variation in the number of Prochristianella sp. can be attributed to changes in the octopus size (Fig. 5).Fig. 5. Relationship between Prochristianella sp*.* abundance and Octopus maya size classes. Significant differences in parasite abundance were observed among the three octopus size groups (Kruskal–Wallis, H (2,47) = 28.55; p = 6.31e-07). A strong positive correlation (r = 0.85) was identified, with octopus size explaining 72% of the variation in Prochristianella sp*.* numbers. The vertical dashed line (grey) represents the minimum catch size allowed (110 mm). The blue circle corresponds to the only large-sized individuals collected during the red tide
Discussion
The 2022 red tide on the north coast of the Yucatán Peninsula provided a valuable opportunity to investigate the progression of Prochristianella sp. infection during the early developmental stages of O. maya. For the first time, individuals measuring below the minimum legal capture size (110 mm) established by the Mexican Standard NOM-008-SAG/PESC-2015 were available for parasitological examination. The analysis of 34 specimens of O. maya, classified as small (< 50 mm), medium (51–100 mm), and large (101–150 mm), confirmed the infection of Prochristianella sp. at an early octopus stage, with a DML of 25 mm and a weight of 7.2 g. These results agree with those of Guillén-Hernández et al. (2018b, 2024) and Marmolejo-Guzmán et al. (2022), who found that Prochristianella sp. exhibited the highest cestode prevalence and abundance in O. maya, the octopus.
These results suggest that the life cycle dynamics and ecological interactions of Prochristianella sp. are particularly well-suited to its host, potentially enabling its accumulation and persistence throughout the octopus's life span. At the same time, the presence of highly infected hosts from the early life octopus stage (> 90%) highlights its role as a core parasite species of O. maya (Holmes 1991).
The progression of Prochristianella sp. infection in O. maya observed in this study aligns closely with the general characteristics of macroparasite infections described in the literature (Delahay et al. 2009; Guillén-Hernández et al. 2024). Macroparasites are known to induce chronic infections with short-lived or negligible immunity, which often facilitates re-infection in their hosts (Delahay et al. 2009).
Therefore, larger octopuses may accumulate higher parasite loads through ecological mechanisms including, 1) prolonged exposure to infected prey through trophic transmission (Guillén-Hernández et al. 2018a), 2) increased consumption of intermediate/paratenic host with age, and 3) seasonal fluctuations in host availability driven by temperature and photoperiod in tropical benthic systems (Coma et al. 2000). Complementing the work of Guillén-Hernández et al. (2024), who reported size-dependent abundance of Prochristianella sp. in O. maya across multiple life stages, including examination of gonadal development effects, this study specifically analyzes infection patterns in smaller octopuses (< 50 mm). These size-related differences in parasite load (< 50 mm) may suggest early-life exposure risks, possibly involving trophic pathways. However, conclusions remain limited by unconfirmed transmission mechanisms due to the unknown life cycle of Prochristianella sp. and the lack of physiological data in this study.
The strong positive relationship between O. maya size and Prochristianella sp. abundance (r^2^ = 0.72; p < 0.001) reflects progressive accumulation through the trophic transmission pathway mediated by host feeding ecology. Lower infection rates in smaller individuals (0–50 mm DML) likely reflect a reduced consumption of intermediate hosts, potentially due to size-limited prey selection or differing prey preferences compared to adults (Markaida 2023; Guillén-Hernández et al. 2024). As an intermediate consumer, O. maya acquires parasites through the ingestion of diverse prey, with larger hosts encountering more infected prey through ontogenetic dietary expansion**.** While crustaceans like Pitho spp. dominate the diet of adult O. maya in Campeche (Markaida 2023), the specific intermediate host responsible for Prochristianella sp. transmission remains unidentified, particularly in Yucatán populations where dietary studies are lacking. Aside from the fact that the host transmitting Prochristianella sp. to the octopus has not been identified, the result suggests that such an intermediate host may be an octopus regular prey item throughout its entire life.
Helminths such as cestodes and larval digeneans often exhibit a positive relationship with host size due to trophic transmission, as larger hosts consume more diverse and abundant prey (Price and Clancy 1983). This pattern is supported by meta-analyses showing that fish length correlates significantly with infection intensity in trophically acquired parasites such as cestodes and digeneans (Poulin 2000). However, effect sizes vary across species and systems. For instance, in Chromis cyanea, the relationship between host size and parasite load explains a broad range of variance (r^2^ = 0.11–0.72) (Fernández-Osorio et al. 2015). These parallels highlight that prolonged exposure and expanding habitat use primarily explain infection patterns across taxa (Sánchez et al. 2018), reinforcing the need for size-structured studies of trophic transmission.
The 2022 red tide mortality event enabled the first parasitological assessment of small Octopus maya below legal harvest size (110 mm DML), complementing previous work on adult infections (Guillén-Hernández et al. 2024). The strong positive correlation between host size and parasite abundance (r^2^ = 0.72) confirmed a progressive accumulation, most likely through both continuous exposure and increased consumption of infected prey, facilitated by expanded habitat use in larger octopuses. These findings establish an ecological baseline for monitoring potential parasite-mediated population effects under future environmental changes. Documenting infection dynamics across ontogeny may prove helpful if parasitism interacts with emerging stressors, such as shifting prey availability or habitat alterations. On the other hand, the effect of red tides on parasite transmission would require determining their impact on the availability of the first intermediate host, which can only be studied through a time series when another red tide occurs.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1COBI (2022) Impacto de la Marea Roja en las comunidades pesqueras de Yucatán en el verano de 2022. Comunidad y Biodiversidad A.C., Guaymas, México, p 14. https://cobi.org.mx/wp-content/uploads/2022/10/COBI-Reporte-Marea-Roja-web.pdf. Accessed 17 Feb 2025
- 2Fernández-Osorio RA, Corrada-Wong RI, Armenteros M (2015) Variaciones de la infracomunidad parásita durante la ontogenia de Chromis cyanea (Perciformes: Pomacentridae) en la costa norte de La Habana, Cuba. https://www.scielo.sa.cr/scielo.php?script=sci_arttext&pid=S 0034-77442015000300717. Accessed 20 Jan 2025
- 3Guarneros-Narváez PV, López-Rocha JA, Velázquez-Abunader I, Rosas Vázquez C (2016) Spatio-temporal analysis of the age distribution of Octopus maya in the Yucatán Peninsula. In: Proceedings of the 69th Gulf and Caribbean Fisheries Institute, Grand Cayman, Cayman Islands, pp 281–283
- 4Hallegraeff GM (2023) Harmful algal blooms: A global overview. In: Hallegraeff GM, Anderson DM, Cembella AD (eds) Manual on harmful marine microalgae, UNESCO, pp 25–49. https://unesdoc.unesco.org/ark:/48223/pf 0000136163. Accessed 20 Jan 2025
- 5Jereb P, Roper CFE, Norman MD, Finn JK (2016) Cephalopods of the world: An annotated and illustrated catalogue of cephalopod species known to date. Volume 3: Octopods and vampire squids. FAO Species Cat Fish Purp 4:1–370. https://openknowledge.fao.org/server/api/core/bitstreams/57241695-4904-470b-b 9e 2-ee 36fbe 1a 947/content
- 6Nepita Villanueva MR, Defeo O (2001) Crecimiento del pulpo Octopus maya (Mollusca: Cephalopoda) de la costa de Yucatán, México: un análisis de largo plazo. Rev Biol Trop 49:93–101. https://www.scielo.sa.cr/scielo.php?script=sci_arttext&pid=S 0034-77442001000100010. Accessed 18 Feb 2025
- 7NOM-008-SAG/PESC-2015 (2015 a) Para ordenar el aprovechamiento de las especies de pulpo en las aguas de jurisdicción federal Del Golfo de México y Mar Caribe. Pub. L. No. NOM-008-SAG/PESC-2015, DOF. https://www.dof.gob.mx/nota_detalle.php?codigo=5432972&fecha=13/04/2016#gsc.tab=0
- 8NOM-009-SAG/PESC-2015 (2015 b) Que establece el procedimiento para determinar las épocas y zonas de veda para la captura de las diferentes especies de la flora y fauna acuáticas, en aguas de jurisdicción federal de los Estados Unidos Mexicanos. Pub. L. No. NOM-009-SAG/PESC-2015 (2015), DOF. Available at: https://www.imipas.gob.mx/portal/documentos/publicaciones/BOLETINES/DOF-NOM-009-SAG-PESC-2015.pdf