One step, two functions: the mechanism of the ABCH class of transporter serving barrier construction and detoxification in insects
Bernard Moussian

Abstract
Click any figure to enlarge with its caption.
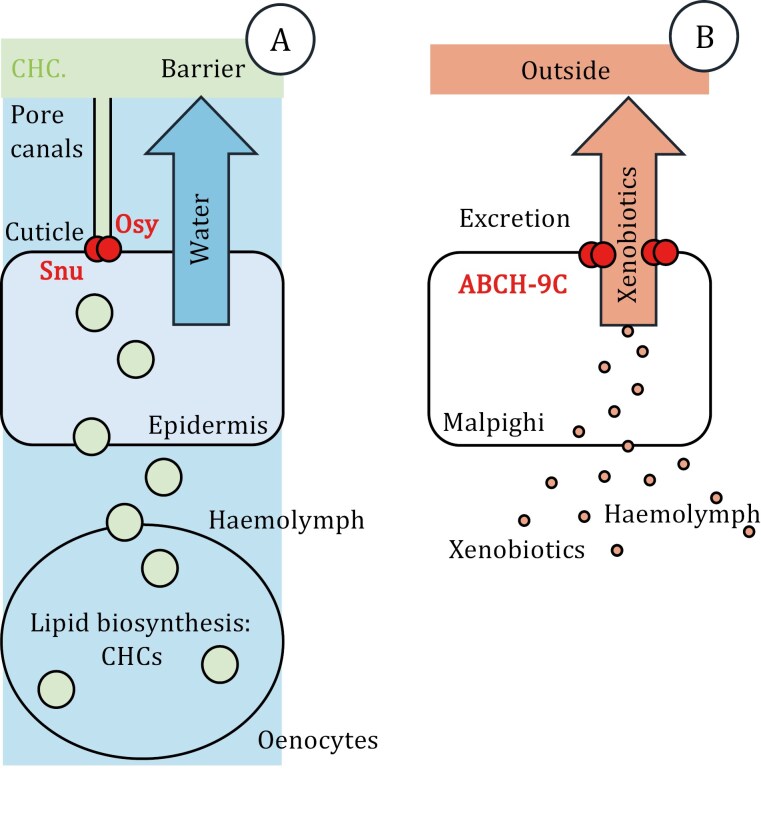 Figure 1
Figure 1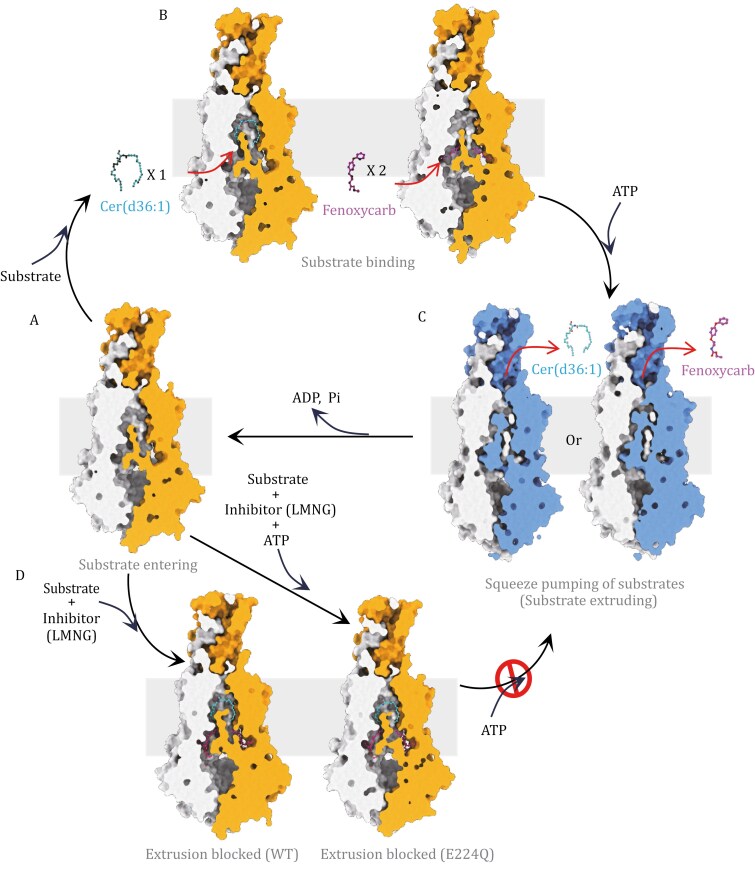 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDrug Transport and Resistance Mechanisms · Trace Elements in Health
Insects occupy a broad range of habitats after terrestrialization and radiation from marine crustaceans 519 million years ago (Schwentner et al., 2017). They found themselves in a new environment, foremost jeopardizing them with low humidity, i.e., dry or desiccation conditions. In addition, their co-evolution with land plants both as beneficial and maleficial companions exposed them to plant-produced xenobiotics (secondary metabolites) that as the plant defence molecules may have potential adverse effects on insects. Insects learned to cope with both situations, among others, through the evolution and activation of H-type ATP-binding cassette (ABC) transporters (Fig. 1). Indeed, ABCH transporters are arthropod-specific ABC transporters that, in all kingdoms of living organisms, using ATP hydrolysis, transport molecules from one side of a membrane to the other, thereby influencing the physiological milieus of both compartments.
ABCH transporters have been reported in various insect species to be important against desiccation. In the fruit fly Drosophila melanogaster, two ABCH transporters Snustorr (Snu) and Oskyddad (Osy) are essential for the construction of a cuticular surface barrier. This barrier consists of hydrocarbons (termed cuticular hydrocarbons, CHCs) that are transferred to the surface via pore and wax canals, which emanate from the apical plasma membrane of epidermal cells and run as nanotubes through the extracellular cuticle. CHCs are a blend of saturated, unsaturated and branched hydrocarbons with a chain length between 20 and 50 carbon residues that varies depending on the stage of the insect and the species. Mutations in the genes coding for these transporters, snu and osy, are embryonic and larval lethal in D. melanogaster (Wang et al., 2020; Zuber et al., 2018). This underlines that these transporters are essential. Their envelope, the outermost cuticle layer that harbours the CHCs is disintegrated probably accounting for the desiccation phenotype of the respective larvae. The CHC levels of flies with suppressed Snu or Osy function, in addition, are reduced. Moreover, Osy was found to be located in pore canals suggesting that CHC transport through the cuticle requires ABCH activity. Knock-down of the Snu ortholog in the pest migratory locust LmABCH-9C by RNA interference has a similar effect on CHC levels, survival and desiccation phenotype (Yu et al., 2017). Likewise, reduction of transcript levels of the Snu ortholog from the storage pest Tribolium castaneum enhances desiccation susceptibility (Broehan et al., 2013). Taken together, as D. melanogaster, T. castaneum and L. migratoria represent distantly related insect clades, ABCH transporters reflect a commonly shared evolutionary adaptation of insects to dry conditions by mediating CHC deposition needed for lipid barrier construction on the cuticle surface.
The second challenge that insects encountered during their evolution, i.e., detoxification against plant secondary metabolites was seemingly also accompanied by the evolution of ABCH transporters. In T. castaneum and the small brown planthopper, Laodelphax striatellus up-regulate the expression of ABCH coding genes when exposed to xenobiotics such as diflubenzuron or chlorpyrifos suggesting their involvement during xenobiotic detoxification by lowering their concentration within the cell (Rösner et al., 2021; Sun et al., 2017). The actual tissue(s) responding to xenobiotic exposure was not identified; indeed, the role of ABCH transporters during detoxification and their position in the underlying pathways has not been studied in detail.
Recently, Jinli Chen, Yanwei Duan, Yuanyuan Zhou and Qing Yang published a landmark article in Cell that unravels the mechanism of ABCH transporter function in transporting molecules across membranes (Chen et al., 2025). Using cryo-electron microscopy (cryo-EM), they elegantly showed that ceramide and the insecticide fenoxycarb are transported by the TcABCH-9C transporter applying a squeeze-pump mechanism (Fig. 2). As a half transporter, TcABCH-9C homo-dimerizes to constitute a full transporter forming a narrow and arched substrate-binding tunnel that is long enough to harbour lipids such as ceramide or two short lipophilic molecules such as fenoxycarb. Upon binding of the substrate on the cytosolic side of the membrane and ATP binding at the nucleotide-binding domain of the protein, the tunnel crushes and the substrate is squeezed to the vent at the other side of the membrane and pumped into the target compartment. This squeeze-pump mechanism is novel for ABC transporters and possibly unique for insect ABCH transporters indicating the evolution of it in conjunction with the double need for the transport of molecules against desiccation and for xenobiotic tolerance. Excitingly, the authors found that the amphiphilic detergent lauryl maltose neopentyl glycol (LMNG) is able to attenuate TcABCH-9C activity in liposomes by inhibiting both ATPase activity and, by consequence, substrate transport. This landmark finding opens the avenue for the development of a new class of LMNG-based insecticides that should be selective for insect ABCH transporters and therefore environmentally friendly. This is urgently needed as with climate change and its effects on biodiversity dynamics, we require a more sophisticated approach to meet human society and environment with more respect and care. In summary, the detailed elucidation of the mechanism of TcABCH-9C function is, hence, a milestone not only in the cell biology of ABC transporters in general but particularly in the research field of pest management in agriculture.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Broehan G , Kroeger T, Lorenzen M et al Functional analysis of the ATP-binding cassette (ABC) transporter gene family of Tribolium castaneum. BMC Genomics 2013;14:6.23324493 10.1186/1471-2164-14-6PMC 3560195 · doi ↗ · pubmed ↗
- 2Chen J , Duan Y, Zhou Y et al Squeeze pumping of lipids and insecticides by ABCH transporter. Cell 2025;188:944–957.e 19.39721587 10.1016/j.cell.2024.11.033 · doi ↗ · pubmed ↗
- 3Rösner J , Tietmeyer J, Merzendorfer H. Functional analysis of ABCG and ABCH transporters from the red flour beetle, Tribolium castaneum. Pest Manag Sci 2021;77:2955–2963.33620766 10.1002/ps.6332 · doi ↗ · pubmed ↗
- 4Schwentner M , Combosch DJ, Pakes Nelson J et al Phylogenomic solution to the origin of insects by resolving crustacean-hexapod relationships. Curr Biol 2017;27:1818–1824.e 5.28602656 10.1016/j.cub.2017.05.040 · doi ↗ · pubmed ↗
- 5Sun H , Pu J, Chen F et al Multiple ATP-binding cassette transporters are involved in insecticide resistance in the small brown planthopper, Laodelphax striatellus. Insect Mol Biol 2017;26:343–355.28299835 10.1111/imb.12299 · doi ↗ · pubmed ↗
- 6Wang Y , Norum M, Oehl K et al Dysfunction of oskyddad causes harlequin-type ichthyosis-like defects in Drosophila melanogaster. P Lo S Genet 2020;16:e 1008363.31929524 10.1371/journal.pgen.1008363 PMC 6980720 · doi ↗ · pubmed ↗
- 7Yu Z , Wang Y, Zhao X et al The ABC transporter ABCH-9C is needed for cuticle barrier construction in Locusta migratoria. Insect Biochem Mol Biol 2017;87:90–99.28610908 10.1016/j.ibmb.2017.06.005 · doi ↗ · pubmed ↗
- 8Zuber R , Norum M, Wang Y et al The ABC transporter Snu and the extracellular protein Snsl cooperate in the formation of the lipid-based inward and outward barrier in the skin of Drosophila. Eur J Cell Biol 2018;97:90–101.29306642 10.1016/j.ejcb.2017.12.003 · doi ↗ · pubmed ↗