Yeast Midas’ touch: recent advances in the valorisation of methanol by yeasts
Alessandra Mauri, Lorenzo García Tejada, Lars M. Blank

TL;DR
This review discusses how yeast can convert methanol into valuable chemicals, offering a sustainable alternative for industrial biotechnology.
Contribution
The paper highlights recent advances in using methylotrophic yeasts for methanol valorisation and their potential in sustainable chemical production.
Findings
Methylotrophic yeasts can metabolize methanol as their sole carbon and energy source.
Genetic engineering has enabled these yeasts to produce high-value compounds like organic acids and recombinant proteins.
Methanol can be sustainably sourced from biomass or CO2 hydrogenation using green H2.
Abstract
Traditionally, methanol has been used as a building block for a plethora of applications. It holds a key role in the chemical industry to produce paints, fabric, solvents, and plastics, and in producing biodiesel as the alcohol counterpart in the transesterification of plant oils and animal fats. Recently, methanol regained attention in industrial biotechnology due to its potential as an alternative substrate. Specifically, methanol is natively metabolised by a group of microorganisms called methylotrophs. Endowed with peculiar physiology, these can utilise the reduced C1 molecule as their sole carbon and energy source. While efforts to implement methylotrophic metabolism in heterologous hosts are still in its infancy, genetic engineering of methylotrophic yeast already succeeded in the conversion of methanol into high-value compounds, such as organic acids or recombinant proteins,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
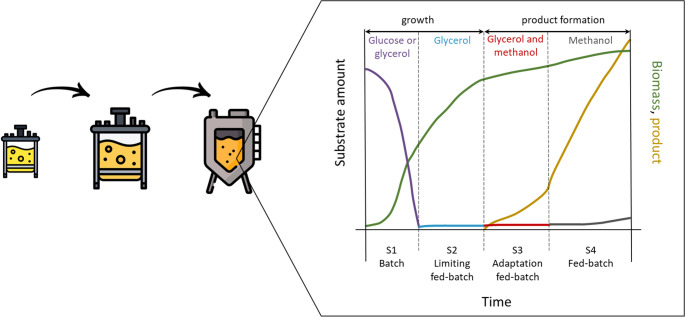 Fig. 3
Fig. 3- —RWTH Aachen University (3131)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial metabolism and enzyme function · Microbial Metabolic Engineering and Bioproduction · Biofuel production and bioconversion
Introduction: envisioning the revolution of a “land-free” biotechnology
Yeast fermentations using sugar as carbon and energy source are dominating food and biofuel production and contribute to the production of many other products. Land use for sugar production, which will be discussed later, is a central aspect for the future of microbial biotechnology. As an anecdote, the increase from 5% to 10% bioethanol in gasoline was a major communication disaster in Germany, with claims that the local bread roll will increase in price, with the consequence that 15 years later E10 fuels are still lower in sale despite an up to 5% price advantage. Many of us remember the tortilla crisis from 2007 in Mexico, although later debunked for the direct connection to bioethanol production in the USA (Keleman and Rañó 2012). With the absolute need to defossilise the chemical industry to meet CO_2_ reduction targets, the use of microbial biotechnology will massively increase, as we partly see already in China. There, with world leading companies in many of the microbially produced products, from vitamins to bulk amino acids, from specialty chemicals to bulk carboxylic acids, the amount of sugar fermented is in the double-digit percent of total sugar available, raising concern of further growth. While sugars dominate the recent growth in fermentation capacities, the research and to some extent production, is exploring other carbon and energy sources, such as CO_2_ + H_2_ (Bernal-Cabas et al. 2025; Daniell et al. 2012; Panich et al. 2021), plastic monomers (Liu et al. 2025; Tiso et al. 2021; Welsing et al. 2025), acetate (Ullmann et al. 2021; Ziegler et al. 2024), and as here discussed, methanol. In the short run, methanol will not contribute to the agreed-on Paris Climate goals, as it is synthesized from fossil resources such as gas or oil or in China even from coal, causing a significant CO_2_ footprint. However, with cheap methanol available that is competitive with sugar prices, firms can invest in the technology and can further optimise the value chain to reduce cost. In the mid to long term, green methanol, i.e., methanol produced from CO_2_ and green hydrogen will be available. The prerequisite is sustainable electricity from wind and solar or other routes like water dams, to split water into O_2_ and the required green H_2_. The point sources of the future for CO_2_ are expertly discussed elsewhere, while we would like to mention that the O_2_ from water splitting will intensify the fermentation industry, as higher production rates will be possible using oxygen enriched air. Safety measures must be taken into account, though. Green methanol as carbon and energy source for biotechnology can be seen as a carbon capture and utilization (CCU) technology. While companies like Solarfoods (https://solarfoods.com) showcase the technological feasibility of direct air capture for biotechnology, the economics argue clearly for CO_2_ point sources, such as biogas plants and cement kiln. Compared to other CCU uses, the replacement of sugar by green methanol comes with a stark reduction in land use, which comes close the “land free” biotechnology. Instead of huge farmland use for corn, sugar beet and sugar cane, land is mainly required for solar panels and windmills, while biogas and water dams contribute to energy storage possibilities. In Europe, farmland use for direct food production is below 25% and, world-wide, on average below 35%, while feed production is as high as 60%. A change in human diet from meat and dairy to more plant-based would allow increased land use for biotechnology, without challenging food security. However, with ever improving wind and solar technologies that are predicted to allow electricity production at rates as low as 1.5 dollar cents per kWh in favourable world regions, like south Chile or some parts of the Arabian peninsula, the here presented “land free” biotechnology using green methanol will become a reality. Ideally, some of the agricultural land is used for renaturing to slow down and finally stop the loss of biodiversity.
We here briefly introduce the current advances and advantages of green methanol as a carbon source for bioprocesses, followed by the metabolic pathways required for it, before summarizing literature examples, and ending with a weighted outlook on the intriguing carbon source methanol.
Methanol, an established building block and commodity
Methanol (CH_3_OH), or methyl alcohol, is the simplest alcohol and a fundamental building block in the chemical industry. Interest in the molecule rose in the 1980 s to utilise it as a fuel alternative or as a platform chemical for chemical conversion onto other simple molecules, such as formaldehyde, acetic acid, methyl methacrylate, and methylamines (IRENA & Methanol Institute 2021; Olah 2005). Another application for methanol resides on its utilization as feedstock for bioprocesses. As a reduced C1 molecule with a lower oxidation state than glucose or sucrose and thus a higher energy density per molecule, it can be utilized as both energy and carbon source for microorganisms. This ability to utilize methanol has been long known for and studied in bacteria (Abou-Zeid and Baghlaf 1983; Söhngen 1905). Methanol metabolism in a yeast species was first described only in the late 1960 s, synchronously to the oil boom (Ogata et al. 1969). This research was founded on the economical availability of petrochemical compounds, with microbial protein production from reduced C1 compounds materializing in the form of single-cell protein (SCP) for food and feed supply (Abou-Zeid and Baghlaf 1983; Wegner 1990). However, the fluctuating prices of these feedstocks combined with cheaper product alternatives rendered methanol-based processes unfeasible for the production of SCP or bulk chemicals, and only the processes that produced high added value products like enzymes remained relevant (Cereghino et al. 2002; Macauley-Patrick et al. 2005).
Methanol is mainly produced using fossil sources in industrial settings, i.e., from synthesis gas, which is in turn derived from natural gas or coal (Olah 2005). Given the current desire to step away from non-renewable energy sources, the increasing availability of the solvent as experienced in the last decades of the twentieth century is highly compromised. Despite the syngas-based process being the gold standard, non-renewable feedstocks are responsible for GHG emissions and experience high volatility in their price and market due to their finite nature. Therefore, new feedstocks have been investigated, and innovative processes have been successfully developed to produce methanol in a greener, i.e., more sustainable way (Table 1). Based on the production process, different classes of methanol can be distinguished. Green methanol is generated by processes of renewable origin, like biomass gasification, or electrolysis fuelled by renewable electricity, whereas blue methanol is produced from non-renewable processes from natural gas with Carbon Capture Storage (CCS) (Madejski et al. 2022). Grey and brown methanol are exclusively produced through non-renewable processes utilizing natural gas and gasified coal, respectively, without CCS. The technologies employed to produce methanol can be summarized in two approaches: conversion through the catalysis of syngas, and the hydrogenation of carbon dioxide (CO_2_). In the first approach, either natural gas under steam reformation, gasified coal or biomass exposed to higher temperatures, are converted into syngas, a mixture of carbon monoxide and molecular hydrogen. This gas mixture is then further catalysed into methanol. The second approach involves a direct methanol synthesis (DMS) from CO_2_ and molecular hydrogen through the utilisation of a catalyst. In Table 1 we provide a comprehensive overview of the industrial processes currently known to produce methanol. Methanol is here classified based on the origin of the feedstocks and the synthesis process, along with the chemical reactions and the catalysts involved (Roy et al. 2018; Gautam et al. 2020; Shanmugam et al. 2021).
Table 1. Overview of the different feedstocks and feedstock origin, respective processes and specifications currently in use to produce methanol industriallyFeedstockOriginProcessReactionsCatalystsType of CH_3_OHNatural gasFossilSteam methane reforming, methanol synthesisCCUS \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:{CH}_{4}\:+\:{H}_{2}O\:\to\:\:CO\:+3{H}_{2}$$\end{document}
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:CO+2{H}_{2}\to\:C{H}_{3}OH\:$$\end{document}
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:{CO}_{2}\:+\:3{H}_{2}\:\to\:\:C{H}_{3}OH\:+{H}_{2}O$$\end{document} NiCu/ZnO/Al₂O₃grey, blueCoalFossilGasification of coal, methanol synthesis \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:C\:+\:O_2\:\to\:\:CO\:/\:CO_2$$\end{document}
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:C\:+\:H_2O\:\to\:\:CO\:+\:H_2$$\end{document}
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:{CO\:+\:2{H}_{2}\:\to\:\:CH}_{3}OH$$\end{document}
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:{CO}_{2}\:+\:3{H}_{2}\:\to\:\:C{H}_{3}OH\:+{H}_{2}O$$\end{document} Cu/ZnO/Al_2_O_3_Cu/ZnO/Al_2_O_3_brownCO_2_ + green H_2_CapturedCatalytic hydrogenation \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:{CO}_{2}\:+\:3{H}_{2}\:\to\:\:C{H}_{3}OH\:+{H}_{2}O$$\end{document} Cu/ZnO/Al₂O₃green (e-)CO_2_CapturedPhotocatalytic conversion \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:C{O}_{2}\:+\:6{H}^{+}\:+\:6{e}^{-}\:\to\:C{H}_{3}OH\:+\:{H}_{2}O$$\end{document} TiO_2_/Pt, Cu-TiO_2_grey, green (e-)CO_2_CapturedElectrocatalytic reduction \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:C{O}_{2}\:+\:6{H}^{+}\:+\:6{e}^{-}\:\to\:C{H}_{3}OH\:+\:{H}_{2}O$$\end{document} Pt, Pd, Ru/Na- or K-modified β-aluminagrey, green (e-)BiomassIndustrial wasteGasification of biomass, methanol synthesis \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:{C}_{x}{H}_{y}{O}_{z}\:+\:{O}_{2}/{H}_{2}O\:\to\:CO\:+\:H2+\:CO2\:+\:CH4$$\end{document}
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:{CO\:+\:2{H}_{2}\:\to\:\:CH}_{3}OH$$\end{document}
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\:{CO}_{2}\:+\:3{H}_{2}\:\to\:\:C{H}_{3}OH\:+{H}_{2}O$$\end{document} Fe–Cr, Cu–ZnCu/ZnO/Al₂O₃green (bio-)
Old vs. new feedstocks: methanol and the “land-free” revolution
Since the 1970 s, when its synthesis from methane became inexpensive, paving the way for yeast biomass and high-protein animal feed production, methanol has been regarded as an ideal fermentation substrate (Higgins and Cregg 1998). Due to the price increase of natural gas that plagued the same decade, the industry shifted its focus to other substrates, which were inexpensive and had high availability, such as sugar-based feedstocks. Nevertheless, using these feedstocks for non-food purposes, e.g., biofuels or chemical production, poses an ethical dilemma (Srinivasan 2009). Moreover, the biodiversity loss linked to extensive land usage is a further side effect (Oakley and Bicknell 2022). Hence, the industry’s interest partly shifted to second-generation (2G) feedstocks, which are mainly represented by lignocellulosic biomass. Despite not competing with the food sector, second-generation feedstocks present non-negligible drawbacks (Monlau et al. 2012). Due to its heterogeneous composition, lignocellulosic biomass processing is time- and energy-demanding. Logistics is highly challenging and requires new and highly developed local infrastructures (Monlau et al. 2012; Si et al. 2016). Also, the heterogeneous composition increases processing cost as non-fermentable residues must be removed either up- or downstream of the fermentation process, which is particularly challenging for non-volatile products (Fig. 1).Fig. 1. Schematic comparison of first, second and third generation feedstocks
Third-generation (3G) feedstocks are hoped to reduce processing cost, preserve land, water and biodiversity, and tackle greenhouse gas emissions (GHG). These feedstocks are represented by captured carbon CO_2_ coupled with the use of renewable energies, such as electricity generated by wind power and photovoltaic cells (Liu et al. 2020, 2023). Atmospheric CO_2_ is an abundant feedstock: its levels have risen rapidly over the last century, reaching over 420 ppm in 2022, significantly exceeding the 350 ppm limit set at the Paris Climate Conference in 2015 (Garcia et al. 2022; NASA, Global Climate Change 2025). Hence, CCS from power-plant streams, industrial streams, or the atmosphere is a prominent approach and benefits from a well-established technology that addresses two major issues at the same time: alternative feedstock provision and GHG emissions. The fate of the carbon captured is double: it can be utilised (CCU) as a 3G feedstock to produce goods or it can be stored in geological sinks (CCS) until future utilisation. Comprehensively, this technology is known as Carbon Capture Utilisation and Storage (CCUS) (Garcia et al. 2022).
Despite significant technological advancement, the industrial production of green methanol is still not competitive compared to the fossil fuel-derived methanol and presents limitations. Specifically, the industrialisation bottlenecks are: (i) costly and complex carbon-free H_2_ supply, (ii) low process productivity and selectivity and (iii) catalysts performance limitations (Lais et al. 2018; Ye et al. 2019). Current costs of green hydrogen via electrolysis are typically 1–1.5 per kg for fossil-based hydrogen (Global Hydrogen Review 2022). Since hydrogen accounts for up to 70% of green methanol’s production cost, this cost disparity represents the most critical barrier to competitiveness. Furthermore, large-scale deployment would require massive renewable electricity inputs, as producing 1 ton of methanol consumes ~ 10–12 MWh (Kang et al. 2021). Conventional Cu-ZnO catalysts typically achieve methanol selectivity of 30–70% at CO_2_ conversion below 30% under standard reaction conditions. To address these limitations, new catalyst formulations, such as Cu–ZnO–Al_2_O_3_ and In-based materials, were developed showing improvement in selectivity, but unvaried methanol yields (Roy et al. 2018). Furthermore, at the plant-scale productivity is typically 0.1–0.5 g MeOH/g catalysth, with only a few catalysts reaching 1.0 g/g catalysth (Wesner et al. 2023; Araújo et al. 2024). Collectively, these three bottlenecks restrict the scalability and economic viability of green methanol production. Nonetheless, the production of green methanol is expected to pave the way to a “land-free” scenario, a term we aim to introduce in this review. Coupling goods production and use of alternative feedstock would mitigate land shortages for food and feed, while addressing GHG emissions. Moreover, a “land-free” approach would promote sustainable development and environmental stewardship in an era of growing awareness for resource conservation.
Methanol metabolism and regulation
As an alcohol with low molecular weight, methanol poses toxicity issues to microbial cells, i.e., by modifying the fluidity of the cell membrane and interfering with the stability of proteins inside (Bennett et al. 2021; Wang et al. 2020). Inside the cell, it can be also enzymatically converted into compounds like formaldehyde or formic acid, which also pose toxicity to cell physiology. Formic acid inhibits the cytochrome c oxidase of the electron transport chain in certain yeast and bacterial species, causing a disbalance in the acid-base equilibrium and sending the cells into oxidative stress and, ultimately, apoptosis (Du et al. 2008; Kassem et al. 2017; Lee et al. 2010). Formaldehyde exerts a stronger toxicity in cells by performing protein and DNA cross-linking, which leads to impaired DNA replication, protein translation and reduced protein activity, sends cells into stress response and ultimately causes cell death (Chen et al. 2016; Jayakody and Jin 2021; Wani and Jain 2024; Yu et al. 2015).
Some microorganisms are able to metabolize methanol in spite of its inherent toxicity, an example of methylotrophy. Methylotrophy comprises a series of metabolic pathways through which highly reduced C1 compounds like methane or methanol are assimilated by microorganisms to obtain energy or funnel the carbon molecules into their catabolism (Abou-Zeid and Baghlaf 1983; Large 2012; Wegner 1990; Yurimoto et al. 2011). Bacterial species can utilize a number of these compounds through different metabolic pathways and have more synthetic modules available to increase the energetic efficiencies of their assimilation (Chistoserdova et al. 2009; Kremp and Müller 2021; Krüsemann et al. 2023). Regarding methanol, combinations of different pathways like the ribulose monophosphate (RuMP) or the reductive glycine, and engineered cofactor systems, like NAD- or quinone-dependent, enable an optimisation of methanol assimilation where the reductive power is conserved and better utilized into cell energetics (Krüsemann et al. 2023; Singh et al. 2022; Wagner et al. 2023). Methylotrophic bacteria, however, have a lower resistance to the toxic intermediates generated in the assimilation process and release high amounts of heat as a result of their rapid growth, both traits that are generally undesirable in bioprocesses. Contrarily, methylotrophic yeasts can exclusively process methanol via the native peroxisome-localized Methanol Utilization (MUT) Pathway involving the Xylulose Mono-Phosphate (XuMP) cycle (Fig. 2A). This pathway consists of three major steps: the initial conversion of methanol to formaldehyde, its subsequent dissimilation into energy and CO_2_, or its assimilation through C3 intermediates into cell components and further metabolic products. The initial conversion to formaldehyde is typically performed by an alcohol oxidase (AOX), though research has found that, in some species, a promiscuous alcohol dehydrogenase (ADH) can also carry out the conversion in the absence of AOX (Yurimoto et al. 2011; Zavec et al. 2021).Fig. 2**A **Localization of the Methanol Utilization Pathway (MUT) within subcellular compartments in methylotrophic yeasts. Metabolites: MeOH, methanol; FALD, formaldehyde; GS, glutathione; FORM, formate; Xu5P, xylulose-5-phosphate; DHA, dihydroxyacetone; GAP, glyceraldehyde-3-phosphate; DHAP, dihydroxyacetone-3-phosphate; F1,6BP, fructose-1,6-bisphosphate; F6P, fructose-6-phosphate; E4P, erythrose-4-phosphate; S1,7BP, sedoheptulose,1–7-bisphosphate; S7P, sedoheptulose-7-phosphate; R5P, ribose-5-phosphate; Ru5P, ribulose-5-phosphate; PYR, pyruvate. Enzymes: ADH, Alcohol dehydrogenase; AOX, alcohol oxidase; CTA1, catalase; FLD, formaldehyde dehydrogenase; FGH, S-formylglutathione hydrolase; FDH, formate dehydrogenase; DAS, dihydroxyacetone synthase; DAK2, dihydroxyacetone kinase; FBA1-2, fructose-bisphosphate aldolase; FBP1, fructose-bisphosphatase; SHB17, sedoheptulose-1,7-bisphosphatase; RKI1-2, ribose-5-phosphate ketol-isomerase; RPE1-2, D-ribulose-5-phosphate-3-epimerase; TPI1, triose-phosphate isomerase; ATP, adenosin triphosphate; ADP; adenosin diphosphate; Pi, phosphate. **B **Stoichiometric balance of the condensation reactions for formaldehyde assimilation through the XuMP cycle
AOX activity relies on O_2_ as an electron acceptor, which is converted to highly reactive hydrogen peroxide (H_2_O_2_). Being a highly oxidizing agent, hydrogen peroxide can pose a dire threat to cellular homeostasis through protein inactivation, membrane disruption or DNA damage (Finnegan et al. 2010; Juven and Pierson 1996). As part of a highly specialized system to deal with this oxidative stress in methylotrophic yeasts, AOX activity is localized within peroxisomes alongside catalase (CAT) to detoxify its H_2_O_2_ by-product into H_2_O and O_2_. (Rußmayer et al. 2015; van der Klei et al. 2006). This localization enables the rapid removal of H_2_O_2_ within the cell by spatially coupling both enzymatic reactions, thereby minimizing the damage done to critical biomolecules and organelles. A second mechanism to reduce oxidative stress within the peroxisome for reactive oxygen species (ROS) scavenging is performed by the glutathione-dependant peroxiredoxin Pmp20 (Bener Aksam et al. 2008; Berrios et al. 2022; Horiguchi et al. 2001). The enzyme reduces hydroperoxides into their non-ROS variants, thus preventing their oxidative capabilities. Mutant yeast cells lacking Pmp20 but expressing CAT have been shown to experience stronger growth impairments than CAT-negative mutants; this emphasises the critical action of the enzyme on mitigating oxidative stress and might point at an underlying activity on general ROS removal (Horiguchi et al. 2001; Yurimoto et al. 2011). Another detoxifying step tackles the non-enzymatical hemiacetal formation of methanol and formaldehyde, which is oxidized to methyl formate by a cytosolic alcohol dehydrogenase (ADH) with methyl formate synthase activity (Murdanoto et al. 1997; Yurimoto et al. 2004).
The produced formaldehyde can then follow a dissimilation route where it is fully oxidized to CO_2_ in the cytosol (Berrios et al. 2022; Yurimoto et al. 2005). It leaves the peroxisome following a non-enzymatic condensation with reduced glutathione and is then subject to two consecutive dehydrogenation reactions to formate and CO_2_ with the co-production of two NADH molecules. A cytosolic system for the regeneration of reduced glutathione is also found in methylotrophic yeast (Yano et al. 2009). Flux Balance Analysis (FBA) studies have shown that a very high percentage (> 50%) of the methanol fed to the cells is dissimilated into CO_2_, especially in protein production processes, where an upregulation of the TCA cycle has also been studied (Jordà et al. 2012; Krüsemann et al. 2023; Leticia Vanz et al. 2012; Singh et al. 2022). Different growth rates and methanol uptake rates, as well as cofeeding strategies also have a direct effect on the amount of methanol directly dissimilated to CO_2_, sometimes rising to values above 90% loss (Jordà et al. 2012, 2014). This showcases a dire weak point in carbon conservation rates in methylotrophic yeasts and reinforces the need to look closely at the choice of feed, feeding rate and cultivation conditions for each individual production application of methylotrophic yeasts.
Contrarily, the assimilation pathway of methanol entails the condensation of formaldehyde with Xylulose-5-Phosphate (Xu5P) through the activity of the dihydroxyacetone synthase (DAS) enzyme, generating one molecule of dihydroxyacetone (DHA) and one molecule of glyceraldehyde-3-phosphate (GAP). DHA is further phosphorylated by the dihydroxyacetone kinase (DAK) into dihydroacetone-3-phosphate (DHAP) with the consumption of one molecule of ATP. These C3 molecules constitute the basis for the rest of the reactions in the assimilation pathway, involving a number of rearrangement reactions through enzymes from the pentose phosphate pathway (PPP). Figure 2A summarizes a series of three condensation reactions of formaldehyde through the PPP reactions proposed by Rußmayer et al. (2015). The net gain of these reactions is one C3 molecule per three molecules of formaldehyde, with the concomitant consumption of three molecules of ATP (Fig. 2B). This C3 molecule can then follow a branching path, where it can be further assimilated catabolically through the lower glycolysis, or anabolically through gluconeogenesis or the PPP.
Contrary to the traditional belief that these rearrangements happened cytosolically through the PPP, (Hartner and Glieder 2006; Yurimoto et al. 2011), Rußmayer et al. (2015) showed that methylotrophic yeasts possess a duplication at genome-level for some of the PPP enzymes that carry an additional C-terminal peroxisomal targeting signal (PTS). They further proved these enzymes were enriched in peroxisomal preparations of K. phaffi, thus demonstrating the existence of a compartmentalized cyclic XuMP pathway within the peroxisome separate from the PPP. While not all enzymes predicted to be involved in the cytosolic PPP model have a peroxisomal isoform, other enzymes with similar activities are proposed to compensate for their absence, like could be the case for DAS catalysing other transketolase as the enzyme has been shown to work in vitro with other physiological substrates (Bystrykh et al. 1981; Ro et al. 1997).
The MUT pathway sees its first line of regulation through its localization within one subcellular compartment, the peroxisome. Peroxisome deficient mutants (Cregg et al. 1990; Liu et al. 1992) exhibit strong growth defects or a complete inability to grow when cultivated on methanol as the carbon source, even though they express functionally active versions of the enzymes required for methanol metabolism (Tan et al. 1995; van der Klei et al. 1991). Furthermore, peroxisome biogenesis is induced in the presence of methanol, as demonstrated by their high occurrence in electron-transmission microscopy images of methylotrophic yeast cells grown on methanol (van der Klei et al. 2006; Veenhuis et al. 1983). Transcriptional studies made on O. polymorpha (van Zutphen et al. 2010) and K. phaffi (Rußmayer et al. 2015) also show that peroxisome related genes (PEX) are upregulated in cells grown on methanol. These genes encode peroxins which are the proteins responsible for the biosynthesis, proliferation and function of peroxisomes. Interestingly, this upregulation of PEX genes was functionally differential, with only the genes and proteins related to the methanol assimilation pathway showing an increased expression when compared to PEX genes that carry other metabolic functions like β-oxidation of fatty acids (Prielhofer et al. 2015). Exemplary, PEX5, receptor of the PTS1 signal, and PTS1-harboring genes have been associated with methanol metabolism and showed to be enriched in methanol-grown yeast cells, in contrast to PEX7, the counterpart receptor for the PTS2 signal peptide associated with fatty acid metabolism (Liang et al. 2012; van Zutphen et al. 2010). Along with the activation of methanol metabolism, the mechanisms to mitigate oxidative stress are also transcriptionally regulated upon ROS and aldehyde production through Yap1p, which upregulates the expression of CAT, Pmp20, and the enzymes part of the glutathione recovery system (Rodrigues-Pousada et al. 2019; Yano et al. 2009).
The choice of carbon source also modulates the MUT pathway. When grown on glucose, the peroxisomal enzymes are significantly downregulated, whereas growth on methanol exerts a strong induction of the pathway (Yurimoto et al. 2011). These effects have been observed in a transcriptomic level for O. polymorpha (van Zutphen et al. 2010) and in a transcriptomic and proteomic level for K. phaffi (Liang et al. 2012; Prielhofer et al. 2015; Rußmayer et al. 2015). Enzyme levels for AOX and DAS are elevated on methanol-grown cells, but DAS exhibits a faster expression after induction (Sakai et al. 1996), a mechanism thought to be evolved to reduce the toxicity of formaldehyde by enabling its condensation immediately upon production after the start of the expression of AOX (Yurimoto 2009). Beside methanol induction, these genes also experience different expression levels on different carbon sources, with varying degrees of activity depending on the species of yeast (Yurimoto et al. 2011). The change between glucose and methanol as carbon source has been extensively studied and described (Kang and Gellissen 2004; Hartner and Glieder 2006; Kunze et al. 2009; Yurimoto et al. 2000; Yurimoto and Sakai 2019). Methanol genes are repressed by glucose even at very low concentrations, a mechanism attributed to the evolutionary advantage of utilizing more efficient carbon sources (Dusny and Schmid 2016). Upon glucose depletion or exposure to non-fermentable substrates like glycerol, the genes for methanol assimilation experience variable expression levels depending on the species: C. boidinii and O. polymorpha show around 10% and 80% of the maximum AOX activity, respectively, whereas K. phaffi does not exhibit derepression of the genes without methanol exposure (Kunze et al. 2009; Yurimoto et al. 2011). The mechanism for the methanol-specific induction has been described to be mediated through species-specific transcription factors: Trm1/2p and Hap complex in C. boidinii (Oda et al. 2015, 2016; Sasano et al. 2008, 2010), Mpp1p in O. polymorpha (Leão-Helder et al. 2003; Wang et al. 2016), Mxr1 and Mit1 in K. phaffi (Gupta et al. 2021; Lin-Cereghino et al. 2006; Wang et al. 2016). These transcription factors mediate the induction by methanol at two levels: first at a derepression level (Trm2/Mxr1) upon glucose depletion and further at an actual induction level by methanol (Trm1/Mpp1/Hap) (Yurimoto and Sakai 2019).
Frontiers of metabolic engineering in Methylotrophic yeasts for non-proteinogenic products
Traditional chemical routes to produce bulk and fine chemicals are mostly cost-effective and reliable. However, increasingly stringent regulations on GHG emissions, coupled with concerns over pollution, the depletion of fossil resources, and market volatility, are driving the search for alternatives. In this context, microbial cell factories are promising platforms for contributing to a shift from a linear to a circular economy (Cho et al. 2022). Although microbial systems currently face challenges in terms of economic competitiveness compared to chemical synthesis, they offer several advantages, including safety, scalability, and well-established methodologies (Jozala et al. 2016). A crucial limitation in the advancement of microbial biomanufacturing lies in the availability and cost of feedstocks. This has driven a growing interest in microbial hosts capable of utilising unconventional, low-cost, and sustainable substrates, such as third-generation derived ones, i.e., green methanol.
Methylotrophic yeasts, particularly the genera Ogataea, Komagataella, Kuraishia, and Candida, have gained attention for their unique metabolism and industrially favourable traits. These organisms can grow on methanol as their sole carbon and energy source. In addition, they possess tolerance to low pH and a Crabtree-negative phenotype, which prevents the accumulation of toxic by-products, such as ethanol (Hagman et al. 2014). Additionally, they display the ability to perform correct and compatible post-translational modification, e.g., protein folding, glycosylation, disulphide-bond formation, and an efficient secretion system (Cereghino and Cregg 2000; Damasceno et al. 2012; Karbalaei et al. 2020). Lastly, they possess a tight gene regulation and strong induction of methanol-inducible promoters.
Protein production in methylotrophic yeasts is reviewed elsewhere (Tsuda and Nonaka 2024; Stasyk and Stasyk 2025). In this review, we focus on non-proteinogenic products and the metabolic engineering advances in Ogataea and Komagataella, while Kuraishia and Candida are not debated here as the research on them is still at early stages.
Ogataea
The recently classified genus Ogataea, formerly known as Hansenula, comprises many species known for their role in fermentation and the production of chemicals (Yamada et al. 1994). This success is mainly due to its high tolerance towards high temperatures (growth described up to 50 °C), acidic pH and osmotic stress. Some of the strains of this genus have been fully sequenced, e.g., Ogataea polymorpha NCYC495 and DL-1, and their genomes have been assembled into chromosomes. Nevertheless, the whole genome sequence is available as contings only and is not fully annotated (Chang et al. 2022). Promoters, terminators, and plasmids have been widely described and characterised. In O. polymorpha, promoters like the methanol oxidase promoter (pMOX), the catalase promoter (pCAT) and the dihydroxyacetone synthase promoter (pDHAS) showcase the highest activities with methanol as a C-source (Fig. 2A) (Zhai et al. 2021). Terminators showed an impact on gene expression, affecting mRNA stability but independently of the C-source (Wefelmeier et al. 2022).
Until recently, genetic and metabolic engineering by precise genome modifications in Ogataea was laborious, consisting in flanking gene markers with 500/1000 bp long DNA fragments and homologous recombination (HR) in the genome (González et al. 1999). The non-homologous end-joining (NHEJ) mechanism is the preferred DNA-repair route in Ogataea, which leads to random genomic integration and represents a bottleneck for targeted genetic mutations. To overcome the latter, gene YKU80, responsible for the NHEJ activity involved in heterodimer formation, was knocked out, increasing HR efficiency (Prasitchoke et al. 2007; Saraya et al. 2012). Recently, genetic engineering possibilities expanded thanks to the establishment of CRISPR-Cas9 systems (Numamoto et al. 2017; Wang et al. 2018; Gao et al. 2021; Juergens et al. 2018).
Many molecules, mainly platform and fine chemicals, have been produced in Ogataea using methanol as the sole carbon and energy source (Manfrão-Netto et al. 2019). L-Malic acid production was achieved via expression of reductive tricarboxylic acid (rTCA) cycle in the cytosol of O. polymorpha NCYC495 leu1.1 ∆yku80 by integration of three codon-optimised genes under the control of methanol-inducible promoters (Riley et al. 2016). In detail, the pyruvate carboxylase (PYC) under the control of pMOX, the malate dehydrogenase (MDH) controlled by pCAT, both from Rhizopus oryzae, and the malic acid transporter (MAE1) from Schizosaccharomyces pombe under control of pDHAS. Malate titre reached up to 14 g/L on minimal medium with 1% (v/v) methanol pulses every 24 h (Wefelmeier et al. 2024).
L-lactate production was achieved via metabolic engineering and Adaptive Laboratory Evolution (ALE). Production of L-lactate was achieved in O. polymorpha NCYC495 leu1.1 via genome integration of the codon-optimised L-lactate dehydrogenase (LDH) from Lactobacillus helveticus in the HIS2 genomic locus and expression under the control of the pMOX promoter and tMOX terminator. The obtained titre was 3.0 g/L, which was further improved to 3.8 g/L through ALE of the same strain by daily 0.5% v/v pulses (Wefelmeier et al. 2023).
Acetone was produced at a titre of 8.6 mg/L. The genetic engineering approach has been the same as described for malate and lactate. The codon-optimised sequences coding for three enzymes from Clostridium acetobutylicum were integrated into the O. polymorpha genome under the regulation of methanol-inducible promoters pMOX and pCAT. In order, these are the acetyl-CoA acetyltransferase, the butyrate-acetoacetate CoA-transferase and the acetoacetate decarboxylase. The engineered yeast was supplied with 0.5% (v/v) methanol as the carbon source, and acetate was added at low concentrations to the cultivation to improve production (Wefelmeier et al. 2024).
Isoprene production is another example of an achievable metabolic engineering possibility in Ogataea. The gene coding for isoprene synthase Isps(p) from the plant Populus alba and the truncated version of the 3-hydroxy-3-methylglutaryl-CoA reductase gene from Saccharomyces cerevisiae (N-terminal signal) were integrated into the O. polymorpha genome under control of pDHAS and pMOX, respectively. The highest titre reported is 4.4 g/L, feeding 0.5% (v/v) methanol (Wefelmeier et al. 2024).
The production of another major platform chemical, i.e., hydroxypropionate (HP), was also achieved in O. polymorpha. Biologically, it is synthesised from the cellular central intermediate malonyl-CoA. A mutated version (N940V, K1106W and S1114R) of malonyl‐CoA reductase (MCR) from Chloroflexus aurantiacus with improved catalytic efficiency for 3‐HP production and under the control of the methanol-induced promoter pAOX was introduced in O. polymorpha genome. Together with precursor and cofactor enhancement and a fed a titre of 7.10 g/L was achieved (Liu et al. 2016; Yu et al. 2023).
Largely used in the detergent and cosmetic industries, fatty alcohols were produced by Zhai et al. (2023) from methanol via a multisided approach and reached a titre of 3.6 g/L. First, the fatty alcohols synthetic pathway was segregated into the peroxisomes by expressing the fatty acyl-CoA reductase (TaFAR1) from Tyto alba, commonly known as barn owl, and the alcohol dehydrogenase (ScADH5) from S. cerevisiae. Additionally, O. polymorpha ZX-F75U was genetically engineered sequentially to optimise the supply of peroxisomal fatty acyl-CoA and NADPH by (i) overexpressing in the peroxisomes the endogenous PYC1, MDH3 and the heterologous ME1 from Rhodosporidium toruloides, (ii) enhance cell robustness by deleting a putative lipase (LPL1) and a membrane protein related to zinc metabolism (IZH3), and (iii) enhance methanol tolerance by overexpressing the dihydroxyacetone synthase gene DAS2 (Zhai et al. 2023).
With its complex 15-carbon cycle structure molecule, its high cetane number and sustainable nature, β-farnesene is a peculiar specialty chemical produced in O. polymorpha. Its production from methanol was achieved with extensive metabolic engineering in the fatty acid overproducing strain M16. Specifically, the geranyl/farnesyl diphosphate synthase-encoding gene (ERG20) was fused with the β-farnesene synthase-encoding gene (FS) from the plant Artemisia annua and expressed under the control of the methanol-inducible promoter pTAL1. Native truncated 3-hydroxy-3-methylglutaryl coenzyme A reductase (tHMGR) and HMGR from Silicibacter pomeroyi, fused ERG10-ERG13,* ERG12*,* ERG8*, and ERG19 were overexpressed. Last, the β-oxidation pathway was systematically overexpressed, and the cytosolic malate synthase (MLS1) was knocked out (Li et al. 2024).
Komagataella
The methylotrophic yeast belonging to the Komagataella genus, classified until 1995 as Pichia pastoris (Yamada et al. 1995) and renamed today as Komagataella phaffii, is a well-established platform for recombinant protein production (Wu et al. 2023). The K. phaffii genome has been fully sequenced and annotated (De Schutter et al. 2009), and its success in research and industry is due to its ability to grow to high-cell densities on minimal media, being Crabtree negative (Cereghino and Cregg 2000) and displaying high-production yields. Notably, genetic tools for K. phaffii have been widely described and developed. In 1985, the gene coding for alcohol oxidase (AOX1), the enzyme involved in the first enzymatic step of the methanol oxidation pathway, and its promoter (pAOX1) were characterised (Cregg et al. 1985; Ellis et al. 1985). Afterwards, expression vectors, integrative vectors, and selection markers were described (Cereghino and Cregg 2000). Like in other methylotrophic yeasts, the recombination machinery in K. phaffii is less efficient than in S. cerevisiae. Therefore, a functional CRISPR-Cas9 system for K. phaffii was recently developed (Weninger et al. 2016; Gao et al. 2022; Wu et al. 2023).
Based on old and new technologies, significant progress has been made to expand the product array in K. phaffii, using methanol as the sole carbon and energy source. Guo et al. (2021) reported malate production using a multisided approach. First, a methanol-inducible rTCA pathway from Rhizopus oryzae was expressed. Genomic integration was achieved via integrative plasmids bearing heterologous genes, pyruvate carboxylase (PYC) and malate dehydrogenase (MDH) under the control of the pAOX1 promoter. To overcome a common bottleneck in malic acid production, the malate transporter (MAE) from S. pombe was integrated into the genome and overexpressed under the control of pAOX1. In addition, to redirect metabolic fluxes from the XuMP cycle (Fig. 2A), glucose-6-phosphate isomerase was deleted. Lastly, the cultivation medium was supplemented with 1 g/L yeast extract as an additional nitrogen source. This multisided approach produced 2.79 g/L of malic acid from methanol (Guo et al. 2021a).
Another organic acid of industrial interest, D-lactic acid, was produced from methanol at a final titre of 3.48 g/L. K. phaffii does not synthesise lactate natively. Therefore, the gene coding for lactate dehydrogenase (LDH) from Leuconostoc mesenteroides was integrated into the genome, under the control of native pAOX1 and tAOX1 terminator. The integration was achieved using a multi-copy integrative vector targeting the non-transcribed spacer (NTS) of the ribosomal DNA (rDNA) locus, yielding a high gene copy number (Yamada et al. 2019). Recently, Bachleitner et al. (2024) produced 17 g/L of lactate from methanol in a bioreactor, using an engineered K. phaffii strain. This was modified by (a) integration in the RGI2 locus of the codon-optimised L-lactate dehydrogenase from Lactiplantibacillus plantarum under the control of pAOX1 and (b) deletion of the L-lactate cytochrome-c oxidoreductase CYB2 using the CRISPR-Cas9 system. Even though this integration was enough to produce lactate, it was only via overexpression of transcriptional activators MXR1 and MIT1 that the titre increased significantly (Bachleitner et al. 2024). To produce itaconic acid, Severinsen et al. (2024) engineered Komagataella phaffii via integration of the heterologous genes from Aspergillus terreus for the itaconate metabolic pathway. Specifically, the gene responsible for the conversion of cis-aconitate to itaconate, cadA, was integrated into the yeast genome under the control of pAOX1. The transporters’ genes mttA and mfsA were integrated into the genome under the control of a moderately strong and strong constitutive promoter, respectively. Further process optimisation, i.e., increase of temperature to 32 °C and adoption of a multiphase process, in a bioreactor led to titres of 55 g/L after circa 5 days of methanol feed.
Another interesting example showcasing the possibilities of using methanol as a fermentation substrate is β-alanine, a non-essential amino acid, and the only natural β-amino acid. This molecule is a precursor of many nitrogen-based compounds, but its industrial synthesis requires harsh, not environmentally friendly conditions. In K. phaffii, 5.6 g/L of β-alanine was produced by introducing copies of the L-aspartate-α-decarboxylase (ADC) from Bacillus subtilis and the aspartate dehydrogenase (AspDH) from Serratia proteamaculans encoding genes in its genome, using integration vectors. Both genes were placed under the control of the methanol-inducible promoter pAOX1 (Miao et al. 2021). Two different groups were able to produce 3-hydroxypropionate (3-HP) from methanol in K. phaffii (Àvila-Cabré et al. 2023; Chen et al. 2025).
Two different groups independently achieved a notable 3-HP titre in this yeast. In detail, Chen et al. (2025) were able to produce 23 g/L of 3-HP following the heterologous expression of two genes from the β-alanine synthetic pathway, AspDH from Brucella anthropic and ADC from Bacillus subtilis, in one and three copies respectively integrated into the genome. Moreover, to optimise the metabolic flux of pyruvate and avoid the formation of acetic acid, the aldehyde dehydrogenase gene (ALD4) was knocked-out. Lastly, the final titre was additionally increased after the integration of the expression cassette constructed fusing the β-alanine pyruvate aminotransferase gene (BAPAT) from Bacillus cereus under the pAOX promoter and the malonate semialdehyde reductase gene (MCRN) under the PS2 promoter. On the other hand, Àvila-Cabré et al. (2023) were able to reach a titre of 21.4 g/L of 3-HP. Here, the β-alanine pathway was constructed via integration into K. phaffii genome of two copies of panD under the control of pAOX1-from Tribolium castaneum (2x), BAPAT from Bacillus cereus under the control of pFDH1, and ydfG from Escherichia coli K-12 under the moderately strong promoter pPOR.
Fatty alcohols are part of the plethora of chemicals produced via genetic engineering of K. phaffii. Shen et al. (2024) engineered and optimised the cytosolic fatty alcohol biosynthesis in the yeast by integrating into its genome the alcohol dehydrogenase gene (ScADH5) from S. cerevisiae under the control of pTEF1 and knocking-out the aldehyde dehydrogenase gene (hfd1). Then the codon optimized fatty acyl-CoA reductases (FAR) from Mus musculus was selected and heterologously expressed under the control of pTEF1 promoter and targeted to the peroxisome via a signal peptide. In this strain, the methanol assimilation pathway was also strengthened, by overexpressing DAS2 under its natural promoter, with good results. Noteworthy, is also the intervention, i.e., overexpression under the control of pPGI1 of PEX8, on the peroxisomes, aiming at coupling methanol assimilation and fatty alcohols production. The highest titre was only obtained when the cytoplasmic and peroxisomal engineering were combined in the diploid K. phaffii SYW15 strain, reaching a titre of 5.6 g/L (Shen et al. 2024). Several other bulk, fine chemicals and pharmaceuticals were synthesised in K. phaffii using methanol as the sole carbon source, although in low amounts, and are listed in Table 2.
Table 2. Alphabetical list of bulk-, fine-, and specialty chemicals produced from methanol in the two most prominent methylotrophic yeast, Ogataea and KomagataellaMicroorganismProductCategoryTitreGenetic engineering strategyYieldgP/gMeOHProductivityReferences Ogataea AcetoneBulk8.6 mg/LHeterologous expression under methanol-inducible promoters of pCAT-THLA from C. acetobutylicum, pMOX-YbgC from H. influenzae, and pMOX-Adc from P. polymyxan.q.n.q. Yu et al. (2023), Wefelmeier et al. (2023), Wefelmeier et al. (2024), Zhai et al. (2023), Li et al. (2024)Fatty alcoholsBulk3.6 g/LHeterologous expression of TaFAR1 from T. alba and ADH5 from S. cerevisiae. Overexpression of IDP2 from S. cerevisiae and deletion of LPL1, IZH3. Overexpression of endogenous PYC1 and MDH3, and RtME1. Overexpression of DAS20.01612.9 mg/L/hHydroxypropionateFine7.10 g/LHeterologous expression under methanol-inducible promoter: pAOX-MCR from C. aurantiacusn.q.n.q.IsopreneBulk4.4 g/LHeterologous expression under methanol-inducible promoters: pDHAS-Isps(p) from P. alba and pMOX-hydroxy-3-methylglutaryl-CoA reductase from S. cerevisiaen.q.n.q.L-Lactic acidBulk3.8 g/LHeterologous expression under methanol-inducible promoter and terminator: pMOX-LDH-tMOX from L. helveticus, and ALE0.0212.6 mg/L/hMalic acidFine13 g/LHeterologous rTCA expression under methanol-inducible promoters: pMOX-PYC, pCAT-MDH, both from R. oryzae, pDHAS-MAE1 from S. pombae0.413.3 g/L/dayβ-farneseneSpecialty14.7 g/LHeterologous expression of pTAL1-ERG20 fused with FS from A. annua and overexpression of tHMGR and HMGR from S. pomeroyi, fused ERG10 and ERG13 (2x), ERG12, ERG8 and ERG19. Overexpression of β-oxidation pathway and knock out of MLS10.04648 mg/L/h Komagataella AstaxanthinSpecialty716.13 mg/LHeterologous expression of CrtE, CrtYB, CrtI, CrtYB, CrtZ, CrtWn.q.n.q.Yamada et al. (2019), Miao et al. (2021), Guo et al. (2021), Gao et al. (2022), Bachleitner et al. (2024), Severinsen et al. (2024), Shen et al. (2024), Chen et al. (2025), Àvila-Cabré et al. (2023), Wang et al. (2025)Chondroitin sulphateSpecialty2.1 g/LHeterologous expression of KFOA and KFOC from E. coli expressing TUAD from B. subtilis. Overexpression of ATPS and APSK.n.q.n.q.CitrininSpecialty0.6 mg/LHeterologous expression of PKSCT, MPL1, MPL2, and MPL4 from M. purpureus, MPL6 and MPL7 from M. ruber, and NPGA from A. nidulansn.q.n.q.D-Lactic acidBulk3.48 g/LHeterologous expression of pAOX1-LDH-tAOX1 from L. mesenteroides0.2236.3 ± 2.0 mg/L/hFatty alcoholsBulk5.6 g/LCombination of (i) heterologous expression of pTEF1-ADH5 from S. cerevisiae and pTEF1-FAR from M. musculus, (ii) overexpression of pDAS-DAS2 and pPGI1-PEX8, (iii) knock-out of hfd1 in K. phaffii SYW15.0.13n.q.3-HydroxypropionateBulk23 g/L21.4 g/LHeterologous expression of AspDH from B. anthropic and ADC from B. subtilis (x3). ALD4 deletion; pAOX-BAPAT–PS_2_-McrN fusion cassette expression with both genes from B. cereusHeterologous expression of pPOR1-panD from T. castaneum (2x), yhxA from B. cereus, and ydfG from E. coli K-120.120.15n.q.0.48 g/L/hHyaluronic acidSpecialty1.7 g/LHeterologous expression of xhasa2 and xhasb from X. laevis. Overexpression of HASC, HASD, and HASEn.q.n.q.Itaconic acidBulk55 g/LHeterologous expression and overexpression of pAOX1-cadA, pPOR1-mttA and pGAP-mfsA from A. terreus0.240.45 g/L/h L-Lactic acidBulk17 g/LHeterologous expression of pAOX1-LDH from L. plantarus; deletion of Nrg1, Mig1-1, and Mig1-2; overexpression of transcriptional activators Mxr1 and Mit1n.q.n.q.LovastatinSpecialty250.8 mg/LHeterologous expression of LovB, LovC, and LovG from A. terreus and NpgA from A. nidulansn.q.n.q.LycopeneSpecialty714 mg/LHeterologous expression of CrtB, CrtI, and CrtE from C. glutamicum, overexpression of hmgr, and hmgs. Increase of ggpps copy numbern.q.n.q.Malic acidFine2.79 g/LHeterologous rTCA expression under methanol-inducible promoters: PYC, MDH both from R. oryzae, MAE1 from S. pombae. Deletion of Ku70, ODC1p, PDC, GPIn.q.n.q.Monacolin JSpecialty593.9 mg/LExpression of LovA and split of CPR pathwayn.q.n.q.ZeaxanthinSpecialtyn.q.Heterologous expression of CrtI, CrtE-CrtZ, CrtYBn.q.n.q.2,3-butanediolBulk2.2 g/LHeterologous expression of AlsS, AlsD, BDH,n.q.n.q.6-methyl salicylic acidFine2.2 g/LHeterologous coexpression of NpgA from A. nidulans and atX from A. terrusn.q.n.q.α-bisaboleneSpecialty270 mg/LHeterologous expression of pTEF1-tHMGR and pGCW14-BIS fused with FPPS from A. grandis (multiple copies). Overexpression of ERG10 and HMGS under pTEF1. Overexpression of MK and PMK.0.012.25 mg/L/hβ-alanineFine5.6 g/LHeterologous expression of PAOX1-ADC, SpeAspDH (overexpressed) from B. subtilis, deletion of Ku70n.q.n.q.β-caroteneSpecialtyn.q.Heterologous expression of CrtE, CrtYB, CrtI, CrtYB, CrtZn.q.n.q.β-farneseneSpecialty338 mg/LHeterologous expression of pGCW14-FS from A. annua fused with FPPS0.022.8 mg mg/L/hTitre, genetic engineering strategy, yield in grams of product/grams of methanol and productivity are listed; (n.q. = non-quantified)
Process considerations in methanol fermentation
Compared to other C1 feedstocks, e.g., methane, the nature and characteristics of methanol offer several benefits as a substrate in industrial processes. Methanol is a colourless, odourless, and water-miscible liquid. High water solubility is especially crucial to avoid gradients in nutrient concentration and mass transfer limitations that could otherwise impair fermentation efficiency (Crater and Lievense 2018). Noteworthy is methanol’s flexibility as a feedstock: deriving from fossil or renewable sources, it allows transitioning from one to the other as the technology develops and market prices or policies shift. Lastly, its transport, manipulation, and storage are convenient, being a liquid at room temperature (Olah 2005). Nevertheless, handling methanol needs careful consideration. The molecule is toxic and possesses high volatility and flammability. A major concern is the risk of explosion due to methanol vapour accumulation, which can easily occur in industrial settings. For these reasons, industrial facilities handling methanol must comply with regulations that vary by country. Typically, large-scale fermentation facilities must have explosion-proof equipment, safe storage, constant and continuous monitoring of methanol concentration, and proper ventilation (Antoniewicz 2019; Gan et al. 2023), increasing capital expenditure (CAPEX).
The remarkable flexibility and benefits of methanol-based processes also derive from the microorganisms employed. Naturally, microbial fermentations and bioconversions do not require extreme working conditions, e.g., high pressure, high temperature, harsh chemicals, or catalysts. This makes them not only sustainable but also cost-effective and safer. Additionally, the availability of methanol-inducible promoters in methylotrophic yeasts, such as the alcohol oxidase gene or the catalase gene promoters, allows precise gene transcription activation and, hence, efficient fine-tuning of metabolic pathway activation (Yan et al. 2022). In addition, they possess tight regulation and noteworthy strength (Cregg et al. 1993; Wu et al. 2023). Consequently, different fermentation approaches have been developed and optimised; indeed, multiple-phase fermentation is the most suitable when using methanol as the induction substrate for methanol-induced product production (Gan et al. 2023). First, it allows the decoupling of the growth and production phases. Since both are highly energetically and metabolically demanding, this increases final biomass density and production efficiency. Secondly, it concedes precise induction of genes controlled by methanol-induced promoters, preventing intermediates toxicity of methanol metabolism, i.e. formaldehyde, and generating higher final titres and yields (Ahmad et al. 2014; Tsuda and Nonaka 2024).
In this context, a typical multi-phase process (Fig. 3) begins with the first phase (S1), designed as a batch phase to grow biomass with either glucose or glycerol, respectively. Once the set amount of biomass is achieved, the second phase (S2) focuses on production, i.e., a fed-batch phase with methanol as source of carbon and energy is initiated (Brierley 1998; Potvin et al. 2012). A three-phase process has also been developed, where the S1 and S3 phase follow the previously described approach but a new intermediate phase (S2) is introduced: a fed-batch phase, which uses glycerol to further increase cell density, like in S1 (Liu et al. 2016; Potvin et al. 2012; Severinsen et al. 2024; Totaro et al. 2021). In the latter S2, the feeding rate can be either constant or follow an exponential feeding profile (Potvin et al. 2012). To have a smoother transition from S2 and S3, methanol and glycerol can be co-fed during an additional phase for a given length of time, during which glycerol concentration in the feed gradually decreases, while the concentration of methanol increases accordingly (Cereghino and Cregg 2000; Zhang et al. 2000).
Graphical description of a generic fermentation process using a methylotrophic yeast. Aseed fermentation on complex medium is followed by a batch and fed-batch phase whereglucose, glycerol and methanol are the main feedstocks and the production of the targetcompound is triggered by methanol addition
In the Ogataea genus, glycerol can derepress the methanol oxidase promoter up to 80% compared to the activation on methanol only. Hence, the transition phase can have a meaningful impact on the final production in Ogataea. Lastly, the fed-batch phase (S3) using methanol can be further optimised by tuning it on the microorganism’s methanol uptake rate and the goal of the process. This can be done by adopting different feeding strategies: constant Dissolved Oxygen feeding (DO-stat), constant specific growth rate feeding, constant methanol concentration feeding, oxygen-limited fed-batch (OLFB), and temperature-limited fed-batch (TLFB) (Potvin et al. 2012). During the entire duration of the process, methanol level monitoring is essential. The substrate concentration must always be kept within a well-defined range, typically between 0.5% v/v and 2.5% v/v. In fact, too high concentrations will hinder growth and toxify the cultivation broth. This is mainly due to formaldehyde accumulation which will lead to cell death; on the other hand, too low concentrations can slow down the production (Wang et al. 2023). Despite these process-control-intensive constraints, using methanol ensures a minimal risk of contamination, an amenable condition in industrial settings where maintaining sterility is challenging (Cregg et al. 2000).
An important economical consideration, particularly in large-scale processes, is the oxygen transfer efficiency (OTE) and the heat release associated with methanol metabolism (Singh et al. 2022). Methanol fermentation, as a highly oxidative process, is oxygen-demanding due to the initial reaction in methylotrophic yeast metabolism and the necessity to re-oxidise the NADH pool generated by the assimilation and product pathways (Fig. 2A). At the metabolic level, pathways with higher carbon efficiency, i.e., those directing a greater proportion of methanol carbon into product rather than CO₂, also exhibit markedly lower O₂ demand, since less substrate oxidation is needed for redox balancing (Wagner et al. 2023). This relationship is particularly relevant in high-cell-density fermentations (HCDF), where insufficient OTE leads to methanol accumulation and incomplete substrate-to-product conversion, reducing final titres and yields in oxygen-limited conditions (Liu et al. 2016; Mate et al. 2013; Shen et al. 2022). To compensate, pure oxygen supplementation is required during the process, which significantly raises Operating Expenditure. Alternatively, process intensification strategies such as super-atmospheric aeration can reduce oxygen-related costs by 20–30% across multiple production cycles by eliminating the need for pure oxygen (Campani et al. 2016). Liu et al. (2016) demonstrated an effective approach to overcome oxygenation limitation in a HCDF without the need of pure oxygen addition. The group increased the reactor air pressure from 0.05 MPa (super atmospheric pressure) to 0.10 ± 0.05 MPa to enhance gas solubility. Although specialised equipment was required, the feasibility of the removal of a pure oxygen supplement was successfully demonstrated increasing the volumetric production by 43% and reducing costs (Charoenrat et al. 2006; Liu et al. 2016).
Conclusion and outlook
In this review, we showcased how methylotrophic yeasts have emerged as promising microbial platforms for converting methanol into value-added specialty and fine chemicals. Within the framework of a circular economy, methanol as a substrate is particularly attractive: it can be derived sustainably from 3G feedstocks, i.e., captured CO₂ and renewable energy; and its use in biotechnology will benefit from existing demands of the chemical industry. Moreover, industrial infrastructure for methanol-based chemical production is in place and at scale, thereby minimising CAPEX and facilitating a smooth transition from fossil-based to renewable sources. Hence, “land-free” methanol offers an advantageous “drop-in” solution that reduces investment efforts during the shift to more sustainable feedstocks for chemical production processes.
To favour and fulfil the ambitious goal of reshaping industrial production and achieve the predicted bioeconomy scenario, intensive technology development and maturity are especially urgent. In fact, the biotechnological conversion of methanol via yeast fermentation has still some major limitations that hinder economic feasibility and attractiveness. This is especially accurate for bulk chemicals production, that are usually produced via well-stablished and efficient chemical processes. Chemical catalysis can achieve carbon conversion efficiencies around and above 90% and volumetric productivities of tons of product per m³ per day. In contrast, bioconversion requires a certain degree of complexity and generally exhibits conversion efficiencies rarely above 60%, along with low volumetric productivities. Considering methanol high degree of reduction, inherently lower conversion yields, influenced by the lower reduction state of the product, are to be expected. Furthermore, the volumetric productivity in a fermentation process is generally lower of several orders of magnitude compared to a catalytic process. Protein production remains the most established and economically viable application. Therefore, bioprocess development should first focus on high-value, low-volume products, i.e., fine and specialty chemicals, that are not easily synthesised via chemical routes. This strategy could accelerate the establishment of C1 feedstocks in the sector, which would boost and expand the technology maturity to then develop processes for other chemicals in a lower price range.
While considerable challenges remain, particularly regarding process economics and strain performance, targeted innovations in strain engineering, promoter design, and upstream feedstock sourcing could position methylotrophic yeasts as competitive and sustainable platforms for the next generation of bio-based manufacturing.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bennett RK, Gregory GJ, Gonzalez JE, et al (2021) Improving the methanol tolerance of an Escherichia coli methylotroph via adaptive laboratory evolution enhances synthetic methanol utilization. Front Microbiol 12:638426. 10.3389/FMICB.2021.638426/BIBTEX
- 2Campani G, Possedente dos Santos M, Gonçalves da Silva G et al (2016) Recombinant protein production by engineered Escherichia coli in a pressurized airlift bioreactor: A techno-economic analysis. Chem Eng Process: Process Intensif 103:63–69. 10.1016/j.cep.2015.10.020
- 3Global hydrogen review (2022) IEA. https://www.iea.org/reports/global-hydrogen-review-2022
- 4Higgins DR, Cregg JM (1998) Introduction to Pichia pastoris. In: Higgins DR, Cregg JM (eds), Pichia Protocols. Humana Press, pp 1–15. 10.1385/0-89603-421-6:1
- 5Horiguchi, H., Yurimoto, H., Kato, N., et al. (2001). Antioxidant system within yeast peroxisome: biochemical and physiological characterization of Cb Pmp 20 in the methylotrophic yeast Candida boidinii. J Biol Chem 276(17):14279–14288. 10.1074/JBC.M 011661200
- 6Juergens H, Varela JA, Gorter de Vries, et al (2018) Genome editing in Kluyveromyces and Ogataea yeasts using a broad-host-range Cas 9/g RNA co-expression plasmid. FEMS Yeast Res 18(3):foy 012. 10.1093/femsyr/foy 012
- 7Kang HA, Gellissen G (2004) Hansenula polymorpha. In: Production of Recombinant Proteins. Wiley, pp 111–142. 10.1002/3527603670.ch 6
- 8Kunze G, Kang HA, Gellissen G (2009) Hansenula polymorpha (Pichia angusta): biology and applications. Yeast biotechnology: diversity and applications, pp 47–64. 10.1007/978-1-4020-8292-4_3