The Plasmodium berghei merozoite protein PbGAC is critically involved in erythrocyte binding during invasion
Junying Gao, Ning Jiang, Yiwei Zhang, Ran Chen, Ying Feng, Xiaoyu Sang, Qijun Chen

TL;DR
This study identifies a protein in the malaria parasite Plasmodium berghei that helps it invade red blood cells by binding to specific receptors.
Contribution
The study reveals the role of PbGAC as a key protein involved in erythrocyte invasion via heparan sulfate-dependent binding.
Findings
PbGAC is located at the apical region of merozoites and binds to heparin and erythrocyte surfaces.
PbGAC interacts with proteins critical for erythrocyte invasion, as shown by mass spectrometry.
Immunization with PbGAC enhances immunity against lethal malaria challenges in mice.
Abstract
The invasion of Plasmodium merozoites into host erythrocytes is initiated through specific ligand–receptor interactions. This interaction results in subsequent invasion events, facilitated by the formation of a moving junction via AMA-1 and associated molecular complexes. Previous studies have implicated erythrocyte surface glycosaminoglycans, particularly heparan sulfate proteoglycans, as critical receptor components in this invasion process. The binding affinity of the PbGAC protein to heparin and erythrocytes was assessed through western blotting, immunofluorescence, flow cytometry techniques, and heparinase II treatment. Mice were immunized with the recombinant PbGAC-His to generate specific polyclonal antibodies for subcellular localization, passive immunization, and immunoprecipitation. Global mass spectrometric analyses were conducted to identify its interacting proteins. We…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
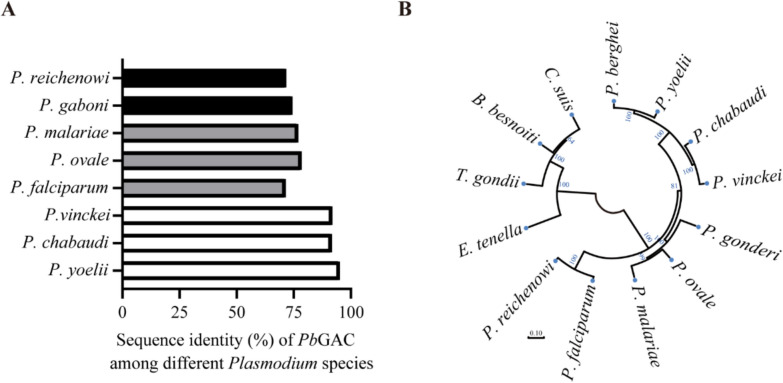 Figure 1
Figure 1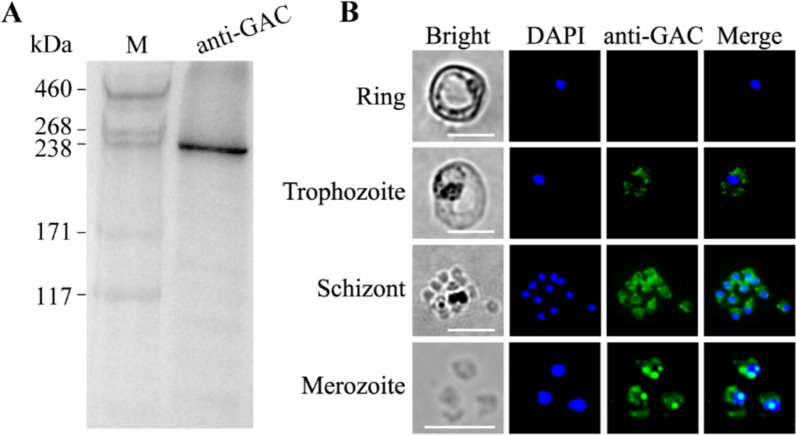 Figure 2
Figure 2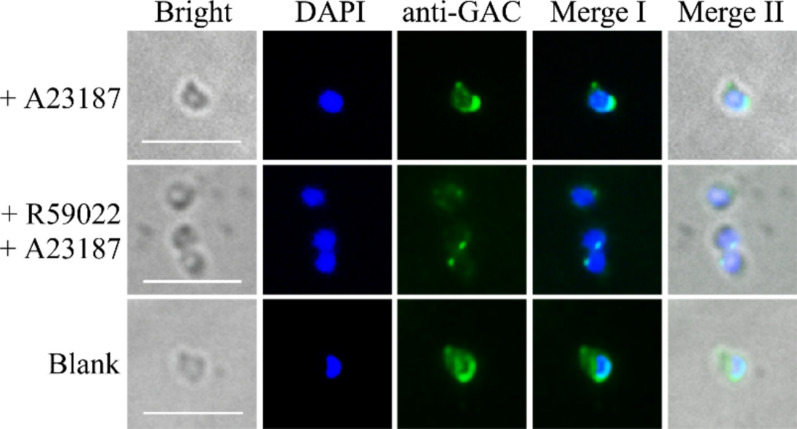 Figure 3
Figure 3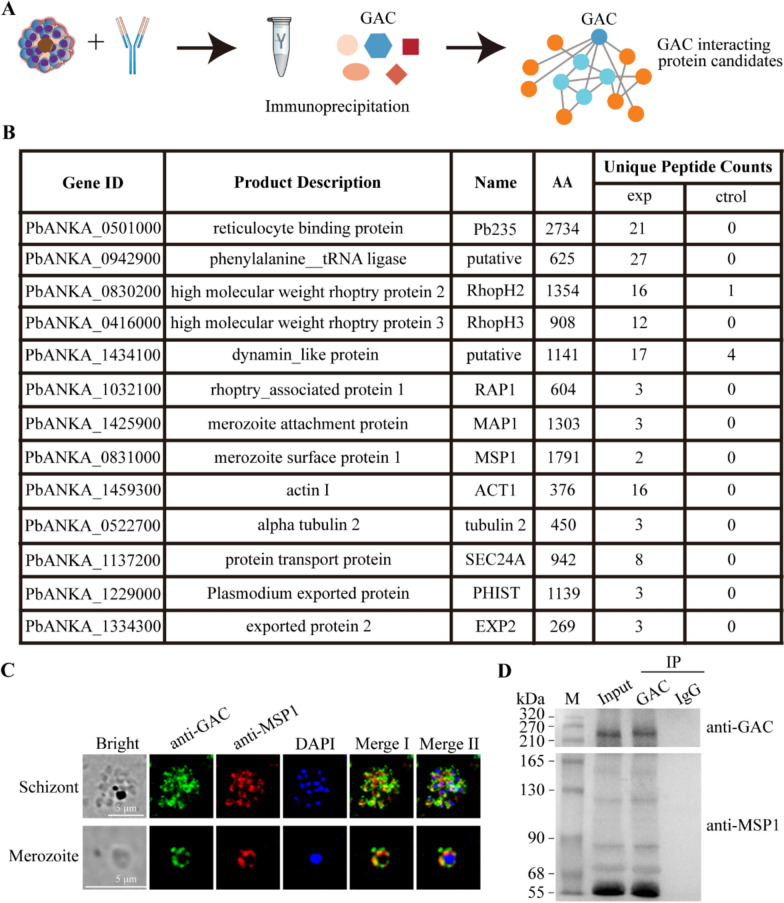 Figure 4
Figure 4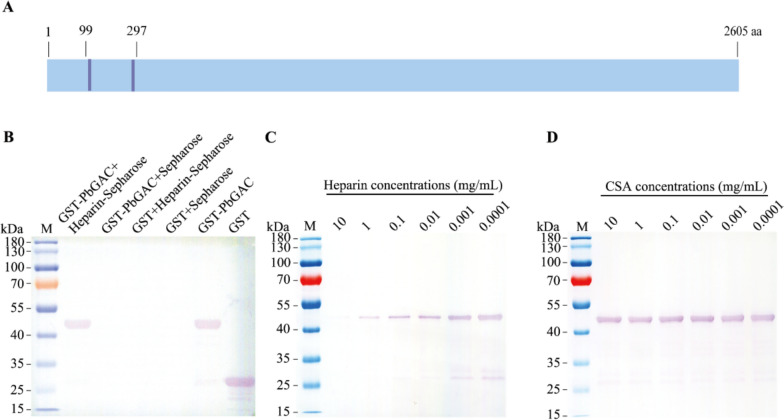 Figure 5
Figure 5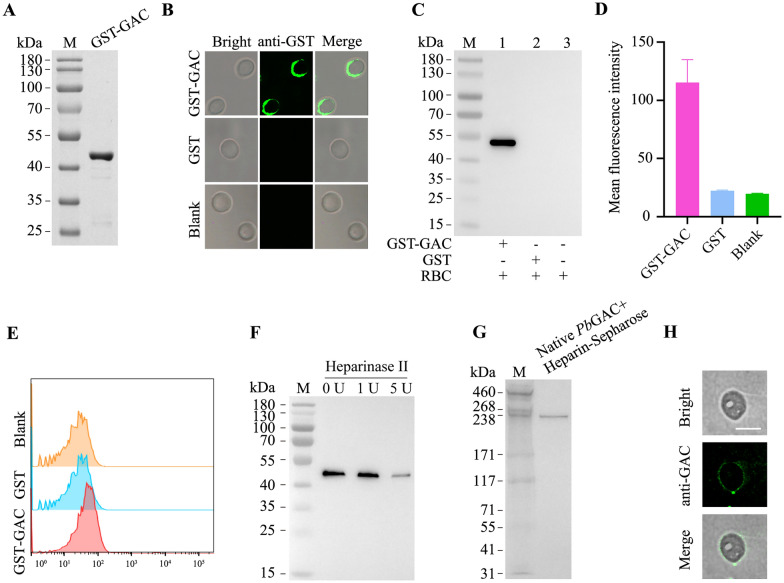 Figure 6
Figure 6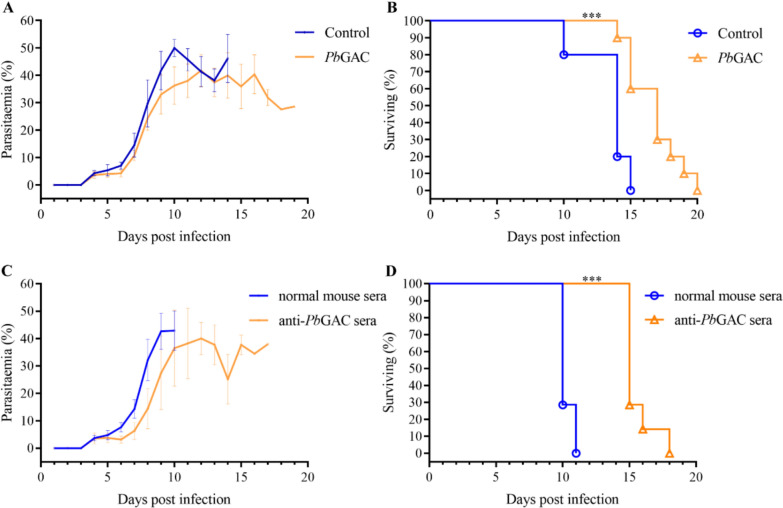 Figure 7
Figure 7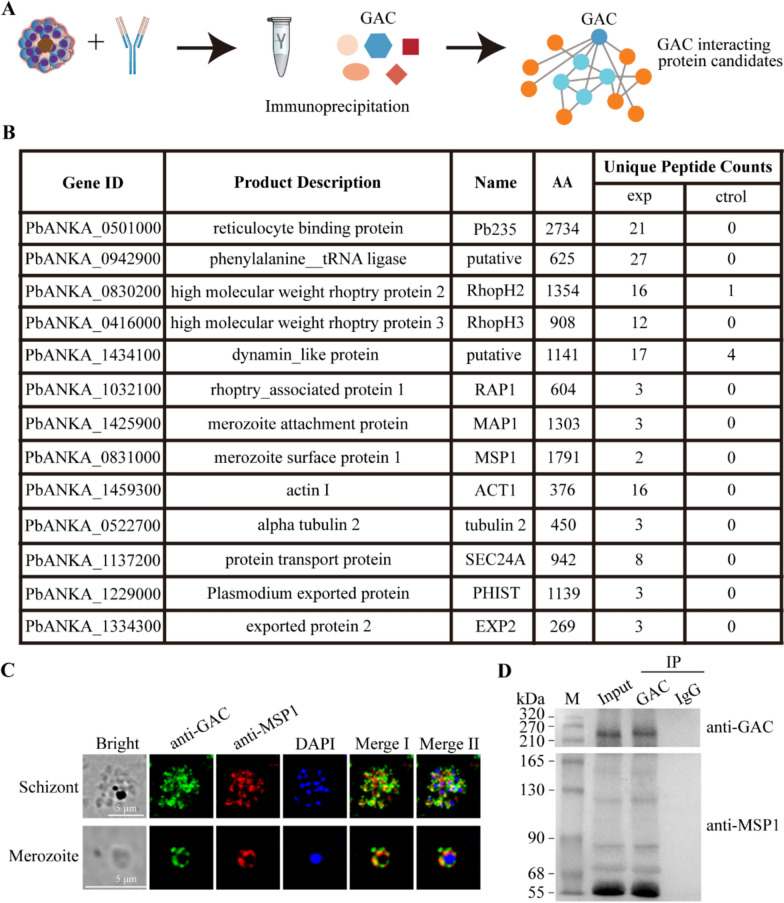 Figure 8
Figure 8- —National Natural Scientific Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Calpain Protease Function and Regulation · Proteoglycans and glycosaminoglycans research
Background
Members of the phylum Apicomplexa constitute a group of obligatory intracellular protozoan parasites responsible for major human parasitic diseases, including malaria and toxoplasmosis. Plasmodium species exhibit a complex biphasic life cycle involving asexual replication in vertebrate hosts and sexual development in mosquito vectors [1]. The pathogenesis of severe malaria is directly correlated with the exponential expansion of asexual blood-stage parasites, where cyclical fever episodes coincide with the synchronized egress of merozoites from lysed erythrocytes [2]. During intraerythrocytic development, invasive merozoites progress through morphologically distinct stages—ring, trophozoite, and schizont—with mature schizonts generating 16–32 daughter merozoites capable of initiating new infection cycles [3]. To bypass the erythrocyte barrier, Plasmodium merozoites utilize a distinctive mechanism for erythrocyte invasion. This process begins with a “long-distance” recognition of surface receptors, which is succeeded by a reorientation phase that sustains these low-affinity interactions. Once the apical end of the merozoite is in proximity to the erythrocyte, a tight junction is established through high-affinity ligand–receptor interactions. Subsequently, this tight junction translocates from the apical to the posterior pole, driven by the actin–myosin motor of the parasite [4]. This erythrocytic invasion process, governed by specialized invasion-associated proteins, represents an essential mechanism for parasite propagation.
The Plasmodium rapid replicative cycle requires coordinated gliding motility to facilitate host-cell entry, egress, and traversal through biological barriers [5–7]. Merozoite invasion of erythrocytes occurs through four mechanistically distinct phases: (1) low-affinity attachment to the erythrocyte membrane, (2) apical reorientation, (3) formation of a tight moving junction, and (4) establishment of the parasitophorous vacuole [8]. At each stage, the process involves the kinetics of merozoite invasion and elucidates the ligand–receptor interactions involved. A series of well-characterized merozoite ligands are sequentially released from merozoite organelles, specifically rhoptries and micronemes, facilitating a tight attachment and reorientation of the merozoite toward its apical end on the erythrocyte surface in preparation for invasion. At the molecular level, these preinvasion events initiate when merozoite surface proteins (MSPs) mediate the initial, low-affinity attachment, which subsequently triggers the expression of erythrocyte binding-like (EBL) and reticulocyte binding-like (RBL) family members as well as apical membrane antigen (AMA-1) on the merozoite surface [9–12]. Initial merozoite–erythrocyte interactions, primarily mediated by glycosylphosphatidylinositol (GPI)-anchored surface proteins, are critical for invasion competency [13]. Key molecular mediators include merozoite surface protein 1 (MSP1), thrombospondin anonymous repeat protein (TRAP), erythrocyte binding-like protein 175 (EBA-175), and reticulocyte-binding protein 4 (PfRh4), which collectively facilitate erythrocyte adhesion [14–18]. The binding of Rh5 to basigin (BSG), a blood group antigen of the Ok blood group system, is an essential binding event, representing the irreversible commitment step for merozoite entry into the erythrocyte and acting as the trigger for all subsequent invasion processes [19]. The Rh5-binding event promotes the formation of a fusion pore between the merozoite rhoptries and the erythrocyte, initiating calcium influx into the erythrocyte and the release of rhoptry contents [20]. Subsequent invasion steps involve proteins derived from apical organelles, such as apical membrane antigen 1 (AMA-1), a type 1 transmembrane protein, which is secreted onto the merozoite surface during egress from the previous host cell. AMA-1 interacts with RON2 to establish the moving junction complex, a critical structure mediating parasite internalization [21–23].
Heparan sulfate (HS), a ubiquitous glycosaminoglycan on vertebrate cell surfaces, serves as a key binding partner for apicomplexan surface antigens and secretory proteins [24–26]. Although HS-binding proteins remain incompletely characterized, current evidence identifies Toxoplasma gondii SAG1, ROP2/4, and GRA2 as invasion-related HS interactors [27]. In Plasmodium, merozoite surface protein PfMSP-1 and merozoite attachment protein PbMAP1 demonstrate HS-binding capacity [18, 24, 28]. Notably, apical organelle-localized proteins also engage specific erythrocyte receptors during invasion [29, 30]. The Toxoplasma glideosome-associated connector (GAC) exemplifies this paradigm, coordinating actin dynamics, microneme protein interactions (MIC2), and phosphatidic acid signaling to regulate gliding motility and invasion. PfGAC is also associated with parasite actin to facilitate invasion [31, 32]. However, the functional interplay between plasmodial GAC homologs and HS receptors remains to be elucidated.
This study elucidates the invasion-related functions of Plasmodium berghei glideosome-associated connector (PbGAC; PbANKA_1137800). We demonstrate that PbGAC exhibits specific HS-binding activity and that PbGAC-specific antibodies confer invasion inhibition in murine models. Immunoprecipitation analyses further reveal its association with merozoite invasion machinery. These findings establish PbGAC as a critical mediator of erythrocyte invasion in P. berghei.
Methods
Animals
Female BALB/c mice (2 weeks old) and Sprague–Dawley rats (2 weeks old) were purchased from Liaoning Changsheng Biological Technology Company (Benxi, China). Animal experiments were carried out under the institutional guidelines of the Shenyang Agricultural University, China (ethical approval no. SYXK < Liao > 2021-0010).
Sequence and phylogenetic analysis of PbGAC
The gene sequence of PbGAC (PbANKA_1137800) and orthologs from other Plasmodium species were retrieved from the malaria database (https://plasmodb.org). Multiple sequence alignment of PbGAC was performed using DNAMAN software [33]. Phylogenetic analysis of PbGAC orthologs was conducted using MEGA 11 [34].
Protein expression and purification
The PbGAC (amino acids 99–297) amplified products were cloned into the BamH I/Xho I sites of the pET-28a and pGEX-4T-1 expression vectors to produce recombinant PbGAC-His and GST-PbGAC proteins. Primers used for cloning the coding sequence are provided in Additional File 1: Supplementary Table S1. The plasmids were transformed into BL21 (DE3) or BL21 Escherichia coli strains for protein expression. The PbGAC-His protein was purified via Ni^2+^ affinity chromatography, while GST-PbGAC was purified through glutathione affinity chromatography. The two recombinant proteins were analyzed by sodium dodecyl sulphate–polyacrylamide gel electrophoresis (SDS–PAGE) and western blotting with tag-specific antibodies as described [18].
Generation of PbGAC-specific antibodies
The purified PbGAC-His protein was used to immunize rats (n = 3) and mice (n = 10). For the initial immunization, PbGAC-His protein was emulsified with Freund’s complete adjuvant, and doses of 50 μg per mouse and 100 μg per rat of PbGAC-His protein were administered subcutaneously. This was followed by three additional immunizations, where PbGAC-His protein was emulsified with incomplete Freund’s adjuvant at the same dosages, with a 14-day interval between each administration. Following the final immunization, sera were collected on day 7. Antibody titers were assessed using enzyme-linked immunosorbent assay (ELISA) with recombinant GST-PbGAC as the coating antigen [18]. Immunoglobulin G (IgG) was purified from rat sera utilizing Protein G Sepharose 4 Fast Flow Resin (Cytiva, USA) according to the manufacturer’s protocol. Briefly, 200 μL of Protein G magnetic beads were transferred to a 1.5-mL Eppendorf (EP) tube, followed by the addition of 1 mL of 20 mM sodium phosphate, ensuring a thorough resuspension of the beads. The mixture was centrifuged at 500g for 5 min at 4 °C, and the wash step was repeated twice. The serum was mixed with 20 mM sodium phosphate buffer in a 1:2 ratio, then incubated with the Protein G magnetic beads at 4 °C for 2 h. Subsequently, 1 mL of 20 mM sodium phosphate buffer was added, and the beads were resuspended, centrifuged, and the supernatant discarded; this wash step was repeated four times. Finally, 1 mL of 0.1 mM citric acid was added to the Protein G magnetic beads and incubated at 4 °C for 10 min, followed by centrifugation, and the supernatant was collected into a new EP tube. Immediately, 200 μL Tris-HCL (pH 9.0) was added to neutralize the eluate.
Localization of PbGAC in P. berghei via an immunofluorescence assay
Plasmodium berghei-infected erythrocytes were harvested directly from the blood of infected mice. Smears were fixed with pure methanol for 10 min at room temperature. The cells were permeabilized with 0.1% Triton X-100 for 10 min and blocked with 3% bovine serum albumin (BSA) at 37 ℃ for 1 h. Cells were incubated overnight at 4 ℃ with anti-PbGAC-specific antibodies (1:300). Following incubation, the cells were washed five times with phosphate-buffered saline (PBS) and then incubated with a secondary anti-rat Alexa Fluor 488 antibody (1:500, Thermo Fisher Scientific, USA) for 30 min at 37 °C, followed by five additional washes with PBS. The cells were mounted using ProLong Diamond Antifade Mountant with DAPI (Thermo Fisher Scientific, USA) and visualized under a fluorescence microscope (Leica, Germany).
Heparin-binding and competition assays
Purified GST-PbGAC and GST proteins (200 μg) were respectively mixed with heparin–sepharose beads (50 μL) (Cytiva, USA) and incubated for 2 h at 4 °C in an inverted mixer. The samples underwent seven washes with PBS and were centrifuged at 500g for 5 min at 4 ℃. The samples were then combined with 50 μL of 1× SDS–PAGE loading buffer and heated at 100 °C for 5 min. After brief centrifugation, the supernatant was collected for SDS–PAGE analysis.
To further assess the specificity of GST-PbGAC binding to heparin, GST-PbGAC was incubated with varying concentrations of heparin and chondroitin sulfate (CSA) (10, 1, 0.1, 0.01, 0.001, and 0.0001 mg/mL) for 30 min. Subsequently, 40 μL of heparin–agarose beads were gradually added to the mixture and incubated at 4 °C for 2 h. The samples were washed seven times with PBS and centrifuged at 500g for 5 min at 4 °C. The competition effect of heparin and CSA on the binding of GST-PbGAC to the heparin–agarose beads was analyzed using western blotting.
Binding of recombinant PbGAC to mouse erythrocytes
GST-PbGAC and GST (200 μg, 200 μL) were respectively incubated with mouse erythrocytes (7 μL packed volume) in PBS for 1 h at 37 °C (final volume 1 mL) as previously described [35]. The erythrocytes were washed seven times with PBS and resuspended in 30 μL 1× SDS–PAGE loading buffer, followed by heating at 100 °C for 5 min. The protein was resolved in a 10% acrylamide gel and transferred to the polyvinylidene difluoride (PVDF) membrane as described [18]. The membrane was probed with the anti-GST mAb (1:3,000, TransGen, China), followed by the horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG (H+L) secondary antibody (1:5,000, Beyotime, China). The protein bands were visualized via enhanced chemiluminescence detection.
To further validate the binding of PbGAC to mouse erythrocytes, freshly collected erythrocytes were washed with PBS, and blood smears were prepared and fixed with methanol for 30 s. The fixed erythrocytes were washed in PBS, blocked with 3% bovine serum albumin (BSA) for 1 h at 37 °C, washed again (five times, 10 min each), and incubated with 200 μg GST-PbGAC or GST in 3% BSA for 2 h at 37 °C. The slide samples were washed five times with PBS and incubated with goat anti-GST antibody (1:3,000, TransGen, China), followed by five washes and incubation with Alexa Fluor 488 goat anti-mouse IgG (H + L) (1:500, Thermo Fisher Scientific, USA). High-resolution images were captured using a fluorescence microscope (Leica, Germany), and the mean fluorescence intensity of the erythrocytes was determined using the FACSAria III flow cytometer (BD Biosciences, USA).
Some erythrocytes were treated with heparinase II (0, 1, and 5 U/mL) in Roswell Park Memorial Institute (RPMI)-1640 for 1 h at 37 °C, washed three times in PBS, and incubated with 200 μg GST-PbGAC or GST. Proteins on erythrocytes were detected using western blotting.
Calcium ionophore-induced PbGAC translocation
P. berghei-infected erythrocytes were cultivated in vitro overnight as previously described [36]. Freshly egressed merozoites were subjected to two washes with Dulbecco’s modified Eagle medium (DMEM), then resuspended in DMEM containing 30 μM R59022 (DGK inhibitor) and placed on coverslips within a six-well plate at room temperature for 30 min [31]. After pretreatment, parasites were incubated with 6 μM A23187 (calcium ionophore) for 15 min at 37 ℃ in a 5% CO_2_ atmosphere. Following incubation, samples were fixed for 10 min with methanol. The fixed cells were permeabilized with 0.1% Triton X-100 for 10 min, followed by blocking at 37 ℃ for 1 h with 3% BSA. Subsequently, the cells were incubated overnight at 4 ℃ with anti-PbGAC antibodies diluted in 3% BSA and then treated with a secondary anti-rat Alexa Fluor 488 antibody (1:500, Thermo Fisher Scientific, USA) for 30 min at 37 °C, after which they were washed five times with PBS. The cells were mounted using ProLong Diamond Antifade Mountant with DAPI (Thermo Fisher Scientific, USA) and visualized under a fluorescence microscope (Leica, Germany).
PbGAC secretion assay
To prepare parasite culture supernatant, purified schizonts were incubated in the absence of red blood cells (RBCs) at 37 °C for 5 min with 2% ethanol [37]. Subsequently, the suspension was placed in an ice bath for 10 min to promote further secretion. Cells were centrifuged at 12,000 rpm for 10 min to collect the supernatants. The supernatants were directly subjected to 6% SDS–PAGE. Following electrophoresis, gels were transferred onto PVDF membranes (Bio-Rad). Specific proteins were detected using PbGAC-specific antibodies (1:500), followed by HRP-conjugated secondary antibodies (1:10,000) and enhanced chemiluminescence (Beyotime, China).
Immunoprecipitation and mass spectrometry
Synchronized P. berghei were harvested at the schizont stage via centrifugation at 400g for 20 min [38]. The parasites were pelleted, washed in cold PBS, and subjected to centrifugation at 14,000g for 10 min at 4 ℃ to eliminate hemoglobin. The samples were lysed using NP-40 lysis buffer (Beyotime, China) containing 1% protease inhibitor phenylmethylsulfonyl fluoride (PMSF) for 30 min on ice, followed by centrifugation at 12,000 rpm for 10 min at 4 ℃. The supernatants of the parasite lysates were split into two portions, with one portion incubated with the rat anti-PbGAC antibodies and another incubated with normal rat IgG overnight at 4 ℃ on an inverted mixer. Subsequently, the samples were respectively combined with protein G-conjugated magnetic beads and incubated for 4 h at 4 ℃ on an inverted mixer. Finally, the beads were washed five times with cold PBS and resuspended in 100 μL 1× SDS–PAGE loading buffer, heated at 100 °C for 5 min. The beads were sedimented by brief centrifugation, and the supernatants were separated by SDS–PAGE. After electrophoresis, the gels were stained with Coomassie Brilliant Blue solution. The protein bands were sliced from the gels and subjected to trypsinization and mass spectrometry analysis as previously described [39].
Immunization with PbGAC-His protein and challenge assay
For the initial immunization, PbGAC-His protein emulsified in complete Freund’s adjuvant was administered subcutaneously at a dosage of 50 μg per mouse (n = 10). This was followed by three additional immunizations with 50 μg PbGAC-His protein emulsified in incomplete Freund’s adjuvant, with a 14-day interval between each administration. The ELISA was performed to quantify the antibody titers with GST-PbGAC as the coating antigen as described previously [18]. Once the antibody titer reached 1:32,000, 1 × 10^6^ parasitized erythrocytes per mouse were intraperitoneally injected. Peripheral blood parasitemia was evaluated using Giemsa-stained thin blood smears, and the survival duration of the mice was monitored.
Passive immunization assays with PbGAC-immune sera
A total of 20 mice were randomly allocated into two groups. On the initial day, each mouse received an injection of the immune serum (0.5 mL per mouse) via the tail vein. On the subsequent day, each mouse was administered 1 × 10^6^ infected erythrocytes through an intraperitoneal injection. On the third day, each mouse was intravenously injected with the same amount of the immune serum. The assessment of peripheral blood parasitemia was conducted using Giemsa-stained thin blood smears, and the survival duration of the mice was monitored.
Results
Sequence and phylogenetic analysis of PbGAC
Here, we found that the amino acid sequence of PbGAC is highly conserved among Plasmodium spp., with > 90% identity between P. yoelii, P. chabaudi, and P. vinckei orthologs and > 70% identity across species infecting humans and apes (Fig. 1a). From an evolutionary perspective, PbGAC is closely related to P. yoelii and emerged later than T. gondii (Fig. 1b).Fig. 1. Bioinformatic analyses of PbGAC and homologous proteins from other parasites. A, Percentage identity of the GAC sequence from P. reichenowi, P. gaboni, P. malariae, P. ovale, P. falciparum, P. vinckei, P. chabaudi, P. yoelii, and P. berghei. B, The unrooted phylogenetic tree was inferred from the GAC alignment. The tree was constructed using the neighbor-joining algorithm, on the basis of a hidden Markov model (HMM) multiple alignment. Bootstrap values are shown in blue. The reliability of the branches was assessed by the bootstrap resampling with 1000 replicates
PbGAC is expressed during the asexual development of P. berghei
To elucidate the localization of the PbGAC, we generated specific antibodies against PbGAC by immunization with HIS-tagged PbGAC recombinant protein. The antibody specifically recognized the native PbGAC in western blotting (Fig. 2a). Furthermore, indirect immunofluorescence assays (IFA) were performed to explore the subcellular localization of PbGAC in asexual-stage parasites. Our observations indicated that PbGAC was undetectable in the ring stage and appeared from the trophozoite stages. During the blood-stage merozoite invasion, PbGAC localized to the cytosol, exhibiting a distinct accumulation at the extreme apical region, suggesting a pivotal role for PbGAC in merozoite function (Fig. 2b).Fig. 2PbGAC is mainly expressed in the apical region and cytosol. A, Western blotting analysis of native PbGAC expressed in P. berghei merozoites detected using PbGAC-specific antibodies. B, Indirect immunofluorescence of PbGAC was performed on free merozoites and parasites at ring, trophozoite, and schizont stages, with PbGAC expression detected using anti-PbGAC IgG (green). Parasite nuclei were stained with DAPI (blue). Scale bar, 5 μm
The distribution of PbGAC in the parasite was modulated by calcium ions
The development of parasites depends on calcium-dependent signaling pathways, which influence host-cell invasion and egress from host cells [40]. The Ca^2+^ ionophore A23187 has been previously utilized to study microneme secretion and host-cell invasion in both T. gondii and Plasmodium parasites [41, 42]. To assess whether PbGAC is associated with calcium levels, merozoites were cultured in the presence or absence of A23187, fixed, and stained with PbGAC antibodies. We observed that PbGAC accumulates at the basal end following parasite egress induced by Ca^2+^ ionophore A23187 (Fig. 3). R59022, an inhibitor of the calcium-dependent DGK1 kinase, is known to inhibit both microneme secretion and parasite motility of T. gondii [43]. We examined whether R59022 could affect PbGAC expression and intracellular localization. P. berghei parasites were cultured in the presence of A23187 and R59022, fixed, and stained with PbGAC antibodies. In contrast to the effect observed only with A23187, the relocalization of PbGAC from the apical to the basal region was abolished in the presence of the DGK inhibitor R59022 (Fig. 3).Fig. 3. Altered distribution of PbGAC in the presence of the Ca^2+^ ionophore A23187 and the inhibitor of calcium-dependent DGK1 kinase, R59022. Distribution of PbGAC on both the merozoite cytosol and at the apical region is more intense after exposure to A23187. R59022 treatment results in the inhibition of apical to basal transport. Scale bar, 5 μm
PbGAC interacts with invasion-associated proteins
To identify potential PbGAC-interacting proteins, we performed a pull-down assay using rat PbGAC-specific antibodies with proteins from synchronized P. berghei schizonts, followed by mass spectrometric identification (Fig. 4a). An IgG from a nonimmunized rat was used as a negative control antibody. Over 300 proteins were identified in the sample with the PbGAC-specific antibodies (Supplementary Table S2). Among the proteins identified, reticulocyte binding protein (gene ID no. PbANKA_0501000), high molecular weight rhoptry protein 2 (RhopH2, gene ID no. PbANKA_0830200), RhopH3 (gene ID no. PbANKA_0416000), actin I (gene ID no. PbANKA_1459300), and a transporter protein (SEC24A, gene ID no. PbANKA_1137200) were found to be coprecipitated with PbGAC. Other proteins such as the merozoite attachment protein (MAP1, gen ID no. PbANKA_1425900), the merozoite surface protein 1 (MSP1, gene ID no. PbANKA_0831000), Plasmodium exported protein (PHIST, gene ID no. PbANKA_1229000), and the exported protein (EXP2, gene ID no. PbANKA_1334300) also interacted with PbGAC (Fig. 4b). Immunofluorescence with both PbGAC- and MSP1-specific antibodies confirmed the colocalization of the two molecules on the merozoite surface and the potential interaction (Fig. 4c). To further confirm the interaction between MSP1 and PbGAC, we performed co-immunoprecipitation (IP) experiments using PbGAC antibody as bait. The result showed that MSP1 and PbGAC interact (Fig. 4d). These findings provide compelling evidence for the involvement of PbGAC in interactions with P. berghei merozoite proteins associated with erythrocyte invasion.Fig. 4. Potential PbGAC interactome. A, Schematic overview of the IP. B, Proteins identified in the pulldown assay with the PbGAC-specific antibodies. C, Colocalization of PbGAC and MSP1 in fixed merozoite and schizont stages. Scale bar 5 μm. D, Interactions between PbGAC and MSP1 determined by coimmunoprecipitation (co-IP) and western blotting
PbGAC specifically binds heparin
We hypothesized that PbGAC may function as an invasion ligand through interaction with heparan sulfate (HS) on the erythrocyte surface, as it possesses several heparin-binding motifs (e.g., XBBXBX and XBBBXXBX; B = basic, X = hydropathic) in the amino acid sequence [44]. To test this hypothesis, we generated recombinant GST-PbGAC and GST fusion proteins for heparin-binding assays. The PbGAC encodes a protein of 2,605 amino acids. To determine PbGAC function in heparin-binding activity, we selected amino acid fragments (amino acids 99–297) that are hydrophilic and contain heparin-binding motif (Fig. 5a). The results indicated that the recombinant protein GST-PbGAC exhibited specific heparin-binding activity, whereas the GST protein did not (Fig. 5b). Furthermore, the PbGAC–heparin interaction could be inhibited by heparin in a dose-dependent manner (Fig. 5c), whereas chondroitin sulfate (CSA) could not (Fig. 5d).Fig. 5GST-PbGAC specifically binds to heparin. A, PbGAC is 2605 amino acids in length, GST-PbGAC contains two heparin-binding motifs (purple). B, The heparin-binding capability of GST-PbGAC was assessed via western blotting utilizing an anti-GST antibody. Only GST-PbGAC binds to heparin–sepharose, in contrast to sepharose alone. GST did not bind to heparin–sepharose. C, The binding of GST-PbGAC to heparin–sepharose was inhibited by heparin in a concentration-dependent manner. D, CSA did not show any inhibitory effect on the binding of GST-PbGAC to heparin–sepharose
PbGAC specifically binds to mouse erythrocytes
It has previously been demonstrated that apical organelle-secreted proteins are released into the culture supernatant during host-cell invasion, involving complex interactions between parasite ligands and host receptors [3]. We detected PbGAC in the supernatant (Supplementary Fig. S1). The binding property of PbGAC on erythrocytes was investigated to elucidate its biological functions using GST-PbGAC (Fig. 6a). GST-PbGAC and GST were incubated separately with mouse erythrocytes, and the proteins bound on the erythrocyte surface were analyzed using IFA, western blotting, and flow cytometry. Both IFA and western blotting demonstrated specific binding of GST-PbGAC to the mouse erythrocytes (Fig. 6b, c; Supplementary Fig. S2). This was further confirmed by flow cytometry (Fig. 6d, e), with a gating strategy shown in Supplementary Fig. S3. Conversely, GST alone did not exhibit any binding to the mouse erythrocytes. To confirm that PbGAC mediates heparin interaction, erythrocytes were pretreated with heparinase II before their incubation with GST-PbGAC. Predigestion with heparinase II, which eliminated heparan sulfate from the erythrocyte surface, resulted in a decreased binding affinity of GST-PbGAC to the erythrocytes (Fig. 6f; Supplementary Fig. S4). These findings indicated that GST-PbGAC specifically binds to the erythrocyte surface. To assess whether native PbGAC can bind to heparin, native PbGAC was incubated with heparin–sepharose, and the bound native PbGAC was eluted and analyzed. Native PbGAC was capable of directly binding to heparin (Fig. 6g). Similar to its binding profile with heparin, native PbGAC also bound to mouse erythrocytes (Fig. 6h).Fig. 6PbGAC binds to mouse erythrocytes. A, Purified GST-PbGAC analyzed via SDS–PAGE. B, Indirect immunofluorescence assay (IFA) employing an anti-GST antibody as the primary antibody and Alexa Fluor 488 goat anti-mouse IgG (H+L) as the secondary antibody demonstrated GST-PbGAC binding to erythrocytes, with GST serving as the control. Scale bar, 5 μm. C, Western blotting with an anti-GST antibody confirmed that only GST-PbGAC binds to erythrocytes. D, Flow cytometry analysis revealed a distinct shift in erythrocyte populations bound by GST-PbGAC compared with GST control. E, Fluorescence intensity of erythrocytes bound to GST-PbGAC was compared with that of GST and blank controls. F, Western blotting showed reduced binding efficiency of GST-PbGAC to erythrocyte surfaces following heparinase II treatment. G, Western blotting with an anti-PbGAC antibody confirmed native-PbGAC binding to heparin–sepharose. H, Immunofluorescence assay demonstrated native-PbGAC binding to erythrocytes. Scale bar, 5 μm
Mice immunized with recombinant PbGAC exhibited resistance to P. berghei infection
To evaluate whether PbGAC-specific antibodies confer in vivo protection, groups of BALB/c mice (n = 10/group) were immunized four times with PbGAC-His in Freund’s adjuvant, with a control group receiving only Freund’s adjuvant. Once serum titers of PbGAC-specific antibodies reached 1:32,000, mice were intraperitoneally injected with 1 × 10^6^ P. berghei-infected erythrocytes in 200 μL of 0.9% NaCl. As expected, PbGAC-immunized mice showed prolonged survival (Fig. 7a, b). In addition, two groups received a single intravenous dose of 500 μL immune serum or PBS as a control. The following day, all mice were challenged with 1 × 10^6^ P. berghei-infected erythrocytes. Consistent with previous results, mice injected with anti-PbGAC serum exhibited significantly longer survival than the control group (Fig. 7c, d). These findings suggest that PbGAC antibodies confer protection against P. berghei infection.Fig. 7PbGAC-specific antibodies confer protection against P. berghei infection. A, B, Control BALB/c mice not immunized with PbGAC displayed higher parasitemia than PbGAC-immunized mice; the error bars represent standard deviation (SD). PbGAC-immunized mice survived an average of 5 days longer than the control group. C, D, Mice injected with serum from a normal mouse exhibited higher parasitemia than those receiving anti-PbGAC immune serum. The anti-PbGAC immune serum group survived 5 days longer; the error bars represent SD ^***^P < 0.001. Data were analyzed by log-rank (Mantel–Cox) test
Discussion
The invasion of host erythrocytes constitutes a hallmark biological process for obligate intracellular parasites such as Plasmodium species. Following invasion, merozoites execute a precisely coordinated, multistage invasion process involving preinvasion recognition and active penetration. This intricate mechanism relies on the spatiotemporal regulation of surface-exposed proteins and invasion-associated organellar components, including rhoptries and micronemes [3]. Many of these proteins are anchored via glycosylphosphatidylinositol (GPI) linkages [45]. Several merozoite invasion ligands have been characterized, notably the merozoite surface proteins (MSPs), which constitute a significant component of the merozoite surface coat [3, 9]. Among these, GPI-anchored merozoite surface protein 1 (MSP1) is the largest, forming a dimer exceeding 500 kDa, and is the most abundant [9, 17, 46, 47]. Although MSP1’s precise role remains to be elucidated, the binding affinity of its fragments MSP1-42 and MSP1-33 to heparin imply that heparin-like glycosaminoglycans serve as receptors facilitating erythrocyte invasion [24]. Furthermore, among the key molecular players, MAP1 has been characterized as heparan sulfate (HS)-binding ligand that mediates the initial adhesion of the merozoites to the erythrocyte surface [18], while the PfRh/EBL protein families orchestrate invasion signaling through interactions with erythrocyte receptors such as glycophorins A/B/C and complement receptor 1 [11, 19]. The irreversible host–parasite interface is stabilized by the AMA1–RON complex interaction, where transmembrane RON2 anchors in the erythrocyte membrane to engage merozoite-surface AMA1 [23]. We have revealed that heparin-binding proteins of both *P. falciparum *and P. berghei are critically involved in erythrocyte invasion [18, 48], highlighting protein–glycosaminoglycan interactions as potential therapeutic targets. This study suggests that PbGAC plays a role in the binding of HS during erythrocyte invasion.
PbGAC is predominantly localized at the apical region and in the cytosol of P. berghei merozoites, suggesting its potential involvement in host–pathogen interactions. Plasmodium falciparum reticulocyte-binding protein homolog 1 (PfRH1) and erythrocyte-binding protein EBA175 are critical mediators of erythrocyte invasion by the parasite [19, 49, 50]. Unlike other EBA/PfRh ligands that exhibit functional redundancy [51], PfRH1 specifically triggers calcium signaling pathways, leading to the release of EBA175 and facilitating junction formation between the erythrocyte and the parasite [52]. We analyzed calcium flux triggered by A23187. Calcium flux-triggered microneme secretion represents a critical regulatory node for apicomplexan motility and invasion [53]. Pharmacological modulation using Ca^2+^ ionophore A23187 induced PbGAC translocation to the basal pole during egress, a process disrupted by DGK1 inhibition via the R59022 compound [43]. This parallels TgGAC dynamics, suggesting conserved regulatory mechanisms. Calcium ions are required at two critical stages during the intraerythrocytic asexual life cycle of Plasmodium: during the initial invasion of the red blood cell and the initiation of egress [20, 54, 55]. In most cases, the Ca^2+^ flux was an excellent indicator of successful invasion. The transient, motility-dependent basal accumulation of PbGAC and observed immune protection in PbGAC-His vaccinated mice (Fig. 7) further underscore its functional significance. Our immunoprecipitation studies reveal PbGAC’s interactome, encompassing MSP1, MAP1, RhopH complex components, RAP1, and actin-regulatory proteins. This molecular synergy aligns with emerging models of moving junction formation [56, 57], where apical-to-basal protein translocation facilitates mechanical penetration.
Several studies have demonstrated that heparin-like molecules inhibit erythrocyte invasion by blood-stage Plasmodium spp. [24, 58, 59]. These findings imply that the heparin-like molecules play an important role in invasion inhibition and possess potential as novel antimalarial therapeutics. Nonetheless, only a limited number of merozoite proteins have been characterized for their capacity to bind heparin-like molecules and facilitate erythrocyte invasion. Surface antigens, including merozoite surface protein 1 (MSP1), are hypothesized to mediate initial erythrocyte attachment [3], potentially serving as receptors for heparin-like molecules. Kobayashi et al. reported that heparin binds at the apical pole of the merozoite surface, with multiple heparin-binding proteins localized preferentially within apical organelles [28]. Our data indicate that PbGAC is expressed during the merozoite stage, with partial localization to the merozoite apical region and cytosol. Furthermore, we demonstrated direct binding between PbGAC and heparin-agarose beads, suggesting that PbGAC binds to the erythrocyte surface. Immunofluorescence assays, western blotting, and flow cytometry confirmed that PbGAC directly binds to the erythrocyte surface. In addition, the binding of GST–GAC with erythrocytes was sensitive to heparinase II pretreatment, indicating dependence on heparan sulfate. The initial attachment is a pivotal step requiring both interaction specificity and cellular process efficiency. Our findings suggest that PbGAC fulfills the complex molecular criteria for initial erythrocyte attachment: it is localized on the merozoite cytosol and apical region, highly conserved across the Plasmodium spp., and remarkably specific for HS binding.
Conclusions
Our findings demonstrate a novel protein, PbGAC, predominantly expressed in the P. berghei merozoite cytosol and the apical region, which is associated with erythrocyte invasion. The distribution of PbGAC in the parasite is modulated by calcium ions. Furthermore, PbGAC specifically interacts with the heparan sulfate receptor on erythrocytes, implying a common mechanism of malarial parasite invasion. The significance of PbGAC in the invasion process is evidenced by its strong interaction with several invasion-associated proteins and the protective effect of specific antibodies against parasite invasion. These mechanistic insights not only advance our understanding of Plasmodium invasion biology but also establish an experimental framework for the systematic functional characterization of P. falciparum virulence factors, potentially accelerating the development of antimalarial therapeutics targeting host–pathogen interactions.
Supplementary Information
Additional file 1: Fig. S1. PbGAC is secreted into the supernatant. The culture supernatant (SN) was stained with anti-PbGAC antibody and analyzed by western blotting.Additional file 2: Fig. S2. The binding of GST-PbGAC to erythrocytes in SDS-PAGE gel.Additional file 3: Fig. S3. Flow cytometry gating strategy.Additional file 4: Fig. S4. The effect of heparinase II treatment on the binding of GST-PbGAC to erythrocytes in SDS-PAGE gel.Additional file 5: Table S1: Primers used for PbGAC cloning.Additional file 6: Table S2: Identification of proteins interacting with PbGAC.