GlcNAc induces GlcNAc catabolic genes and inhibits filamentation via YlRep1-YlNgs1 signaling in the dimorphic yeast Yarrowia lipolytica
Zhen-Hua Wang, Meng-Yang Xu, Xiang-Dong Gao

TL;DR
This study shows that GlcNAc inhibits filamentation in the yeast Yarrowia lipolytica through a signaling pathway involving YlRep1 and YlNgs1, which also activates GlcNAc catabolic genes.
Contribution
The novel finding is that YlRep1-YlNgs1 signaling in Y. lipolytica represses filamentation-related genes and inhibits filamentation, independent of GlcNAc catabolism or pH changes.
Findings
YlRep1 and YlNgs1 are essential for inducing GlcNAc catabolic genes in Y. lipolytica.
GlcNAc inhibits filamentation in Y. lipolytica via YlRep1-YlNgs1 signaling, independent of pH changes.
YlRep1-YlNgs1 represses filamentation-related genes without requiring GlcNAc catabolism.
Abstract
Many fungal species can utilize N-acetylglucosamine (GlcNAc) as a carbon source. Studies in the pathogenic yeast Candida albicans have revealed that GlcNAc utilization and the induction of GlcNAc catabolic genes depend on the Ndt80 family transcription factor CaRep1 and the histone acetyltransferase CaNgs1. Additionally, GlcNAc induces filamentation via both alkalinization of the medium and CaNgs1 signaling. However, the roles of YlRep1 and YlNgs1 in GlcNAc catabolism and filamentous growth are not clear in the dimorphic yeast Yarrowia lipolytica. In this study, we demonstrate that YlRep1 and YlNgs1 are essential for the induction of GlcNAc catabolic genes in Y. lipolytica, which is similar to the function of CaRep1 and CaNgs1. YlRep1 and YlNgs1 interact physically and exhibit transcriptional activation activity on a reporter gene. Interestingly, unlike in C. albicans, GlcNAc inhibits…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
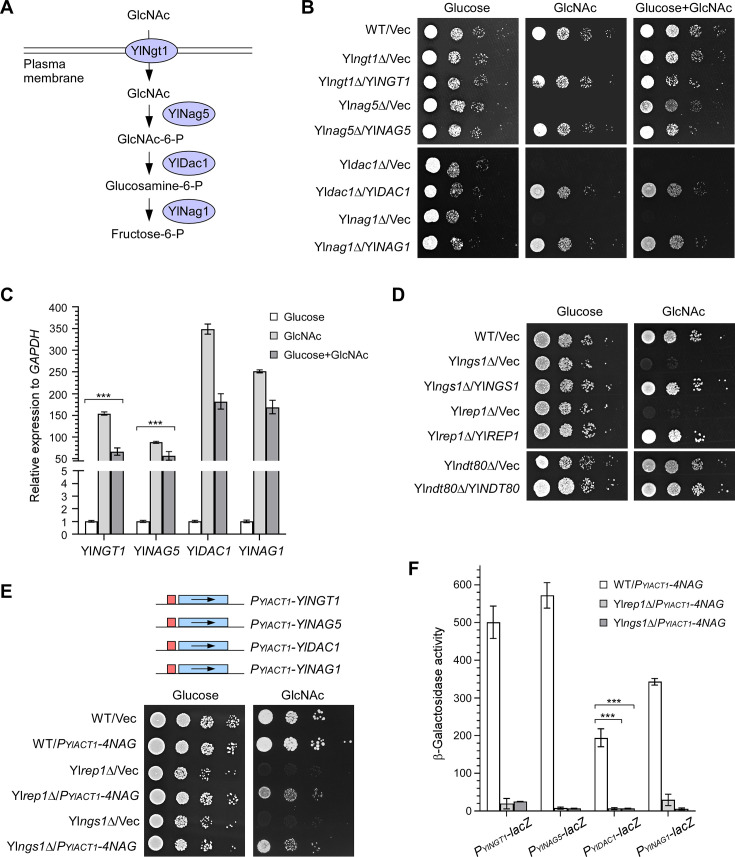 Fig 1
Fig 1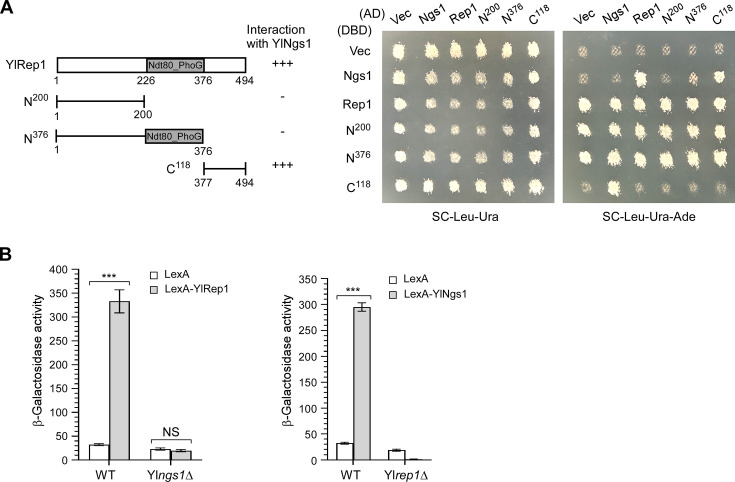 Fig 2
Fig 2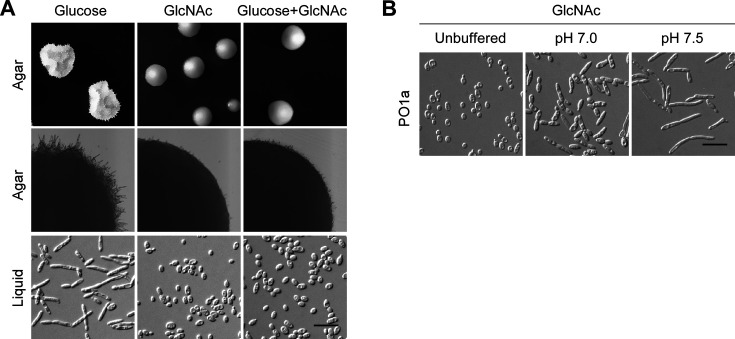 Fig 3
Fig 3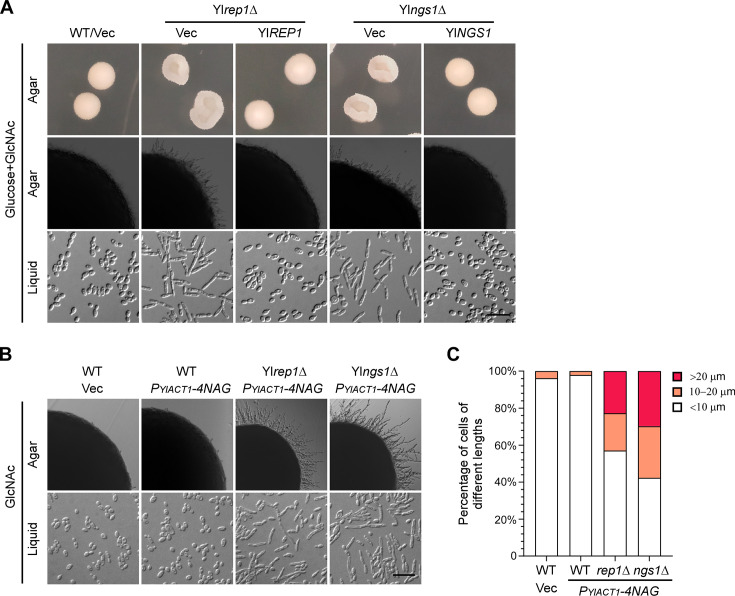 Fig 4
Fig 4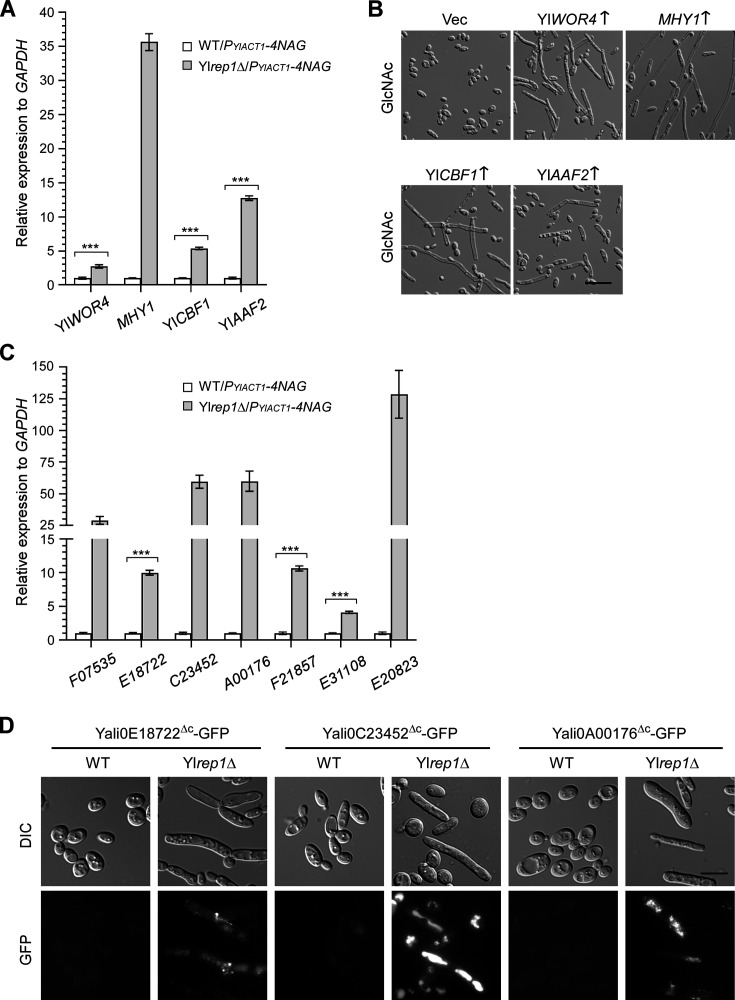 Fig 5
Fig 5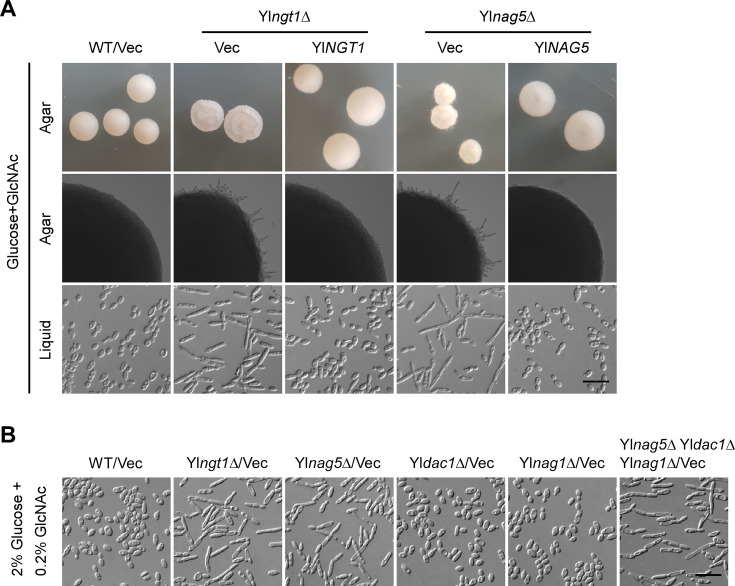 Fig 6
Fig 6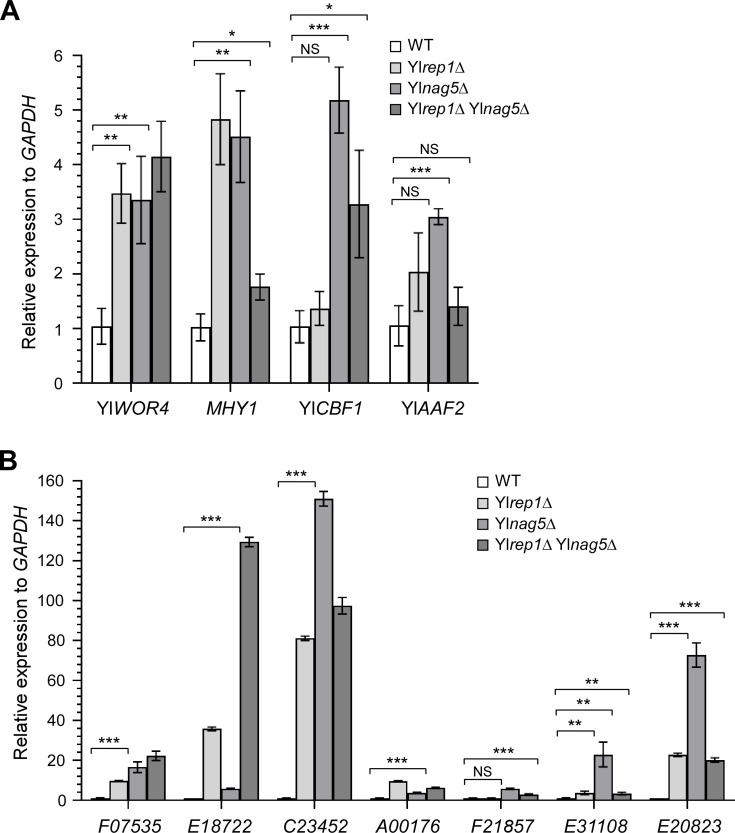 Fig 7
Fig 7| Carbon source | Initial pH | Final pH | Decrease in pH | Percentage of cells longer than 20 µm |
|---|---|---|---|---|
| 1% GlcNAc | 5.36 | 4.93 | 0.43 | 0 |
| 1% glucose | 5.47 | 4.81 | 0.66 | 28 |
| 1% GlcNAc | 7.0 (Na2HPO4-citric acid buffer) | 6.78 | 0.22 | 7 |
| 1% glucose | 7.0 (Na2HPO4-citric acid buffer) | 6.05 | 0.95 | 53 |
| Direction of regulation | Transcription factor genes | Cell wall protein genes |
|---|---|---|
| Up (≥2-fold) | ||
| Down (≥2-fold) |
|
| Genes | YlNgs1 repressed only | YlNgs1-YlRep1 co-repressed | YlRep1 repressed only |
|---|---|---|---|
| Transcription factor genes | |||
| Cell wall protein genes | None |
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal and yeast genetics research · Glycosylation and Glycoproteins Research · Polysaccharides and Plant Cell Walls
INTRODUCTION
Yarrowia lipolytica is a nonconventional yeast with genomic, physiological, and metabolic characteristics that differ from those of the model yeasts Saccharomyces cerevisiae and Candida albicans (1). It has a wide range of industrial applications in the production of heterologous proteins, organic acids, terpenes, and biofuels (2, 3). Like other dimorphic yeasts, Y. lipolytica can switch from oval-shaped yeast form to filamentous forms, which is induced by environmental factors, such as carbon source, nitrogen source, temperature, pH, low oxygen, or osmotic pressure (4, 5). Dimorphic transition is considered a strategy for fungi to acquire nutrients and adapt to the environment (6). For some pathogenic fungi, such as the human pathogen C. albicans, the formation of hyphae promotes invasion into tissues and biofilm formation (7, 8).
Y. lipolytica can utilize different carbon sources, and cells exhibit different morphologies depending on the type of carbon sources available in the environment. In the presence of carbon sources, such as oleate, olive oil, and castor oil, Y. lipolytica cells typically remain in the oval-shaped yeast form (9, 10). However, when exposed to carbon sources, such as glucose, fructose, peptone, lactate, and citrate, Y. lipolytica cells elongate and develop filaments (5, 10). The molecular mechanisms that govern this morphological transition remain unclear. A recent study suggests that glucose-induced filamentation in Y. lipolytica is regulated by sugar signaling pathways, most likely via the cAMP-PKA-dependent pathway (11). Among the common carbon sources, glycerol is a preferred choice for Y. lipolytica, and cells grown in glycerol are in the yeast form (10). The conserved nutrient-sensing TORC1-Sch9 signaling pathway is involved in the inhibition of filamentation by glycerol (10).
N-acetylglucosamine (GlcNAc) is another carbon source that Y. lipolytica can utilize. This amino sugar is the building block of important biological molecules, such as cell wall peptidoglycan in bacteria, cell wall chitin in fungi and parasites, exoskeletons of arthropods, and the extracellular matrix of animal cells (12, 13). GlcNAc can be utilized as a carbon source and nitrogen source by most bacteria and fungi, except for some species such as the model yeasts S. cerevisiae and Schizosaccharomyces pombe, because they lack GlcNAc catabolic genes (13). In fungi, GlcNAc is catabolized to fructose-6-phosphate in a stepwise manner by GlcNAc transporter, GlcNAc kinase, GlcNAc-6-phosphate deacetylase, and glucosamine-6-phosphate deaminase, which are encoded by NGT1, HXK1/NAG5, DAC1/NAG2, and NAG1, respectively (13–15). The phosphorylated form of GlcNAc, GlcNAc-6-phosphate, is a precursor of uridine diphosphate-GlcNAc (UDP-GlcNAc), which is important for synthesizing cell wall chitin in fungi and the extracellular matrix in animal cells, as well as other cellular processes. There is a reverse pathway to synthesize GlcNAc-6-phosphate from fructose-6-phosphate, which is important when GlcNAc is absent in the environment (13).
In addition to the roles of an energy source and components of cell-surface structure, extracellular GlcNAc is also a signaling molecule. GlcNAc can downregulate the production of curli, extracellular surface fibers that function in biofilm formation in the bacteria Escherichia coli (16). In animal cells, increased GlcNAc can stimulate O-linked attachment of GlcNAc to nuclear and cytoplasmic proteins, which modulates signaling and influences protein expression, degradation, and trafficking (17). In the human pathogen C. albicans, GlcNAc induces filamentation, white-opaque transition, and GlcNAc-induced cell death (15). GlcNAc is also a potent inducer of the yeast-to-filament transition in two thermally dimorphic fungi, Histoplasma capsulatum and Blastomyces dermatitidis (18).
The fungal GlcNAc sensor Ngs1 was first discovered in C. albicans (19). It is an N-acetyltransferase related to the S. cerevisiae histone acetyltransferase Gcn5. GlcNAc binds to CaNgs1 and activates its N-acetyltransferase activity. CaRep1, a negative regulator of MDR1 transcription, is essential for growth on GlcNAc in C. albicans (19, 20). It is an Ndt80 family transcription factor, which also includes CaNdt80 and CaRon1 (21, 22). Because CaNgs1 does not bind to DNA, CaRep1 recruits CaNgs1 to the promoters of target genes, where CaNgs1 activates gene expression by histone acetylation (19). The Rep1-Ngs1 signaling is essential for GlcNAc utilization and the induction of GlcNAc catabolic genes in response to GlcNAc. In the filamentous fungus Trichoderma reesei, Ngs1 and the transcription factor Ron1 (ortholog of C. albicans CaRep1 but not CaRon1) are also essential for GlcNAc catabolism (23, 24).
In addition to GlcNAc utilization, CaNgs1 but not CaRep1 is also important for GlcNAc-induced filamentation (19, 22). Cangs1Δ cells and Cangs1Δ Cahxk1Δ cells were severely defective in hyphal development in medium containing both GlcNAc and galactose. Moreover, CaNgs1 is essential for the induction of hyphal-specific genes by GlcNAc (19). In Candida tropicalis, which is closely related to C. albicans, GlcNAc strongly inhibits hyphal growth (25), and this inhibition requires CtRep1 (26). The Ndt80 family transcription factor CtRon1 also regulates filamentation. However, it is required for serum-induced filamentation but not for the inhibition of filamentation by GlcNAc (26).
Here, we show that, similar to C. albicans, YlRep1-YlNgs1 is necessary for the induction of GlcNAc catabolic genes in response to GlcNAc in Y. lipolytica. However, unlike in C. albicans, GlcNAc inhibits filamentation in Y. lipolytica. This inhibition is independent of GlcNAc catabolism but requires both YlRep1-YlNgs1 and the GlcNAc kinase YlNag5. Therefore, YlRep1-YlNgs1 plays dual and opposing roles in GlcNAc-regulated catabolism and dimorphic transition in Y. lipolytica.
RESULTS
Y. lipolytica shares a similar GlcNAc catabolic pathway and YlRep1-YlNgs1 regulation of GlcNAc catabolic genes with C. albicans
Similar to C. albicans, GlcNAc catabolism in Y. lipolytica is thought to require stepwise reactions catalyzed by YlNgt1, YlNag5, YlDac1, and YlNag1 (14) (Fig. 1A). We found that Ylngt1Δ, Ylnag5Δ, Yldac1Δ, and Ylnag1Δ cells failed to grow in GlcNAc medium but grew well in glucose medium (Fig. 1B), indicating that the four proteins are essential for GlcNAc utilization. Yldac1Δ and Ylnag1Δ cells also failed to grow in the medium containing both glucose and GlcNAc (Fig. 1B), resembling the growth defect observed in C. albicans Cadac1Δ and Canag1Δ cells (27). This finding suggests that the accumulation of metabolic intermediates is toxic to the cells. Similar to what is observed in C. albicans, GlcNAc strongly induced the expression of YlNGT1, YlNAG5, YlDAC1, and YlNAG1 genes in Y. lipolytica (Fig. 1C). Interestingly, the strong induction of these genes by GlcNAc persisted even in the presence of a high concentration of glucose (1%). These results suggest that Y. lipolytica shares a similar GlcNAc catabolic pathway with C. albicans.
GlcNAc induces GlcNAc catabolic genes in a YlRep1- and YlNgs1-dependent manner in Y. lipolytica. (A) The GlcNAc catabolic pathway in Y. lipolytica. (B) YlNGT1, YlNAG5, YlDAC1, and YlNAG1 genes are essential for growth in GlcNAc medium. Wild-type (WT), Ylngt1Δ, Ylnag5Δ, Yldac1Δ, and Ylnag1Δ cells carrying pINA445 (Vec) or pINA445-Gene were spotted at 1:10 serial dilution on YNB agar containing the indicated sugar. Pictures were taken after 2 days at 30°C. (C) GlcNAc induces the expression of YlNGT1, YlNAG5, YlDAC1, and YlNAG1 genes. The transcription levels of these genes in wild-type cells grown in liquid YNB medium containing the indicated sugar were determined by qRT-PCR and normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH). (D) GlcNAc catabolism depends on YlRep1 and YlNgs1 but not YlNdt80. Wild-type (WT), Ylngs1Δ, Ylrep1Δ, and Ylndt80Δ cells carrying pINA445 (Vec) or pINA445-Gene were spotted at 1:10 serial dilution and grown on YNB-Glucose and YNB-GlcNAc agar for 2 d at 30°C. (E) Ylngs1Δ and Ylrep1Δ cells carrying PYlACT1-4NAG grow in GlcNAc medium. Wild-type (WT), Ylrep1Δ, and Ylngs1Δ cells carrying pYL27 (Vec) or pYL27-4NAG (PYlACT1-4NAG) were spotted at 1:10 serial dilution and grown on YNB-Glucose and YNB-GlcNAc agar for 2 days at 30°C. The plasmid pYL27-4NAG carries YlNGT1, YlNAG5, YlDAC1, and YlNAG1 genes under the control of the YlACT1 promoter. (F) YlRep1 and YlNgs1 are essential for the induction of GlcNAc catabolic genes by GlcNAc. Wild-type (WT), Ylrep1Δ, and Ylngs1Δ cells carrying both pYL25-4NAG (PYlACT1-4NAG) and pINA445-PGene-lacZ were grown in YNB-GlcNAc medium. β-Galactosidase activities of promoter-lacZ constructs were measured. All the spot plate assays were done at least three times on different days. For qRT-PCR and β-galactosidase assays, mean data ± standard deviation from three independent experiments carried out on different days were plotted. The unpaired two-tailed Student’s t-test was used to examine the statistical significance of the difference between two samples. Statistically significant differences are indicated by the asterisks (**, P < 0.001).*
In C. albicans, the histone acetyltransferase CaNgs1 serves as the GlcNAc sensor and transducer (19). CaNgs1 is constitutively targeted to the promoters of GlcNAc-inducible genes via the Ndt80 family transcription factor CaRep1, and both CaNgs1 and CaRep1 are required for the transcriptional activation of GlcNAc catabolic genes (19, 21). We identified YlNgs1 (YALI0E20185p, 28.0% identity) and YlRep1 (YALI0D24860p, 24.9% identity) as the Y. lipolytica orthologs of C. albicans CaNgs1 and CaRep1, respectively. Y. lipolytica also contains another Ndt80 family transcription factor, YlNdt80 (YALI0B14773p), which shares 34.0% and 31.8% amino acid sequence identity to CaNdt80 and CaRon1, respectively, but very low homology to CaRep1.
Similar to C. albicans Carep1Δ and Cangs1Δ cells, Y. lipolytica Ylrep1Δ and Ylngs1Δ cells were severely impaired in growth on GlcNAc medium (Fig. 1D), indicating that YlRep1 and YlNgs1 may have similar roles in regulating GlcNAc catabolism as their orthologs in C. albicans. In contrast, Ylndt80Δ grew well on GlcNAc medium (Fig. 1D), suggesting that YlNdt80 is not involved in GlcNAc catabolism.
To examine whether YlRep1 and YlNgs1 are required for the induction of GlcNAc catabolic genes by GlcNAc, and more importantly, to examine the morphology of Ylrep1Δ and Ylngs1Δ cells grown in GlcNAc medium (see later section), we constructed a plasmid that carries YlNGT1, YlNAG5, YlDAC1, and YlNAG1 genes, each of them was under the control of the strong constitutive promoter of YlACT1 (encodes actin) (Fig. 1E, upper panel). As expected, growth was restored in Ylrep1Δ and Ylngs1Δ cells carrying P_YlACT1_-4NAG in GlcNAc medium, although it was significantly slower than that of wild-type cells (Fig. 1E, lower panel). By using promoter-lacZ reporters, we found that P_YlNGT1_-lacZ, P_YlNAG5_-lacZ, P_YlDAC1_-lacZ, and P_YlNAG1_-lacZ reporters exhibited high levels of expression in wild-type cells grown in GlcNAc medium. In contrast, their expression levels were very low in Ylrep1Δ or Ylngs1Δ cells carrying P_YlACT1_-4NAG (Fig. 1F). This result indicates that YlRep1 and YlNgs1 are essential for the induction of GlcNAc catabolic genes in response to GlcNAc.
Together, our results suggest that the GlcNAc catabolic pathway and YlRep1-YlNgs1 regulation of GlcNAc catabolic genes in Y. lipolytica are similar to those of C. albicans.
YlRep1 and YlNgs1 interact physically, and YlRep1-YlNgs1 activates a reporter gene interdependently
In C. albicans, CaRep1 recruits CaNgs1 to the promoters of GlcNAc-inducible genes (19). Similar to CaRep1 and CaNgs1, yeast two-hybrid analysis showed that YlRep1 and YlNgs1 also interact physically (Fig. 2A). Surprisingly, this interaction appears to be mediated by the C-terminus of YlRep1 rather than the N-terminus. This is unexpected, as a recent report showed that CaNgs1 binds to the N-terminus of CaRep1 (28). This discrepancy suggests that notable differences exist between YlRep1 and CaRep1.
YlRep1 interacts with YlNgs1, and YlRep1-YlNgs1 activates a reporter gene in an interdependent manner. (A) Two-hybrid assay between YlRep1 and YlNgs1. pGBDU-C1 (Vector, containing DNA-binding domain, DBD), pGBDU-YlNGS1, pGBDU-YlREP1, and YlREP1 segments were paired with pGAD-C1 (Vector, containing activation domain, AD), pGAD-YlNGS1, pGAD-YlREP1, and YlREP1 segments. Cells were grown on an SC-Leu-Ura plate and then replica plated to an SC-Leu-Ura-Ade plate. Pictures were taken after 2 days at 30°C. The YlRep1 segments are depicted. The strength of interaction is indicated from strong (+++) to no effect (−). Note that full-length YlRep1, YlRep1-N200, and YlRep1-N376 exhibited auto-activation activity when fused to Gal4-DBD. The two-hybrid assay was done three times on different days. (B) YlRep1 and YlNgs1 activate lexAop-PYlLEU2-lacZ expression in an interdependent manner. Plasmids pYL21-LexA and pYL21-LexA-YlREP1 were transformed into wild-type and Ylngs1Δ cells carrying the reporter plasmid pINA445-lexAop-PYlLEU2-lacZ. Similarly, plasmids pYL21-LexA and pYL21-LexA-YlNGS1 were transformed into wild-type and Ylrep1Δ cells carrying the reporter plasmid pINA445-lexAop-PYlLEU2-lacZ. Cells were grown in liquid YNB-Glucose + GlcNAc medium. Cell lysates were measured for β-galactosidase activity. Mean data ± standard deviation from three independent experiments done on different days were plotted. The unpaired two-tailed Student’s t-test was used to examine the statistical significance of the difference between two samples. Statistically significant differences are indicated by the asterisks (**, P < 0.001). NS, not statistically significant.*
To determine whether YlRep1 and YlNgs1 may have transcriptional activation activity, we monitored the ability of LexA-YlRep1 and LexA-YlNgs1 fusion proteins (containing the DNA-binding domain of LexA, a.a. 1-87) to influence the expression of a lexAop-P_YlLEU2_-lacZ reporter in Y. lipolytica. In medium containing both glucose and GlcNAc, LexA-YlRep1 increased reporter expression by 10.3-fold, and LexA-YlNgs1 increased reporter expression by 9.1-fold in wild-type cells (Fig. 2B), indicating that they activate gene expression. In contrast, LexA-YlRep1 failed to increase reporter expression in Ylngs1Δ cells (Fig. 2B), indicating that YlRep1 depends on YlNgs1 for gene activation. Interestingly, LexA-YlNgs1 also failed to increase reporter expression in Ylrep1Δ cells (Fig. 2B), indicating that YlRep1 is also required for gene activation by YlNgs1. The dependency on YlRep1 is likely due to the requirement of GlcNAc binding to YlNgs1 to activate gene expression (19). GlcNAc may not accumulate efficiently in Ylrep1Δ cells. Alternatively, YlRep1 may contribute to gene activation through other mechanisms. These results suggest that YlRep1 and YlNgs1 are interdependent for transcriptional activation.
Together, our results suggest that YlRep1 and YlNgs1 interact physically and work together to activate gene expression.
GlcNAc inhibits filamentation in Y. lipolytica
GlcNAc is a strong inducer of filamentous growth in C. albicans (29). Based on this finding, GlcNAc has also been used previously to induce filamentation in Y. lipolytica, but often in combination with a citrate buffer at near-neutral pH (4, 30, 31). However, we found that GlcNAc alone is not a good inducer of filamentation. In contrast to glucose, which promotes filamentation in Y. lipolytica, GlcNAc inhibits filamentation as wild-type cells were in oval-shaped yeast form and did not elongate or form filaments in liquid or solid GlcNAc media as they did in glucose media (Fig. 3A). In addition, wild-type cells cultivated in liquid or solid media containing both glucose and GlcNAc displayed morphology identical to that of cells cultivated in GlcNAc media (Fig. 3A), indicating that GlcNAc can inhibit filamentation induced by glucose. The inhibition of filamentation by GlcNAc is not due to background mutations in the lab strain PO1a, as the wild-type strain W29 displayed the same cell morphology as strain PO1a under the same growth conditions (Fig. S1).
GlcNAc inhibits filamentation in Y. lipolytica. (A) GlcNAc inhibits filamentation. Wild-type strain PO1a was grown on YNB agar containing the indicated sugar for 3 days (upper row) or 2 days (middle row) or in liquid medium for 16 h (bottom row) at 30°C. (B) Alkaline pH induces filamentation in GlcNAc medium. Wild-type strain PO1a carrying pINA445 (Vec) was grown in liquid YNB-GlcNAc medium (unbuffered, pH ~5.4) or YNB-GlcNAc medium buffered to pH 7.0 or pH 7.5 with Na2HPO4-citric acid buffer for 16 h at 30°C. Bars, 20 µm. All the assays were done at least three times on different days.
Although GlcNAc inhibits filamentation, alkaline pH can still induce filamentation in cells grown in GlcNAc medium (Fig. 3B). Wild-type cells grown in yeast nitrogen base (YNB)-GlcNAc medium buffered at pH 7.0 were elongated compared to those grown in unbuffered medium (pH ~5.4). 2% of cells (n > 200) grown in unbuffered medium were longer than 10 µm. In contrast, 65% and 7% of cells (n > 200) grown in GlcNAc medium buffered at pH 7.0 were longer than 10 and 20 µm, respectively. Cells grown in GlcNAc medium buffered at pH 7.5 formed long filaments, and 82% of cells (n > 200) were longer than 20 µm. This finding agrees with previous reports that filamentation can be induced in GlcNAc medium by alkaline pH and supports the idea that the “filament-inducing” effect of GlcNAc is due to neutral pH (see Discussion).
GlcNAc inhibition of filamentation does not depend on the alteration of ambient pH
In C. albicans, in contrast to glucose catabolism, which decreases the pH of the culture medium, GlcNAc catabolism raises the pH of the medium, which indirectly induces filamentation (32). To investigate whether GlcNAc catabolism may also alter ambient pH in Y. lipolytica, we determined the initial pH of the culture medium and the final pH after 16 h of cultivation with the wild-type strain at 30°C. In both unbuffered media (pH ~5.4) and buffered pH 7.0 media with GlcNAc or glucose as the sole carbon source, there was a slight decrease in pH after the cultivation, and GlcNAc catabolism decreased ambient pH less than glucose catabolism (Table 1). Although GlcNAc did not induce filamentation in unbuffered medium and induced a low degree of filamentation in buffered pH 7.0 medium, glucose induced filamentation in both unbuffered and buffered pH 7.0 media (Table 1). This result suggests that the slight decrease in ambient pH by GlcNAc catabolism is unlikely to be the major cause of GlcNAc inhibition of filamentation. Therefore, the inhibition of filamentation by GlcNAc does not depend on the alteration of ambient pH in Y. lipolytica, which is different from C. albicans.
YlRep1-YlNgs1 is important for the inhibition of filamentation by GlcNAc
Since YlRep1-YlNgs1 signaling is important for GlcNAc-induced expression of GlcNAc catabolic genes, we asked whether YlRep1-YlNgs1 is required for the inhibition of filamentation by GlcNAc. To this end, we examined the morphology of Ylrep1Δ and Ylngs1Δ cells grown in media containing both glucose and GlcNAc. In contrast to wild-type cells that formed smooth colonies devoid of radial filaments on agar and were oval-shaped yeast form in liquid medium, Ylrep1Δ and Ylngs1Δ cells formed wrinkled colonies with short radial filaments on agar and were elongated and formed filaments in liquid medium (Fig. 4A). This result suggests that both YlRep1 and YlNgs1 are required for the inhibition of filamentation by GlcNAc.
YlRep1 and YlNgs1 play a key role in the inhibition of filamentation by GlcNAc. (A) Morphology of Ylrep1Δ and Ylngs1Δ colonies and cells grown in Glucose + GlcNAc medium. Wild-type (WT), Ylrep1Δ, and Ylngs1Δ cells carrying pINA445 (Vec) or pINA445-Gene were grown on YNB-Glucose + GlcNAc agar for 3 days (upper row) or 2 days (middle row) or in liquid medium for 16 h (bottom row) at 30°C. (B) Ylrep1Δ and Ylngs1Δ cells exhibited increased filamentation compared to wild-type cells in GlcNAc medium. Wild-type (WT), Ylrep1Δ, and Ylngs1Δ cells carrying pYL27 (Vec) or pYL27-4NAG (PYlACT1-4NAG) were grown in liquid YNB-GlcNAc medium for 1 day at 30°C. Bar, 20 µm. (C) Percentage of cells of different lengths as in panel B. The cell lengths for each strain were categorized into three groups: <10 µm, 10–20 μm, and >20 µm. All the assays were done at least three times on different days. The data shown are representative of independent experiments.
We also examined the morphology of wild-type, Ylrep1Δ, and Ylngs1Δ cells carrying P_YlACT1_-4NAG, which grew in GlcNAc medium (Fig. 1E). On GlcNAc agar, wild-type cells carrying P_YlACT1_-4NAG or empty vector formed no radial filaments. In contrast, Ylrep1Δ cells and Ylngs1Δ cells carrying P_YlACT1_-4NAG formed long radial filaments (Fig. 4B, upper panel). In liquid GlcNAc medium, wild-type cells carrying P_YlACT1_-4NAG or empty vector were in oval-shaped yeast form. Only 4% of wild-type cells carrying an empty vector and 2% of wild-type cells carrying P_YlACT1_-4NAG (n > 400) were in the range of 10–20 μm in length. None of them was longer than 20 µm. In contrast, Ylrep1Δ cells and Ylngs1Δ cells carrying P_YlACT1_-4NAG formed short filaments. 20% of Ylrep1Δ cells carrying P_YlACT1_-4NAG and 28% of Ylngs1Δ cells carrying P_YlACT1_-4NAG were in the range of 10–20 μm in length. Moreover, 23% of Ylrep1Δ cells carrying P_YlACT1_-4NAG and 30% of Ylngs1Δ cells carrying P_YlACT1_-4NAG were longer than 20 µm (Fig. 4B and C). These results suggest that YlRep1 and YlNgs1 are important for the inhibition of filamentation by GlcNAc.
YlRep1 and YlNgs1 repress a set of transcription factors and cell wall protein genes, some of which are associated with filamentation
To investigate how YlRep1 and YlNgs1 inhibit filamentation, we wanted to determine whether YlRep1 and YlNgs1 regulate the genes associated with filamentation. To this end, we conducted RNA-Seq analyses in wild-type, Ylrep1Δ, and Ylngs1Δ cells carrying P_YlACT1_-4NAG grown in liquid GlcNAc medium. In Ylrep1Δ cells, 327 genes exhibited significant differential expression (≥2-fold, P < 0.05) compared to wild-type cells. Of these, 213 genes (65.1%) were upregulated, whereas 114 genes (34.9%) were downregulated (Table S1). In Ylngs1Δ cells, 556 genes exhibited significant differential expression (≥2-fold, P < 0.05) compared to wild-type cells. Of these, 362 genes (65.1%) were upregulated, whereas 194 genes (34.9%) were downregulated in Ylngs1Δ cells (Table S2). YlRep1 shares a large portion of target genes with YlNgs1. YlRep1 and YlNgs1 co-repress 188 genes, which account for 88.3% of YlRep1-repressed genes but only 51.9% of YlNgs1-repressed genes (Fig. S2). This result suggests that YlRep1 regulates gene expression mainly by recruiting YlNgs1 to target genes, whereas YlNgs1 can also be recruited to target genes by other transcription factors.
Transcription factors play important roles in regulating gene expression. YlRep1 repressed 17 transcription factor genes and highly repressed four of them (≥5-fold, P < 0.05) (Table 2). Fourteen of the 17 YlRep1-repressed genes (82.4%) are also repressed by YlNgs1 (Table 3). Among these genes, YlWOR4 (YALI0F19822), MHY1, YlCBF1 (YALI0B3354), YlAAF2 (YALI0E06105), and YALI0E03410 were the top genes that exhibited the highest read counts. The upregulation of YlWOR4, MHY1, YlCBF1, and YlAAF2 was confirmed by quantitative reverse transcription PCR (qRT-PCR) analysis (Fig. 5A). While wild-type cells remained in the yeast form in liquid GlcNAc medium, the overexpression of YlWOR4, MHY1, YlCBF1, or YlAAF2 caused filamentation in wild-type cells (Fig. 5B). These findings suggest that YlRep1 may inhibit filamentation in part by repressing these transcription factor genes in GlcNAc medium.
YlRep1 and YlNgs1 repress a set of transcription factors and cell wall protein genes, some of which are associated with filamentation. (A) The upregulation of YlWOR4, MHY1, YlCBF1, and YlAAF2 was validated by qRT-PCR. Wild-type (WT) and Ylrep1Δ cells carrying pYL27-4NAG (PYlACT1-4NAG) were grown in YNB-GlcNAc medium. The transcription levels of these genes were determined by qRT-PCR and normalized to GAPDH. (B) Overexpression of YlWOR4, MHY1, YlCBF1, or YlAAF2 caused filamentation in wild-type cells grown in GlcNAc medium. Wild-type cells carrying pYL13 (Vec), pYL13-YlWOR4, pYL13-MHY1, pYL13-YlCBF1, or pYL13-YlAAF2 were grown in liquid YNB-GlcNAc medium for 16 h at 30°C. The indicated genes were overexpressed under the control of the strong constitutive promoter of the YlTEF1 gene. Bar, 20 µm. (C) The upregulation of YALI0F07535, YALI0E18722, YALI0C23452, YALI0A00176, YALI0F01857, YALI0E31108, and YALI0E20823 was validated by qRT-PCR. Wild-type (WT) and Ylrep1Δ cells carrying pYL27-4NAG (PYlACT1-4NAG) were grown in YNB-GlcNAc medium. The transcription levels of these genes were determined by qRT-PCR and normalized to GAPDH. (D) YlRep1 represses the adhesin-like genes YALI0E18722, YALI0C23452, and YALI0A00176. Wild-type (WT) and Ylrep1Δ cells carrying pYL25-4NAG (PYlACT1-4NAG) as well as pYL14-YALI0E18722ΔC, pYL14-YALI0C23452ΔC, or pYL14-YALI0A00176ΔC were grown in YNB-GlcNAc medium and photographed for GFP fluorescence. Bar, 10 µm. The cell morphology and GFP fluorescence assays were done three times on different days. For the qRT-PCR assay, mean data ± standard deviation from three independent experiments done on different days were plotted. The unpaired two-tailed Student’s t-test was used to examine the statistical significance of the difference between two samples. Statistically significant differences are indicated by the asterisks (**, P < 0.001).*
During the yeast-to-filament transition, the cell wall undergoes extensive reorganization to support hyphal growth. This process is accompanied by the upregulation and downregulation of some cell wall protein genes. RNA-Seq analysis revealed that, among the cell wall protein genes, 15 genes were upregulated (≥2-fold, P < 0.05), whereas nine genes were downregulated in Ylrep1Δ cells (Table 2). Of these, seven genes were highly upregulated (≥5-fold, P < 0.05), whereas only one gene was highly downregulated. Interestingly, all the 15 YlRep1-repressed cell wall protein genes are also repressed by YlNgs1 (Table 3), suggesting that they are co-repressed by YlRep1 and YlNgs1. We measured the expression of seven highly upregulated genes by qRT-PCR. Consistent with the RNA-Seq data, all seven genes were markedly upregulated in Ylrep1Δ cells (Fig. 5C). Among the seven highly upregulated genes, YALI0F07535 and YALI0E31108 were upregulated by more than 10-fold in Ylrep1Δ cells. They encode proteins similar to S. cerevisiae structural cell wall proteins Tir3 and Y. lipolytica cell wall protein Cwp1, respectively. These genes may be the primary targets of YlRep1 repression.
Among the seven highly upregulated cell wall protein genes, YALI0E18722, YALI0C23452, and YALI0A00176 encode proteins similar to S. cerevisiae adhesin Flo11 (Table 2). The three adhesin-like genes were upregulated by more than 15-fold and displayed high read counts in Ylrep1Δ cells. YALI0C23452 and YALI0A00176 are also highly upregulated during alkaline-induced filamentation (33). We tagged the three adhesin-like genes at the 3′-terminus of ORF with GFP (the short C-terminus containing the GPI modification site was deleted from each protein) and detected these proteins in live cells. Ylrep1Δ cells carrying three GFP-tagged genes all exhibited brighter GFP fluorescence than wild-type cells carrying the same construct (Fig. 5D). This result suggests that the three adhesin-like genes are targets of YlRep1 repression. For unclear reasons, the GFP fluorescence was predominantly retained intracellularly instead of being localized on the cell surface. This was likely due to the recycling of cell surface proteins into the vacuoles.
It should be noted that Ylrep1Δ and Ylngs1Δ cells carrying P_YlACT1_-4NAG grew slower than wild-type cells carrying P_YlACT1_-4NAG or empty vector in liquid GlcNAc medium. This may have an impact on gene expression. Together, our results suggest that YlRep1 and YlNgs1 co-repress a set of transcription factors and cell wall protein genes, some of which are associated with filamentation.
GlcNAc catabolism is not required for GlcNAc inhibition of filamentation
In C. albicans, Cahxk1Δ and Cahxk1Δ Cadac1Δ Canag1Δ cells are unable to catabolize GlcNAc but can still be induced to form hyphae, suggesting that GlcNAc catabolism is not required for the induction of filamentation by GlcNAc (27). To determine whether GlcNAc catabolism is required for the inhibition of filamentation by GlcNAc in Y. lipolytica, we examined the morphology of Ylngt1Δ and Ylnag5Δ cells (YlNag5 is the ortholog of C. albicans CaHxk1) grown in medium containing both glucose and GlcNAc (note: Yldac1Δ and Ylnag1Δ cells did not grow in this medium). In contrast to wild-type cells that did not form any filaments, Ylngt1Δ and Ylnag5Δ cells formed wrinkled colonies with short radial filaments on solid medium and were elongated and formed filaments in liquid media (Fig. 6A). YlNgt1 is the GlcNAc transporter on the cell surface. Since Ylngt1Δ cells are strongly defective in growth in GlcNAc medium (Fig. 1B), the growth seen here is most likely supported by glucose. This result suggests that GlcNAc needs to enter the cells efficiently to inhibit filamentation. The observation that Ylnag5Δ cells still formed filaments in this medium suggests that GlcNAc catabolism might be required for the inhibition of filamentation by GlcNAc.
GlcNAc catabolism is not required for the inhibition of filamentation by GlcNAc. (A) Colony and cell morphologies of Ylngt1Δ and Ylnag5Δ strains in Glucose + GlcNAc medium. Wild-type (WT), Ylngt1Δ, and Ylnag5Δ cells carrying pINA445 (Vec) or pINA445-Gene were grown on YNB-Glucose + GlcNAc agar for 3 days (upper row) or 2 days (middle row) or in liquid medium for 16 h (bottom row) at 30°C. (B) GlcNAc inhibits filamentation in Yldac1Δ and Ylnag1Δ cells. Wild-type (WT), Ylngt1Δ, Ylnag5Δ, and Ylnag5Δ Yldac1Δ Ylnag1Δ cells carrying pINA445 (Vec) were grown in liquid YNB-2% Glucose + 0.2% GlcNAc medium for 16 h at 30°C. Yldac1Δ and Ylnag1Δ cells carrying pINA445 were grown in the same medium until their cell densities were comparable to that of wild-type cells (~24 h). All the assays were done at least three times on different days. Bars, 20 µm.
To make sure that this idea is correct, we wanted to examine the morphology of Yldac1Δ and Ylnag1Δ cells, which are also unable to catabolize GlcNAc. To solve the problem of growth inhibition by GlcNAc, we cultured Yldac1Δ and Ylnag1Δ cells continuously in liquid medium containing both glucose and GlcNAc to isolate growing cells. The concentration of GlcNAc was initially set at 0.05% and gradually increased to 0.2%. We eventually isolated Yldac1Δ and Ylnag1Δ cells that grew in medium containing both 2% glucose and 0.2% GlcNAc. Interestingly, Yldac1Δ and Ylnag1Δ cells were in the oval-shaped yeast form. None of the Yldac1Δ and Ylnag1Δ cells were longer than 20 µm. In contrast, Ylngt1Δ, Ylnag5Δ, and Ylnag5Δ Yldac1Δ Ylnag1Δ cells were elongated and formed filaments (Fig. 6B). Twenty-nine percent of Ylngt1Δ, 25% of Ylnag5Δ, and 27% of Ylnag5Δ Yldac1Δ Ylnag1Δ cells (n > 400) grown in this medium were longer than 20 µm. This result suggests that the inhibition of filamentation by GlcNAc requires YlNag5 but not GlcNAc catabolism.
The transcriptional repression of YlRep1 may require YlNag5
Because Ylrep1Δ cells and Ylnag5Δ cells formed filaments in liquid medium containing both glucose and GlcNAc ([Fig. 4A and 6A](#F4 F6)), this raises the possibility that YlNag5 may be required for YlRep1 to repress filamentation-related genes. To explore this possibility, we measured the expression levels of major YlRep1-repressed transcription factor and cell wall protein genes (identified by RNA-Seq) in wild-type, Ylrep1Δ, Ylnag5Δ, and Ylrep1Δ Ylnag5Δ cells grown in medium containing both glucose and GlcNAc (note: this medium is different from the GlcNAc medium used for RNA-Seq). For the four transcription factor genes, MHY1 and YlWOR4 were markedly upregulated in Ylrep1Δ cells grown in this medium. They were also significantly upregulated in Ylnag5Δ and Ylrep1Δ Ylnag5Δ cells (Fig. 7A). Although YlAAF2 and YlCBF1 were not significantly upregulated in Ylrep1Δ cells grown in this medium, they were significantly upregulated in Ylnag5Δ cells, and YlCBF1 was also significantly upregulated in Ylrep1Δ Ylnag5Δ cells (Fig. 7A). For the seven cell wall protein genes, all of them were significantly upregulated in Ylrep1Δ cells except YALI0F21857, and they all were significantly upregulated in Ylnag5Δ and Ylrep1Δ Ylnag5Δ cells (Fig. 7B). These results indicate that major YlRep1-repressed transcription factor and cell wall protein genes are all upregulated in Ylnag5Δ cells grown in medium containing both glucose and GlcNAc, suggesting that the repression of these genes by YlRep1 may require YlNag5.
The transcriptional repression of YlRep1 may require YlNag5. The major transcription factor and cell wall protein genes repressed by YlRep1 were upregulated in Ylnag5Δ cells. The transcription levels of transcription factor genes YlWOR4, MHY1, YlCBF1, YlAAF2 (A), and cell wall protein genes YALI0F07535, YALI0E18722, YALI0C23452, YALI0A00176, YALI0F01857, YALI0E31108, and YALI0E20823 (B) in wild-type (WT), Ylrep1Δ, Ylnag5Δ, and Ylrep1Δ Ylnag5Δ cells grown in YNB-Glucose + GlcNAc medium were determined by qRT-PCR and normalized to GAPDH. All qRT-PCR data showed mean ± standard deviation from three independent experiments done on different days. The unpaired two-tailed Student’s t-test was used to examine the statistical significance of the difference between two samples. Statistically significant differences are indicated by the asterisks (, P < 0.05; **, P < 0.01; **, P < 0.001). NS, not statistically significant.
DISCUSSION
GlcNAc is a common source of carbon and nitrogen in nature, present in both terrestrial and aquatic environments, as well as in animal intestines. Surprisingly, it is also a signaling molecule that stimulates morphogenetic transitions in fungi. In this study, we show that GlcNAc inhibits filamentation in the dimorphic yeast Y. lipolytica. The Rep1-Ngs1 complex not only regulates the induction of GlcNAc catabolic genes but also the inhibition of filamentation by GlcNAc. GlcNAc inhibition of filamentation requires the GlcNAc kinase YlNag5 but operates independently of GlcNAc catabolism. These results provide new insights into the GlcNAc-regulated dimorphic transition in Y. lipolytica.
GlcNAc is a potent inducer of filamentation in C. albicans (29). Based on this observation, researchers studying dimorphic transition in Y. lipolytica also tested GlcNAc for its ability to induce filamentation, often using it in combination with a citrate buffer at near-neutral pH (pH 6–7) (see a summary in Table S3) (4, 30, 31). Sometimes, additional treatment of the cells, such as brief cold treatment, was also employed to increase the efficiency (4, 31). Since this combination proved effective, many papers that cited the results have described GlcNAc as an inducer of filamentation in Y. lipolytica. However, because citrate and particularly neutral pH are strong inducers of filamentation (5), the exact role of GlcNAc in regulating filamentation in Y. lipolytica remains unclear. One study reported that GlcNAc alone induces filamentation poorly compared to glucose and suggested that the apparent “filament-inducing” effect of GlcNAc might actually be due to neutral pH and citrate (see Table S3) (5). Our findings support this idea. While GlcNAc inhibits filamentation, we observed that cells grown in GlcNAc were still able to form filaments in response to alkaline pH induction, in agreement with previous reports. Similarly, GlcNAc has been shown to strongly inhibit filamentation in C. tropicalis (25), which is closely related to C. albicans. However, the mechanism underlying this inhibition is not clear.
In C. albicans, GlcNAc induces filamentation via both alkalization of the culture medium and CaNgs1 signaling (19, 21, 32). In contrast, GlcNAc inhibits filamentation primarily via YlRep1-YlNgs1 signaling in Y. lipolytica. Although GlcNAc catabolism slightly decreases ambient pH, it is less strong than glucose catabolism. Based on the observation that glucose induces filamentation even though glucose catabolism decreases ambient pH, the decrease of ambient pH by GlcNAc catabolism is not critical for the inhibition of filamentation.
Why does GlcNAc promote filamentation in C. albicans but inhibit filamentation in Y. lipolytica and C. tropicalis? These opposing, species-specific responses likely reflect the distinct ecological niches and evolutionary pressures faced by these species. C. albicans, which typically resides within the human body, uses filamentation as a strategy to invade host tissues more effectively and evade immune responses (8). Since GlcNAc is naturally present in the host environment as a component of the extracellular matrix of host cells and a cell wall component of commensal microbiota, it likely acts as a signal to C. albicans that it is in a host environment, triggering filamentation as a survival and invasive strategy. In contrast, Y. lipolytica and C. tropicalis primarily inhabit soil and water (34–36). GlcNAc may act more as a nutrient source than a morphogenic signal for filamentation in Y. lipolytica and C. tropicalis.
The Ndt80 family transcription factor Rep1 (called Ron1 in filamentous fungi) is essential for GlcNAc utilization and the induction of GlcNAc catabolic genes in yeast species C. albicans, C. tropicalis, and Y. lipolytica, as well as in filamentous fungi, such as T. reesei (19, 23, 26). The histone acetyltransferase Ngs1 is widely conserved across fungal species that can metabolize GlcNAc, where Ngs1 functions as the GlcNAc sensor and transducer (19, 24). Several studies, including this one, have shown that the Rep1-Ngs1 complex plays a critical role in inducing GlcNAc catabolic genes in various fungal species (19, 24, 37). These results suggest that the role of Rep1-Ngs1 in regulating GlcNAc catabolic genes might be evolutionarily conserved.
In C. albicans, the induction of GlcNAc catabolic genes requires CaRep1-CaNgs1 signaling, whereas GlcNAc-induced filamentation requires CaNgs1 but not CaRep1 (19, 21). CaNgs1 is thought to bind to the promoter of the transcription factor CaBrg1 and induce its expression in the presence of GlcNAc. CaBrg1, in turn, downregulates the expression of CaNrg1, a major repressor of filamentation, leading to hyphal development (19). Notably, this regulation is independent of GlcNAc catabolism. In the human pathogen C. tropicalis, GlcNAc strongly inhibits filamentation (25), which is similar to the regulation observed in Y. lipolytica. Interestingly, CtRep1 is also required for the inhibition of filamentation by GlcNAc, as filamentation occurred in Ctrep1Δ cells but not in wild-type cells grown in medium containing both glucose and GlcNAc (26). This finding closely parallels our observations in Y. lipolytica. In this study, we demonstrate that both YlRep1 and YlNgs1 are essential for GlcNAc inhibition of filamentation in Y. lipolytica. YlRep1 likely functions by recruiting YlNgs1 to the promoters of target genes. Together, they repress filamentation-related genes, including the key regulator of filamentation MHY1 (which encodes a transcription factor) and three adhesin-like genes.
Our results indicate that the YlRep1-YlNgs1 complex plays dual and opposing roles in regulating gene expression. In response to GlcNAc, it activates GlcNAc catabolic genes but represses filamentation-related genes. In C. tropicalis, where GlcNAc also inhibits filamentation, although it is not yet known about CtNgs1’s role, CtRep1 also activates GlcNAc catabolic genes but inhibits filamentation (26), which resembles Y. lipolytica YlRep1. In C. albicans, where GlcNAc induces filamentation, although CaRep1 is not involved in regulating GlcNAc-induced filamentation, CaRep1 is initially reported to repress the expression of MDR1 efflux pump involved in drug resistance (20), indicating that CaRep1 also plays dual and opposing roles in regulating gene expression. In this study, we show that YlRep1 and YlNgs1 exhibit transcriptional activation activity, which could explain how the Rep1-Ngs1 complex activates GlcNAc catabolic genes in response to GlcNAc. However, it remains unclear how YlRep1-YlNgs1 represses gene expression.
In this study, we observed that GlcNAc failed to inhibit filamentation in Ylnag5Δ and Ylnag5Δ Yldac1Δ Ylnag1Δ cells grown in medium containing both glucose and GlcNAc, suggesting that YlNag5 is important for the inhibition of filamentation by GlcNAc. Supporting this finding, we observed that major transcription factor and cell wall protein genes repressed by YlRep1 were upregulated in Ylnag5Δ cells grown in medium containing both glucose and GlcNAc, indicating that YlNag5 may be required for YlRep1 to repress filamentation-related genes. However, the exact mechanism underlying this YlNag5 function is unclear.
In C. tropicalis, where GlcNAc also inhibits filamentation, GlcNAc still inhibited filamentation in Cthxk1Δ cells (CtHxk1 is the ortholog of YlNag5) while CtHxk1 overexpression drastically increased filamentous growth on Lee’s media containing GlcNAc (38), suggesting that CtHxk1 promotes filamentation rather than inhibiting it. Interestingly, a previous report indicated that overexpression of YlNag5 led to strong filamentation in Y. lipolytica cells grown in media containing glucose, glycerol, or GlcNAc (14), suggesting that YlNag5 may function similarly to CtHxk1. However, in our study, we did not observe a similar effect when YlNag5 was overexpressed.
Our study characterizes the inhibitory effect of GlcNAc on filamentous growth in Y. lipolytica and identifies YlRep1-YlNgs1 as a key regulator in the inhibition of filamentation by GlcNAc. It also suggests that the repression of filamentation-related genes by YlRep1 may require the GlcNAc kinase YlNag5. However, there are also limitations in this study. For example, some of the mutants generated in this study, such as Ylrep1Δ and Ylngs1Δ cells carrying P_YlACT1_-4NAG, grew slower than wild-type cells carrying P_YlACT1_-4NAG or empty vector in liquid GlcNAc medium. Moreover, Yldac1Δ and Ylnag1Δ cells also grew slower than wild-type cells in medium containing 2% glucose and 0.2% GlcNAc. The slow-growth phenotype of these mutants may have an impact on gene expression and filamentation. We observed that YlRep1-YlNgs1 exhibits opposing effects on filamentation compared to C. albicans CaNgs1. It is not clear what factor may determine the divergent outcomes between these two yeast species. It is also not clear why GlcNAc fails to inhibit filamentation in Ylnag5Δ and Ylnag5Δ Yldac1Δ Ylnag1Δ cells. Future investigations are needed to address these questions.
MATERIALS AND METHODS
Strains and media
The Y. lipolytica strains used in this study are listed in Table S4 in the supplemental material. PO1a (MATA leu2-270 ura3-302) was used as the wild-type strain. Y. lipolytica strains were grown at 30°C. Culture media include yeast extract-peptone-dextrose (YPD) medium (20 g/L peptone, 10 g/L yeast extract, 2% glucose) and YNB medium (6.7 g/L yeast nitrogen base without amino acid) containing GlcNAc (1%), glucose (1%), glycerol (1%), or glucose + GlcNAc (1% glucose, 1% GlcNAc) supplemented with 80 mg/L leucine, 20 mg/L uracil, or both, when required. Na_2_HPO_4_-citric acid buffer was used to adjust the medium to pH 7.0 or pH 7.5 when required. S. cerevisiae strains used are listed in Table S4. E. coli strain DH5α was used for plasmid amplification.
Plasmid construction
The plasmids used in this study are listed in Table S5. The oligonucleotide primers are listed in Table S6. To generate pINA445-YlREP1, YlREP1 carrying a 2,000 bp promoter and 400 bp 3′-UTR was amplified from genomic DNA by PCR and inserted into HindIII-digested vector pINA445 (CEN, LEU2) using ClonExpress II One Step Cloning Kit (Vazyme Biotech Co., China). Similarly, YlNGS1 (carrying 1,400 bp promoter and 400 bp 3′-UTR), YlNDT80 (carrying 500 bp promoter and 500 bp 3′-UTR), YlNGT1 (carrying 2,000 bp promoter and 500 bp 3′-UTR), YlNAG5 (carrying 841 bp promoter and 500 bp 3′-UTR), YlDAC1 (carrying 2,000 bp promoter and 517 bp 3′-UTR), and YlNAG1 (carrying 2,000 bp promoter and 500 bp 3′-UTR) were inserted into pINA445, yielding pINA445-YlNGS1, pINA445-YlNDT80, pINA445-YlNGT1, pINA445-YlNAG5, pINA445-YlDAC1, and pINA445-YlNAG1, respectively.
To generate the plasmid that carries four GlcNAc catabolic genes (NAG) under the control of YlACT1 promoter, the 947 bp YlACT1 promoter (nucleotides −950 to −4 relative to the first nucleotide in the start codon) was amplified by PCR from genomic DNA and fused to the ORFs of four GlcNAc catabolic genes by overlapping PCR. The resulting expression cassettes P_YlACT1_-YlNGT1, P_YlACT1_-YlNAG5, P_YlACT1_-YlDAC1, and P_YlACT1_-YlNAG1 were inserted into pYL25 (CEN, YlURA3) and pYL27 (CEN, YlLEU2), yielding pYL25-4NAG and pYL27-4NAG, respectively.
To monitor the transcriptional activities of YlNGT1, YlNAG5, YlDAC1, and YlNAG1 promoters in Ylrep1Δ cells and Ylngs1Δ cells versus wild-type cells, the promoter region of each gene plus the ATG start codon was amplified by PCR and inserted into pINA445-lacZ using the ClonExpress II One-Step Cloning Kit, yielding pINA445-P_YlNGT1_-lacZ, pINA445-P_YlNAG5_-lacZ, pINA445-P_YlDAC1_-lacZ, and pINA445-P_YlNAG1_-lacZ.
To generate LexA-YlRep1 and LexA-YlNgs1 for one-hybrid assay, the DNA-binding domain (a.a. 1-87) of the E. coli lexA gene was amplified and inserted into HindIII-digested pYL21, yielding pYL21-lexA. Similarly, lexA-YlREP1 and lexA-YlNGS1 were generated by overlapping PCR and inserted into pYL21, yielding pYL21-lexA-YlREP1 and pYL21-lexA-YlNGS1. pINA445-lexAop-P_YlLEU2_-lacZ was reported previously (39).
To overexpress the transcription factor genes, the ORFs of YlWOR4, YlCBF1, MHY1, and YlAAF2 plus 300 bp 3′-UTR were amplified by PCR and inserted into pYL13 (CEN, YlLEU2, P_YlTEF1_), yielding pYL13-YlWOR4, pYL13-YlCBF1, pYL13-MHY1, and pYL13-YlAAF2, respectively.
To visualize the transcriptional activities of the adhesin-like genes that were highly repressed by YlRep1, YALI0C23452^1-794^ carrying a 2,674 bp promoter, YALI0E18722^1-608^ carrying a 1,775 bp promoter, and YALI0A00176^1-825^ carrying a 2,000 bp promoter were amplified by PCR and inserted into BamHI-digested pYL14, yielding pYL14-YALI0C23452^ΔC^, pYL14-YALI0E18722^ΔC^, and pYL14-YALI0A00176^ΔC^, respectively.
For the two-hybrid assay, full-length YlNGS1 ORF was amplified from genomic DNA by PCR and inserted into BamHI-digested pGAD-C1 (2μ, LEU2, GAL4-AD) and pGBDU-C1 (2μ, URA3, GAL4-DBD), yielding pGAD-YlNGS1 and pGBDU-YlNGS1, respectively. Similarly, full-length YlREP1, YlREP1^1-200^, YlREP1^1-376^, and YlREP1^377-494^ segments were amplified from genomic DNA by PCR and inserted into BamHI-digested pGAD-C1 and pGBDU-C1, yielding pGAD-YlREP1, pGAD-YlREP1 segments, pGBDU-YlREP1 and pGBDU-YlREP1 segments.
Yeast strain construction
YlNGT1, YlNAG5, YlDAC1, YlNAG1, YlREP1, YlNGS1, and YlNDT80 were deleted in Y. lipolytica strains by homologous recombination following standard procedure (39).
β-Galactosidase assay
The β-galactosidase activity in the cells was determined by the crude cell extract assay using O-nitrophenyl-β-D-galactopyranoside (ONPG) as the substrate, as reported previously (39).
Yeast two-hybrid assay
pGAD-C1-based plasmids were transformed into the S. cerevisiae haploid strain pJ69-4A. pGBDU-C1-based plasmids were transformed into the haploid strain pJ69-4α. Pairs of haploid strains were mated on YPD plates and then replica plated onto SC-Leu-Ura plates to select for diploid cells that harbor both bait and prey plasmids. Diploid cells were patched on an SC-Leu-Ura plate and replica plated onto an SC-Leu-Ura-Ade plate to check for growth. Growth indicates interaction between the DNA-binding domain (DBD) and activation domain (AD) fusion proteins.
RNA-Seq analysis
Y. lipolytica cells were grown in liquid YNB-GlcNAc medium supplemented with uracil at 30°C and collected at an OD600 of ~0.8. Each sample was in triplicate. RNA-Seq was conducted at Personalbio (Shanghai, China). Differential expression analysis between two conditions was performed on Personalbio Genescloud. Differentially expressed genes were defined as those for which the adjusted P value <0.05 and the fold change ≥2.0.
RNA extraction and quantitative real-time PCR analysis
Yeast cells grown in 50 mL of culture at 30°C were collected at an OD_600_ of ~0.8. Total RNAs were extracted using Yeast RNA Kit (Omega, China). RNA integrity was examined by 1% agarose gel electrophoresis. The yields of RNAs were examined by the NanoDrop One Spectrometer. One microgram total RNA per sample was used for reverse transcription using the HiScript II Q RT SuperMix for qPCR (Vazyme Biotech Co., China). qPCRs were carried out using the ChamQ Universal SYBR qPCR Master (Vazyme Biotech Co., China) following the manufacturer’s instructions. The primers of qPCR were exquisitely selected to ensure only one peak per primer set, and the Cq values of all amplification curves were in the range from 15 to 30. Each sample was in triplicate and analyzed using Bio-Rad CFX Maestro (version 1.1) with normalized mode (ΔΔCq).
Microscopy
An Olympus BX51 microscope (Tokyo) and a Retiga 2000R CCD camera (QImaging Corporation) were used to visualize cell morphology and green fluorescent protein. The images were acquired using QCapture Suite (QImaging Corporation).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dujon B, Sherman D, Fischer G, Durrens P, Casaregola S, Lafontaine I, De Montigny J, Marck C, Neuvéglise C, Talla E, et al.. 2004. Genome evolution in yeasts. Nature 430:35–44. doi:10.1038/nature 0257915229592 · doi ↗ · pubmed ↗
- 2Groenewald M, Boekhout T, Neuvéglise C, Gaillardin C, van Dijck PWM, Wyss M. 2014. Yarrowia lipolytica: safety assessment of an oleaginous yeast with a great industrial potential. Crit Rev Microbiol 40:187–206. doi:10.3109/1040841 X.2013.77038623488872 · doi ↗ · pubmed ↗
- 3Miller KK, Alper HS. 2019. Yarrowia lipolytica: more than an oleaginous workhorse. Appl Microbiol Biotechnol 103:9251–9262. doi:10.1007/s 00253-019-10200-x 31686142 · doi ↗ · pubmed ↗
- 4Pérez-Campo FM, Domínguez A. 2001. Factors affecting the morphogenetic switch in Yarrowia lipolytica. Curr Microbiol 43:429–433. doi:10.1007/s 00284001033311685511 · doi ↗ · pubmed ↗
- 5Ruiz-Herrera J, Sentandreu R. 2002. Different effectors of dimorphism in Yarrowia lipolytica. Arch Microbiol 178:477–483. doi:10.1007/s 00203-002-0478-312420169 · doi ↗ · pubmed ↗
- 6Cullen PJ, Sprague GF Jr. 2012. The regulation of filamentous growth in yeast. Genetics 190:23–49. doi:10.1534/genetics.111.12745622219507 PMC 3249369 · doi ↗ · pubmed ↗
- 7Lengeler KB, Davidson RC, D’souza C, Harashima T, Shen WC, Wang P, Pan X, Waugh M, Heitman J. 2000. Signal transduction cascades regulating fungal development and virulence. Microbiol Mol Biol Rev 64:746–785. doi:10.1128/MMBR.64.4.746-785.200011104818 PMC 99013 · doi ↗ · pubmed ↗
- 8Berman J, Sudbery PE. 2002. Candida albicans: a molecular revolution built on lessons from budding yeast. Nat Rev Genet 3:918–930. doi:10.1038/nrg 94812459722 · doi ↗ · pubmed ↗