The centromeric gene OsDCL plays essential roles in plant development and yield production in rice
Xiangbo Li, Chao Xue, Rui Guo, Zhiying Wang, Kai Liu, Mingchen Shen, Lu Ye, Jijin Chen, Hanli You, Jun Miao, Liang Chen, Wenhui Bian, Yong Zhou, Zhiyun Gong

TL;DR
The OsDCL gene in rice is crucial for plant development and increasing yield by improving photosynthesis.
Contribution
OsDCL is identified as a centromeric gene that enhances rice yield through chlorophyll synthesis and photosynthesis regulation.
Findings
OsDCL regulates chlorophyll synthesis in rice.
The gene enhances photosynthesis and contributes to higher yield.
It is a key genetic resource for breeding high-yield rice.
Abstract
The rice centromeric gene OsDCL boosts yield by regulating chlorophyll synthesis and photosynthesis, making it a key genetic resource for breeding high-yield rice with superior photosynthetic efficiency.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
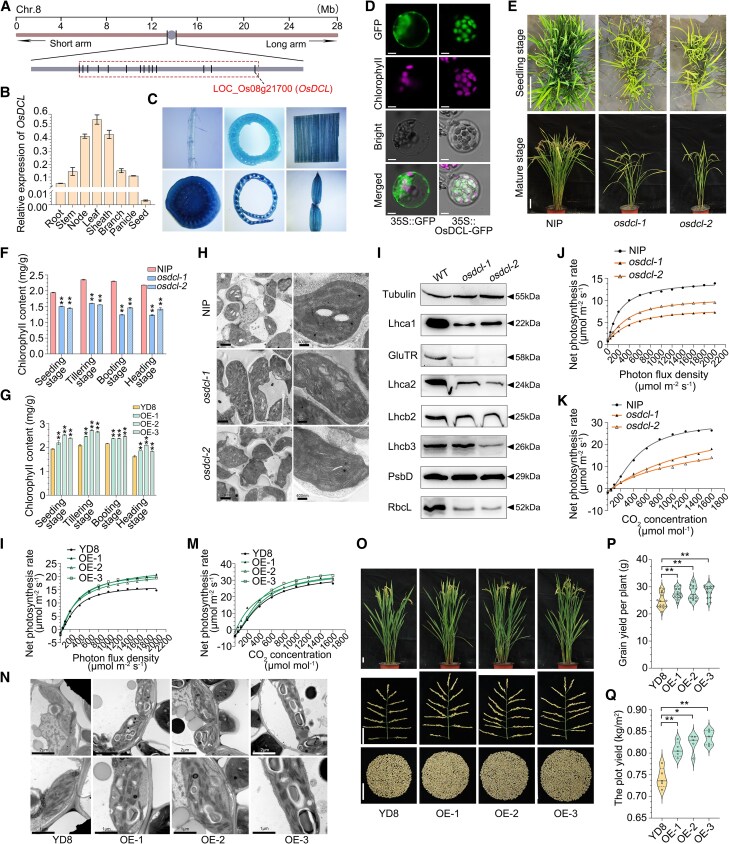 Figure 1
Figure 1- —National Key Research and Development Program of China10.13039/501100012166
- —Jiangsu Province Government
- —Natural Science Foundation of Jiangsu Province10.13039/501100004608
- —National Natural Science Foundation of China10.13039/501100001809
- —Post-graduate Research and Practice Innovation Program of Jiangsu Province
- —Jiangsu Higher Education Institutions
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Photosynthetic Processes and Mechanisms · Genetic Mapping and Diversity in Plants and Animals
Dear Editor,
Active genes exist in the euchromatic regions of centromeres across several species (Nagaki et al. 2004; Wu et al. 2011; Lian et al. 2024). Until now, there have been limited studies addressing the functions of the active genes located in the centromeric region of plants. Previously, we identified several putative active genes within the centromeric region of rice (Oryza sativa L.) chromosome 8 (Xue et al. 2022). Among them, LOC_Os08g21700 encodes a chloroplast precursor, homologous to the chloroplast and leaf defective (DCL) gene of tomato and the AtDCL gene of Arabidopsis, and is named OsDCL (Fig. 1A). In tomato, DCL is essential for plastid ribosomal RNA and functions at an early stage of chloroplast development (Keddie et al. 1996; Bellaoui et al. 2003). In Arabidopsis, aberrant expression of the AtDCL results in yellowing leaves, sterile flowers, anomalous cotyledons, and even death (Bellaoui and Gruissem 2004). Nevertheless, the functional roles of OsDCL in rice remain unelucidated.
We initially investigated the expression pattern of OsDCL. RT-qPCR analysis showed that OsDCL was constitutively expressed in all tested tissues, with the highest expression in leaf, sheath, and node, and the lowest expression in seed (Fig. 1B). Promoter-driven GUS reporter analysis showed a largely similar expression pattern (Fig. 1C). Transient expression of the 35S::OsDCL-GFP construct in rice protoplasts demonstrated that OsDCL was localized in the chloroplasts (Fig. 1D). To further uncover the biological function of OsDCL in rice, we utilized CRISPR/Cas9 technology to generate knockout mutants of OsDCL in the Nipponbare (NIP) background (see Supplementary Materials and Methods). Two homozygous mutants (named osdcl-1 and osdcl-2), carrying a single base pair deletion and a single base pair insertion in the target sequence (Supplementary Fig. S1), respectively, were subjected to detailed analysis. Throughout the entire growth period, the osdcl-1 and osdcl-2 mutants exhibited a yellow–green leaf phenotype (Fig. 1E). The mutants showed significantly reduced chlorophyll contents at seedling, tillering, booting, and heading stages (Fig. 1F). We conducted RNA-seq analysis using 2-week-old seedlings of osdcl-1 and NIP. Compared with NIP, the osdcl-1 mutant displayed significant alterations in gene expression profile, comprising 1,577 upregulated and 1,506 downregulated genes (Supplementary Fig. S2A). Gene ontology enrichment analysis showed that the 1,506 downregulated genes were significantly enriched in cellular component classes associated with chloroplast, plastid and thylakoid parts (Supplementary Fig. S2B). Furthermore, the expression of several key genes in chlorophyll synthesis, photosynthesis, and chloroplast development pathway were decreased in the osdcl mutants (Supplementary Fig. S3A). Transmission electron microscopy (TEM) showed that knockout of OsDCL led to abnormally developed chloroplasts, characterized by disorganized thylakoid membrane structure and abnormal grana configurations (Fig. 1H). Western-blot analysis indicated a substantial reduction in the abundance of several critical photosynthetic proteins, specifically Lhca1, GluTR, Lhca2, Lhcb3, and RbcL, in the osdcl mutants (Fig. 1I). Because chlorophyll content and chloroplast development are closely related to rice photosynthesis (Wang et al. 2015; Zhang et al. 2023), we compared the photosynthetic rates between NIP and the osdcl mutants. Analysis of the photosynthetic light response and CO_2_ response curves demonstrated that the net photosynthetic rates of osdcl-1 and osdcl-2 were substantially lower than that of NIP (Fig. 1J, K). From the seedling stage onward, the osdcl mutants displayed pronounced growth inhibition, manifested primarily by reduced tiller formation (Supplementary Fig. S4A, B). At the mature stage, the osdcl mutants exhibited significantly reduced grain size, plant height, panicle number, grain number per panicle, and grain yield per plant compared to NIP (Supplementary Fig. S4C-L). These results indicate that the loss-of-function of OsDCL seriously impacts chloroplast development and photosynthetic process, subsequently resulting in a substantial hindrance in plant growth and a consequential decrease in yield.
To further evaluate the genetic effects of OsDCL in rice, we constructed transgenic lines overexpressing OsDCL in the japonica variety Yandao8 (YD8) background, which is a high-yield variety that was once widely cultivated. Three independent overexpression lines (OE-1, OE-2, and OE-3) with significantly increased OsDCL expression levels were used for further analysis (Supplementary Fig. S5). Compared to YD8, chlorophyll content in the leaves of the three OE lines increased by 10.0% to 13.4% at seedling, tillering, booting and heading stages (Fig. 1G). The expression of several key genes in chlorophyll synthesis, photosynthesis, and chloroplast development pathway were increased in the OE lines (Supplementary Fig. S3B). Additionally, the net photosynthetic rates were obviously increased in the three OE lines (Fig. 1L, M). TEM analysis showed that the chloroplasts of the OE lines contained regularly distributed grana (Fig. 1N).
In field conditions, the three OE lines demonstrated greater growth potential. Compared to YD8, the OE lines had more tillers (Supplementary Fig. S6A, B). While the OE lines showed significantly increased grain length and width compared to the wild-type, their grain thickness was reduced, accounting for the observed decrease in grain weight (Supplementary Fig. S6C-H). Compared to YD8, the plant height, panicles number per plant and grain number per panicle of the OE lines were also significantly increased (Fig. 1O; Supplementary Fig. S6I-K). At maturity, compared to YD8, the OsDCL overexpression lines showed increased biomass yield per plant (Supplementary Fig. S6L). Moreover, grain yield per plant of the three OE lines increased by 10.6% to 13.5% (Fig. 1O, P). These findings were corroborated by field plot trials, where the three OE lines demonstrated yield increases of 8.8%, 10.9%, and 12.6% per plot, respectively (Fig. 1Q). However, three OE lines exhibited reduced appearance quality (Supplementary Fig. S7). These data indicate that overexpression of OsDCL promotes plant growth and significantly increases grain yield production in rice.
To date, functional studies on actively transcribed genes within rice centromeric regions have made limited progress (Xie et al. 2023). In this study, we characterized OsDCL as a functionally essential gene residing within the centromeric region of chromosome 8 in rice. Our study demonstrates that OsDCL mutations induce severe chloroplast ultrastructural defects, leading to reduced chlorophyll content and impaired photosynthetic capability, ultimately stunting plant growth. Conversely, overexpression of OsDCL elevates chlorophyll accumulation, enhances photosynthetic performance, and boosts grain yield. These findings provide evidence that transcriptionally active genes in centromeric regions can play functional roles, suggesting that centromeres may not be entirely genetically silent. Given the centromeric localization of OsDCL, future investigations should systematically examine whether its regulatory mechanisms exhibit fundamental divergence from those governing euchromatic gene expression.
Supplementary Material
kiaf500_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bellaoui M, Gruissem W. Altered expression of the Arabidopsis ortholog of DCL affects normal plant development. Planta. 2004:219(5):819–826. 10.1007/s 00425-004-1295-515197595 · doi ↗ · pubmed ↗
- 2Bellaoui M, Keddie JS, Gruissem W. DCL is a plant-specific protein required for plastid ribosomal RNA processing and embryo development. Plant Mol Biol. 2003:53(4):531–543. 10.1023/B:PLAN.0000019061.79773.0615010617 · doi ↗ · pubmed ↗
- 3Keddie JS, Keddie JS, Carroll B, Jones JD, Gruissem W. The DCL gene of tomato is required for chloroplast development and palisade cell morphogenesis in leaves. EMBO J. 1996:15(16):4208–4217. 10.1002/j.1460-2075.1996.tb 00795.x 8861949 PMC 452145 · doi ↗ · pubmed ↗
- 4Lian QC, Huettel B, Walkemeier B, Mayjonade B, Lopez-Roques C, Gil L, Roux F, Schneeberger K, Mercier R. A pan-genome of 69 Arabidopsis thaliana accessions reveals a conserved genome structure throughout the global species range. Nat Genet. 2024:56(5):982–991. 10.1038/s 41588-024-01715-938605175 PMC 11096106 · doi ↗ · pubmed ↗
- 5Nagaki K, Cheng Z, Ouyang S, Talbert PB, Kim M, Jones KM, Henikoff S, Buell CR, Jiang J. Sequencing of a rice centromere uncovers active genes. Nat Genet. 2004:36(2):138–145. 10.1038/ng 128914716315 · doi ↗ · pubmed ↗
- 6Wang Q, Xie W, Xing H, Yan J, Meng X, Li X, Fu X, Xu J, Lian X, Yu S, et al Genetic architecture of natural variation in rice chlorophyll content revealed by a genome-wide association study. Mol Plant. 2015:8(6):946–957. 10.1016/j.molp.2015.02.01425747843 · doi ↗ · pubmed ↗
- 7Wu YF, Kikuchi S, Yan H, Zhang W, Rosenbaum H, Iniguez AL, Jiang J. Euchromatic subdomains in rice centromeres are associated with genes and transcription. Plant Cell. 2011:23(11):4054–4064. 10.1105/tpc.111.09004322080597 PMC 3246336 · doi ↗ · pubmed ↗
- 8Xie E, Chen JW, Wang BX, Shen Y, Tang D, Du GJ, Li YF, Cheng ZK. The transcribed centromeric gene Os MRPL 15 is essential for pollen development in rice. Plant Physiol. 2023:192(2):1063–1079. 10.1093/plphys/kiad 15336905369 PMC 10231452 · doi ↗ · pubmed ↗