Under control: A WRKY transcription factor positively regulates the expression of a helper NLR immune receptor in Nicotiana benthamiana
Josephine H R Maidment

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
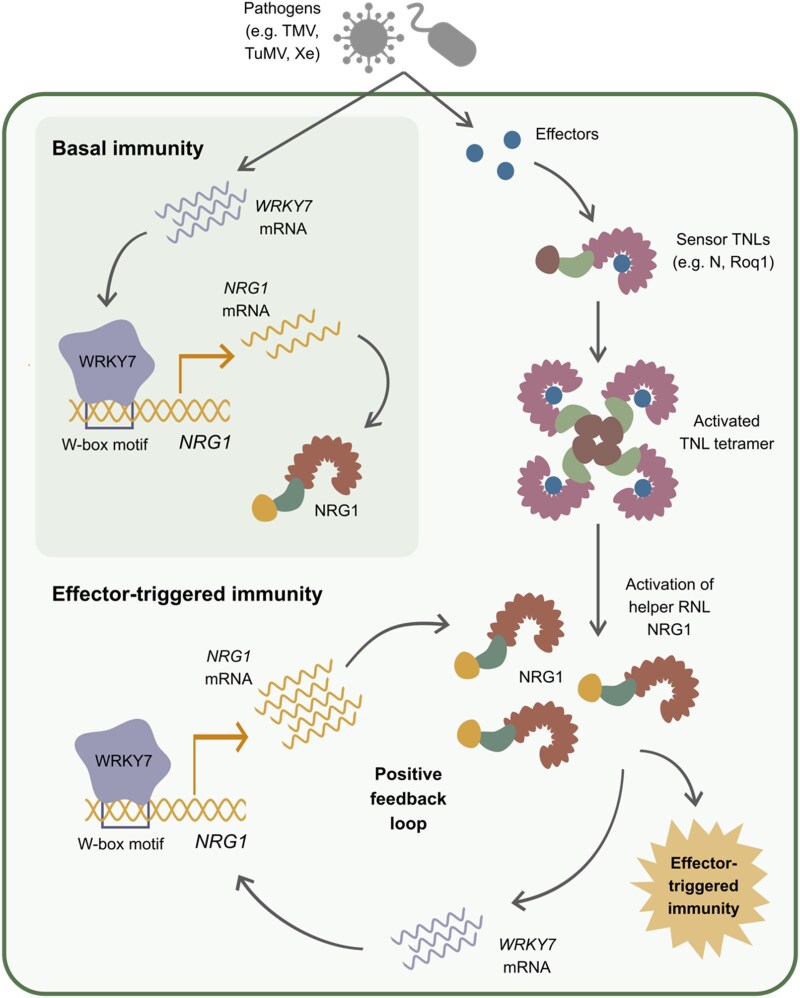 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsToxin Mechanisms and Immunotoxins · Transgenic Plants and Applications
To defend themselves against pathogens, plants have an innate immune system that can be conceptually divided into 2 layers (reviewed by Bentham et al. 2020). Cell surface receptors recognize molecular signatures of pathogen attack in the extracellular space, while cytoplasmic nucleotide-binding leucine-rich repeat (NLR) receptor proteins detect molecules, known as effectors, which are secreted by the pathogen into the plant cell to promote infection. Together, these 2 classes of receptors coordinate defense responses to restrict the spread and impact of a pathogen. Basal immune responses, predominantly conditioned by cell surface receptors, include production of reactive oxygen species, callose deposition, synthesis of antimicrobial compounds, and upregulation of defense genes. By contrast, effector-triggered immunity frequently culminates in localized cell death, known as the hypersensitive response. This extreme outcome serves to restrict the spread of a pathogen to neighboring cells.
In some cases, a single NLR protein can detect an effector and activate immunity. However, certain NLRs have become specialized as effector sensors and require helper NLRs to trigger plant defenses (reviewed by Contreras et al. 2023). One such helper NLR is NRG1, which is required by a subset of sensor NLR proteins containing an N-terminal TIR domain (TNLs) to induce immune responses following effector recognition. Structural and biochemical studies have elucidated the molecular detail of how effector recognition by a sensor TNL leads to activation and oligomerization of NRG1 (Lapin et al. 2019; Jacob et al. 2021; Jia et al. 2022; Ibrahim et al. 2024). Interestingly, NRG1 oligomerization requires input from cell surface receptors (Feehan et al. 2023), highlighting that NRG1 is an important node integrating inputs from both layers of the plant immune system. However, while our mechanistic understanding of NRG1 activation has advanced in recent years, comparatively little is known about the transcriptional regulation of NRG1.
WRKY transcription factors (TFs) are abundant in plant genomes. They are so named for a signature WRKYGQK motif found in their DNA binding domain, which binds conserved W-box cis elements in promoters of target genes to modulate gene expression. WRKY TFs have regulatory roles in plant development as well as abiotic and biotic stress responses. Several have well-defined functions in regulating expression of defense-related genes and influencing disease outcomes (reviewed by Javed and Gao 2023).
In a recent study published in Plant Physiology, Wu et al. (2025) identified a WRKY TF, WRKY7, which regulates the expression of NRG1 in Nicotiana benthamiana. Wu et al. performed DNA pulldown assays with the NRG1 promoter to identify candidate transcriptional regulators. Transient overexpression of candidates in N. benthamiana subsequently identified 7 proteins that limited the spread of turnip mosaic virus, suggesting that these proteins are positive regulators of plant immunity. Upon observing that the NRG1 promoter contained W-box motifs, the authors selected one of these proteins, WRKY7, for further investigation.
Using electrophoretic mobility shift and yeast 1-hybrid assays, the authors demonstrated direct binding of WRKY7 to the promoter of NRG1. In addition, they investigated the role of WRKY7 in modulating NRG1 expression in vivo using a transient dual luciferase reporter assay, with firefly luciferase expression driven by the NRG1 promoter. Overexpression of WRKY7 increased firefly luciferase activity, but mutation of the W-box motifs in the NRG1 promoter abolished this increase, supporting the conclusion that WRKY7 binds W-box motifs in the NRG1 promoter to enhance NRG1 expression.
After observing that infection by either tobacco mosaic virus (TMV) or turnip mosaic virus induced WRKY7 and NRG1 expression but without activating cell death, Wu et al. investigated the effect of WRKY7 on basal immunity. Silencing WRKY7 led to a reduction in NRG1 transcription and to an increase in susceptibility to GFP-TMV, evidenced by increased viral coat protein levels as compared with wild type (WT) plants. This indicates that WRKY7 has a role in modulating basal immunity, potentially through regulation of NRG1 expression.
To explore the contribution of WRKY7 to effector-triggered immunity mediated by the TNLs N (which recognizes the TMV replicase p50) and Roq1 (which detects the effector XopQ secreted by the bacterial pathogen Xanthomonas euvesicatoria), Wu et al. silenced WRKY7 in N. benthamiana plants carrying N or Roq1. Following transient expression of p50 or XopQ, reduced cell death, indicating a reduced immune response, was observed in WRKY7-silenced plants as compared with WT plants. WRKY7-silenced plants carrying N or Roq1 were more susceptible to TMV or X. euvesicatoria, respectively, than WT plants, further implicating WRKY7 in effector-triggered immunity.
Finally, Wu et al. observed that WRKY7 expression is strongly induced during effector-triggered immunity. Interestingly, induction of WRKY7 expression was compromised in NRG1 knockout plants, suggesting that NRG1 and WRKY7 are engaged in a positive feedback loop, with each amplifying the expression of the other. Positive feedback loops can rapidly and robustly amplify responses to stimuli and, in the absence of additional regulatory mechanisms, irreversibly determine cell fate. Effector-triggered immunity typically culminates in programmed cell death to restrict the spread of the pathogen; the WRKY7-NRG1 circuit may contribute to a swift induction of cell death and resistance.
In this work, Wu et al. shed light on the role of WRKY7 in regulating NRG1 expression and modulating basal and effector-triggered immunity (Fig.). The authors note that there appear to be multiple cis elements upstream of the NRG1 coding region and that NRG1 expression is likely modulated by numerous TFs to integrate different signals. Additionally, other WRKY TFs may be involved in regulation of NRG1 through the identified W-box motifs in the NRG1 promoter. Conversely, it remains to be explored whether WRKY7 regulates expression of other components of the signaling pathway leading to NRG1 activation or the plant immune system more broadly.
While untangling the intricate network of regulation of NLR expression requires significant work, it is a highly useful endeavor. Transfer of NLR proteins between plant species has the potential to confer novel pathogen resistance in agriculturally important crops. Furthermore, a combination of structural data and developments in artificial intelligence for protein modeling and design has accelerated efforts to engineer synthetic NLR proteins with new-to-nature recognition profiles (reviewed by Zdrzałek et al. 2023). However, overexpression of some NLR proteins leads to deleterious phenotypes, such as spontaneous cell death and restricted growth, while insufficient expression prevents robust activation of defense. An improved understanding of the transcriptional, translational, and posttranslational regulation of NLR proteins, such as NRG1, may aid efforts to introduce NLR genes into crops. This study by Wu et al. represents a timely contribution to our understanding of the transcriptional regulation of an important component of the plant immune system.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bentham AR, de la Concepcion JC, Mukhi N, Zdrzałek R, Draeger M, Gorenkin D, Hughes RK, Banfield MJ. A molecular roadmap to the plant immune system. J Biol Chem. 2020:295(44):14916–14935. 10.1074/jbc.REV 120.01085232816993 PMC 7606695 · doi ↗ · pubmed ↗
- 2Contreras MP, Lüdke D, Pai H, Toghani A, Kamoun S. NLR receptors in plant immunity: making sense of the alphabet soup. EMBO Rep. 2023:24(10):e 57495. 10.15252/embr.20235749537602936 PMC 10561179 · doi ↗ · pubmed ↗
- 3Feehan JM, Wang J, Sun X, Choi J, Ahn HK, Ngou BPM, Parker JE, Jones JDG. Oligomerization of a plant helper NLR requires cell-surface and intracellular immune receptor activation. Proc Natl Acad Sci U S A. 2023:120(11):e 2210406120. 10.1073/pnas.221040612036877846 PMC 10089156 · doi ↗ · pubmed ↗
- 4Ibrahim T, Yuen ELH, Wang H-Y, King FJ, Toghani A, Kourelis J, Vuolo C, Adamkova V, Castel B, Jones JDG, et al A helper NLR targets organellar membranes to trigger immunity [preprint; not peer reviewed]. bio Rxiv. 2024. 10.1101/2024.09.19.613839 · doi ↗
- 5Jacob P, Kim NH, Wu F, El-Kasmi F, Chi Y, Walton WG, Furzer OJ, Lietzan AD, Sunil S, Kempthorn K, et al Plant “helper” immune receptors are Ca 2+-permeable nonselective cation channels. Science. 2021:373(6553):420–425. 10.1126/science.abg 791734140391 PMC 8939002 · doi ↗ · pubmed ↗
- 6Javed T, Gao SJ. WRKY transcription factors in plant defense. Trends Genet. 2023:39(10):787–801. 10.1016/j.tig.2023.07.00137633768 · doi ↗ · pubmed ↗
- 7Jia A, Huang S, Song W, Wang J, Meng Y, Sun Y, Xu L, Laessle H, Jirschitzka J, Hou J, et al TIR-catalyzed ADP-ribosylation reactions produce signaling molecules for plant immunity. Science. 2022:377(6605):eabq 8180. 10.1126/science.abq 818035857644 · doi ↗ · pubmed ↗
- 8Lapin D, Kovacova V, Sun X, Dongus JA, Bhandari D, Von Born P, Bautor J, Guarneri N, Rzemieniewski J, Stuttmann J, et al A coevolved EDS 1-SAG 101-NRG 1 module mediates cell death signaling by TIR-domain immune receptors. Plant Cell. 2019:31(10):2430–2455. 10.1105/tpc.19.0011831311833 PMC 6790079 · doi ↗ · pubmed ↗