The Arctic copepod Calanus hyperboreus is more tolerant to marine heatwaves than temperate copepods in the Oslofjord
Mathieu Lutier, Andrea Emilie Thorstensen Skari, Nele Thomsen, Helena C. Reinardy, Khuong V. Dinh

TL;DR
Arctic copepods in the Oslofjord show higher heat tolerance than expected, suggesting local adaptation to warming waters.
Contribution
The study reveals local thermal adaptation in Calanus hyperboreus, challenging assumptions about its vulnerability to warming.
Findings
Calanus hyperboreus survived up to 16–20°C, higher than temperate species in the Oslofjord.
C. hyperboreus showed minimal DNA damage, indicating physiological adaptation to warmer conditions.
The findings suggest potential resilience of C. hyperboreus to marine heatwaves in southern Arctic regions.
Abstract
Calanus hyperboreus plays a key role in the functioning of Arctic ecosystems. It is considered highly vulnerable to ocean warming and marine heatwaves, which would reduce its range, expected to shift northward. Yet, C. hyperboreus is reported as far south as the Skagerrak, where it is considered non-native and transported by ocean currents. We argue that this may be an isolated population adapted to warmer temperatures. To test this hypothesis, we exposed C. hyperboreus from the Oslofjord to temperatures from 0°C to 24°C, for 5 days. We recorded survival to identify the upper threshold of thermal tolerance and DNA damage to detect sublethal effects. The thermal response of C. hyperboreus was compared with that of the dominant copepod species in the Oslofjord, Calanus finmarchicus and Metridia longa. We found that the survival of C. hyperboreus did not decrease before reaching 16°C–20°C…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
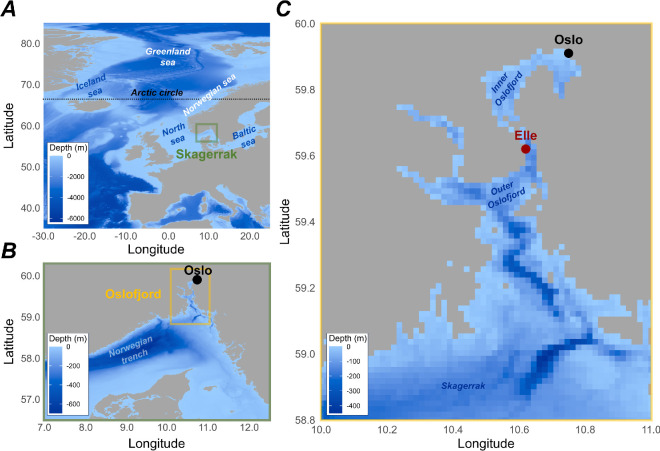 Figure 1
Figure 1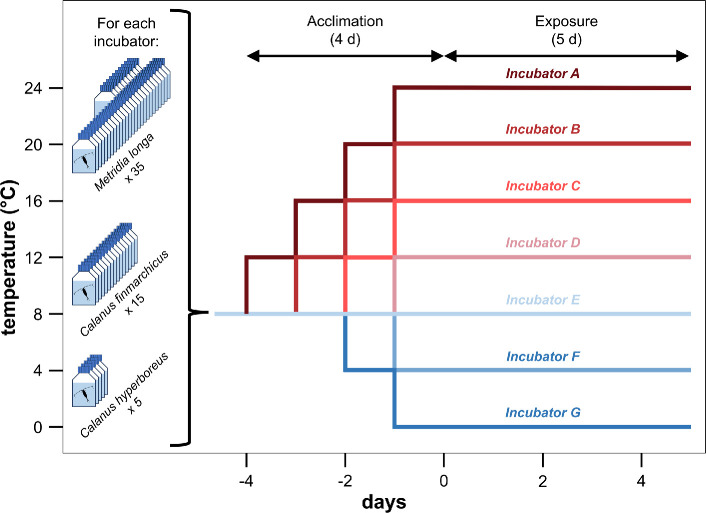 Figure 2
Figure 2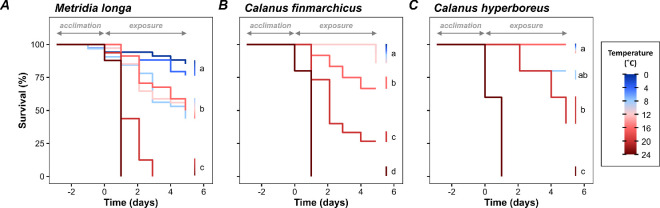 Figure 3
Figure 3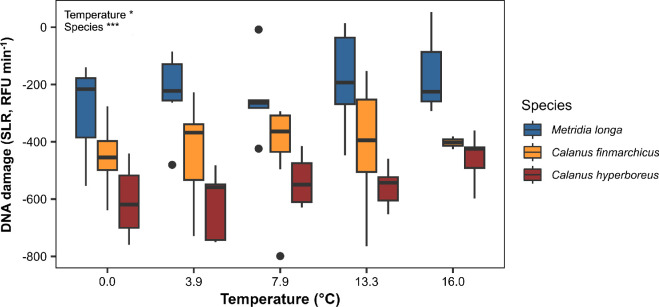 Figure 4
Figure 4- —Norges Forskningsrådhttp://dx.doi.org/10.13039/501100005416
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine and coastal ecosystems · Ocean Acidification Effects and Responses · Marine Ecology and Invasive Species
Introduction
Climate change will cause ocean temperatures to increase by 1°C−5°C by the end of the twenty-first century, a phenomenon called ocean warming (OW) [1,2]. The Arctic Ocean is the most vulnerable, warming 4−7 times faster than the global ocean average [3]. Marine heatwaves (MHW) are periods when surface waters are much warmer than the normal seasonal average, up to +5°C, which can persist for months [4]. Climate change makes MHW exponentially more frequent [5]. Temperature is one of the main parameters controlling the biology of ectothermic organisms [4,6]. Each organism has a temperature optimum and an upper tolerance threshold beyond which its homeostasis and survival are compromised [7,8]. Thus, OW and MHW are restricting the habitats and shifting the distribution of many species [9–11]. This causes a poleward shift of species distributions and reduces the range of polar species, threatening some of them with extinction [12,13].
A striking example is the Arctic copepod Calanus hyperboreus (Krøyer), which could experience a northward shift of its distribution by up to 70 km per decade [14]. Calanus hyperboreus presents unique features in copepods being one of the largest, up to 7 mm prosome length, and the richest in lipids, up to 60% of its dry body weight (DW) [15–17]. It is the preferential prey of higher trophic levels, particularly Arctic fishes and seabirds, in regions where it dominates zooplankton [15]. After storing lipid reserves over spring/summer, C. hyperboreus migrates to the deep sea where it overwinters at 600−3000 m depth [15,17]. By natural mortality and respiration, this sequesters a huge amount of carbon in the deep sea [18]. Therefore, changes in the distribution of C. hyperboreus would have large consequences for ecosystems.
Calanus hyperboreus shows highest abundances in waters with temperature ranges between 0°C and 5°C and is rare above 7°C [19–21]. The upper thermal tolerance threshold for its survival has been determined at 10°C–15°C [22,23]. Thus, the heart of the species’ distribution range is in Arctic regions and mainly in the Greenland Sea, where the highest abundances are recorded (figure 1; [15]). Yet, C. hyperboreus is observed as far south as the Skagerrak at the mouth of the Baltic Sea (figure 1; [24–29]). In these areas, the upper tolerance limit of 10°C–15°C is commonly exceeded [30] which is amplified by MHW [31]. Calanus hyperboreus is considered a non-native species to the North Sea, where it is thought to be transported from the north by ocean currents [24,27,32]. Overall, the species abundance is negligible in the Norwegian and North Seas (figure 1). However, data from recent cruises found high abundances, up to 3.2 g DW m^−2^, of C. hyperboreus in the deepest part of the Skagerrak (approx. 600 m; [28,29]). Such abundance is similar to those reported in many Arctic regions such as the Makarov, Canada, Nansen and Amundsen Basins [33–36]. This highlights a clear disconnection in C. hyperboreus distribution with negligible abundances between the Iceland Sea and Skagerrak. The isolation of the Skagerrak C. hyperboreus population is also supported by the smaller size of its genome in comparison with Arctic conspecifics [25]. Therefore, we hypothesize that C. hyperboreus from Skagerrak is an isolated population adapted to warmer water temperatures.
Geography of the Oslofjord, the study location. (A) The position of the Skagerrak in the North Sea, a subregion of the Atlantic Ocean, is shown (green square). (B) The position of the Oslofjord in the Skagerrak is shown (yellow square). (C) The geography of the Oslofjord is shown, as well as the location of Elle, the sampling station (red dot). The positions of major seas, the Arctic Circle (dotted line), and points of interest cited in the article are indicated with text, to be used as spatial references.
To test this hypothesis, we exposed C. hyperboreus collected in the Oslofjord (north Skagerrak) to temperatures from 0°C to 24°C. Survival was recorded for 5 days to determine the upper thermal tolerance threshold. To detect sublethal effects, we also measured the amount of DNA damage that can reflect oxidative stress caused by acute heat shock [37]. For reference, we compare the thermal response of C. hyperboreus with that of the dominant copepod species in the Oslofjord at the time of sampling, Calanus finmarchicus (Gunnerus) and Metridia longa (Lubbock). Calanus finmarchicus and M. longa are commonly observed in the Oslofjord, and we expect them to be more tolerant to warming. Determining the existence of a heat-tolerant population is crucial because local adaptations can provide a reservoir of resistant genotypes during rapid environmental changes [38,39].
Material and methods
Sampling
2.1.
The Oslofjord copepod community was sampled on 5 June 2023, near Elle Station (59.62° N, 10.62° E; figure 1). The Oslofjord is the northern branch of the Skagerrak, the strait forming the border between the North Sea and the Baltic Sea (figure 1). Sampling was carried out using a WP2 net (200 μm mesh) with three vertical hauls from the bottom (203 m) to the surface. The seawater temperature below the thermocline, where most calanoid copepods are distributed during the day [40,41], was recorded with a Conductivity, Temperature, and Depth instument (CTD) and was approximately 8°C. Samples were then transported to the laboratory at the University of Oslo, where copepod species were sorted at approximately 8°C, under a microscope, in a climate room. Species and life stages were identified based on morphological criteria and the number of segments on their urosome. Copepodite 5 (copepodite V) and adult females (AF) were kept for the experiment at a ratio of 69 and 31%, respectively. The other two dominant species were M. longa (51% AF, 46% adult males (AM) and 3% copepodite V) and C. finmarchicus (87% copepodite V, 9% AF, 3% AM and 1% copepodite IV). For each species, we used a mix of life stages (as presented above) that represented the population demography at the time of sampling. After sorting, copepods were maintained in opened bottles of filtered seawater at a density of 45 copepods l^−1^ in an incubator at 8°C. The same batch of seawater was used to maintain copepods throughout the experiment. This seawater was collected on 5 June 2023 at Drøbak Aquarium (pumped at 40 m depth in the Oslofjord). The seawater was filtered at 1 μm, treated with a UV lamp and stored at 4°C. Air was bubbled into the seawater to ensure good oxygenation. Copepods were not fed during the experiments. All three species have large lipid reserves to survive the low-feeding season [18,42,43], and these are unlikely to have decreased during the short duration of the experiment. This was verified by lipid droplet measurements (see below).
Experimental design
2.2.
Calanus finmarchicus and M. longa were exposed to seven different temperatures: 0, 4, 8, 12, 16, 20 and 24°C. These conditions cover the range of temperatures recorded on the whole water column at the sampling station during monthly measurements in 2020 that ranged from 3°C to 20°C (E.S. Garvang, 2024, personal communication). The 24°C condition was selected to test the influence of future OW and MHW. Each temperature condition was maintained in one of seven incubators in which copepod exposures were performed.
On 14 June 2023, individual copepods were placed in plastic flasks (VWR^®^ tissue culture flasks) filled with 65 ml of seawater. A total of 35 M. longa, 15 C. finmarchicus and 5 C. hyperboreus were then placed in each of the seven incubators set at an initial temperature of 8°C (figure 2). The different sample sizes for the three species are because of the different abundances found in the field sample. For 4 days, the temperature was then increased (for conditions above 8°C) or decreased sequentially (for conditions below 8°C) by 4°C d^−1^ so that all final temperature conditions were reached on the same day in the different incubators (figure 2). The exposure lasted 5 days because it is the minimum duration of a MHW in the environment [44]. Survival was checked daily by gently tapping on the wall of the flask with a pin to stimulate the copepods, and immobile copepods were considered dead. Temperature was recorded in each incubator every 15 min using HOBO^©^ TidbiT v2 loggers (one logger per incubator) placed in a 65 ml beaker, sealed with parafilm and filled with seawater. The average temperature in each incubator during the exposure period was calculated and used for statistics. Photographs of copepods were taken daily, before removal, for dead copepods, and at the end of the experiment, before sampling, for survivors, using a stereomicroscope (Leica M 125 C, LAS X software). Prosome size, lipid droplet and prosome surface areas were measured using ImageJ. Lipid reserve (%) was determined as lipid/ prosome area.
Experimental design used to determine the temperature tolerance of three species of copepods from the Oslofjord: M. longa, C. finmarchicus and C. hyperboreus. Variations in temperature over time are presented for the seven incubators, each containing 35 M. longa, 15 C. finmarchicus and 5 C. hyperboreus. The first 4 days corresponded to the gradual changes of temperature in the incubators. The exposure then lasted 5 days for all the temperature conditions.
DNA damage analyses
2.3.
The Fast MicroMethod assay (FMM) was used for determining the amount of DNA damage by assessing the unwinding kinetics of double-stranded DNA in alkaline solution with fluorescence measurements [45,46]. For each species, at the end of the experiment, 5 ± 1 copepods (depending on the final survival; electronic supplementary material, table S1) from each temperature condition were individually snap**-**frozen in liquid nitrogen and stored at −80°C. Our protocol is adapted from Halsband et al. [47]. Individual copepods were gently homogenized with a hand pestle in 260 μl of 20 mM Ethylenediaminetetraacetic acid (EDTA) + 10% Dimethyl sulfoxide (DMSO). About 20 μl of homogenate, or 20 mM EDTA + 10% DMSO for the blank, was loaded, with five analytic replicates per sample, into a microplate (96-well black-walled, Greiner Bio-One Ltd). Samples were lysed on ice for 40 min in the dark after adding 20 μl of lysis buffer to each well (20 µl 9M urea, 0.1% Sodium dodecyl sulfate (SDS), 0.2M EDTA, 2% Quant-iT™ PicoGreen^®^). After lysis, unwinding was initiated by adding 200 μl of unwinding solution (mixture of 20 mM EDTA and 2 M NaOH until pH is adjusted to 13.0) to each well. Fluorescence was then recorded immediately using a SynergyMx^®^ microplate reader (BioTek^©^) and every minute for 30 min (excitation 485 nm, emission 520 nm).
We estimated the amount of DNA damage for each sample as the slope of the linear regression (SLR) of fluorescence (expressed in relative fluorescence unit, RFU) with time, expressed in RFU min^−1^ [48,49]. This is different from the strand scission factor, which is commonly used to estimate the amount of DNA damage but was less suitable for our experimental design (see more in the electronic supplementary material, note S1). An increase in SLR shows an increase in DNA damage. The linear regression of fluorescence over unwinding time was fitted between 0 and 20 min, representing the linear part of the DNA unwinding. We only retained significant linear regressions (p < 0.05), which respected the assumptions of normality and homoscedasticity of the residuals. The three best analytic replicates were kept for each sample using z-scores, i.e. the three replicates that diverged the less from the mean SLR of the five replicate wells [50]. The mean SLR was then calculated from these three replicates for each sample and kept for statistics.
Statistics
2.4.
Analyses were performed using the R software v. 4.3.1, and the statistical significance threshold was 0.05. The effects on survival of temperature, species, prosome length, lipid reserves and the interaction of all these factors were tested using a Cox proportional-hazards model using the coxme function of the coxme package [51]. We did not include DNA damage as a factor influencing survival because it is not measurable in dead copepods. The different factors were tested and the model with the lowest Akaike information criterion and Bayesian information criterion values was retained. Therefore, only temperature and species were kept while size and lipid reserves did not affect survival. The assumption of proportional hazards of the Cox model was graphically checked by plotting the Schoenfeld residuals against time using the ggcoxzph function of the survminer package [52]. For significant factors, i.e. temperature and/or species, post hoc pairwise comparisons were performed using the pairwise_survdiff function. Sample sizes are unbalanced for the three species (n = 245 for M. longa, n = 105 for C. finmarchicus and n = 35 for C. hyperboreus). Cox models are valid for an unbalanced design, but different samples sizes affect the statistical power, i.e. the ability to detect an effect of the factors [53–55]. To test differences in survival response of species at different exposure temperatures, a first general model was built for the three species, with an unbalanced design. Then individual models were tested for each species separately, with a balanced design. Kaplan–Meier survival curves were used to visualize survival as a function of time. Type II analysis of variance (ANOVA), for unbalanced designs (n = 28 for M. longa, n = 29 for C. finmarchicus and n = 24 for C. hyperboreus; electronic supplementary material, table S1), was used to test the effects of temperature, species and their interactions on DNA damage (SLR). Size and lipid reserves did not affect DNA damage. The homogeneity of variances and normality of residuals were checked graphically. The differences between groups for significant factors were tested using Tukey honestly significant difference (HSD) tests.
Results
Temperatures in the incubators were stable during the 5 days of exposure and were 0.0 ± 0.4°C, 3.9 ± 0.5°C, 7.9 ± 0.2°C, 13.3 ± 0.2°C, 16.0 ± 0.2°C, 19.9 ± 0.3°C, and 23.8 ± 0.2°C (mean ± s.d., n = 670). The survival curves were significantly different for species (p < 0.001) and temperatures (p < 0.001) (Cox proportional-hazards model, electronic supplementary material, table S2). Survival of M. longa was generally lower than those of C. finmarchicus and C. hyperboreus, which were not significantly different (post hoc tests, figure 3). The interaction of the two factors was significant (p < 0.001), indicating that the response to temperature was different for the three species. Individual models for each species confirm that temperature significantly decreased the survival of M. longa (p < 0.001, figure 3A), C. finmarchicus (p < 0.001, figure 3B), and C. hyperboreus (p < 0.001, figure 3C).
Survival of three copepod species from the Oslofjord as a function of time and temperature. Kaplan–Meier survival curves are shown for (A) M. longa, (B) C. finmarchicus and (C) C. hyperboreus. The temperature during the exposure period is represented by the colour gradient. Survival curves that differ significantly for different temperatures are indicated by different letters. Some survival curves overlap, this is the case for (B) temperatures 0.0, 3.9, 7.9 and 13.3°C and (C) 0.0, 3.9, 7.9, 13.3 and 16.0°C. Negative values for time on the x-axis indicate the days before the final temperature condition is reached, i.e. the ‘acclimatization’ period. The exposure period is indicated by positive values.
Metridia longa showed no difference in survival between 0.0°C and 3.9°C with a final survival rate of 81 ± 4% (post hoc test, figure 3A). Survival then significantly decreased at 7.9°C–16.0°C with a final survival rate of 48 ± 3%. Finally, there was no survival at 19.9°C and 23.8°C after 1 and 2.9 days of exposure respectively. Calanus finmarchicus showed no difference in survival between 0.0°C and 13.3°C with a final survival rate of 95 ± 6% (post hoc test, figure 3B). Survival significantly decreased at 16.0°C and 19.9°C with final survival rates of 67 and 27%, respectively. No survival was recorded at 23.8°C after 1 day of exposure. Calanus hyperboreus overall showed no difference in survival between 0.0°C and 16.0°C with a final survival rate of 96 ± 8% (post hoc test, figure 3C). Survival then significantly decreased at 19.9°C with a final survival rate of 40%. No survival was recorded at 23.8°C after 1 day of exposure.
Overall, regardless of temperature, DNA damage differed significantly for the three species (ANOVA type II, p < 0.001). DNA damage was higher in M. longa (−230 ± 164) than in C. finmarchicus (−426 ± 157) and lowest in C. hyperboreus (−556 ± 113) (Tukey’s HSD test, figure 4). The results of type II ANOVA indicate that temperature (p = 0.038) significantly increased DNA damage for all three species (electronic supplementary material, table S3). There is no interaction between temperature and species, indicating that the response of DNA damage to temperature was similar between species.
DNA damage in copepods from the Oslofjord that survived 5 days of exposure to different temperatures. Data are presented as box and whisker plots (averages and s.d.). The species are M. longa (blue), C. finmarchicus (orange) and C. hyperboreus (red). The results of type II ANOVA testing the effects of temperature, species and the interaction of the two factors on DNA damage are shown in the upper left corner.
Discussion
We experimentally study for the first time, to our knowledge, the ecophysiology of C. hyperboreus sampled in the Skagerrak, approximately 1500 km south of its Arctic range, where it is considered a non-native species, transported by ocean currents, with low resistance to warm temperatures. Unexpectedly, we find that C. hyperboreus is more resistant to warming than M. longa and C. finmarchicus, which are commonly found in the Oslofjord. This suggests the existence of a local C. hyperboreus population in Skagerrak, adapted to withstand warm temperatures. This should be considered when studying OW and MHW impacts on the species.
In the Oslofjord, the survival of M. longa is most vulnerable to OW and MHW, much more than that of C. finmarchicus. We show that survival of M. longa decreases above 4°C–8°C with no survival above 16°C–20°C, while survival of C. finmarchicus decreases above 13°C–16°C with no survival above 20°C–24°C. Metridia longa is abundant in cold Arctic and boreal waters [56,57] and do not survive above 14°C–18°C [22], with the Oslofjord being the southern limit of its distribution [58]. Calanus finmarchicus is a boreal species abundant between 0°C and 10°C [20,23] and do not survive above 20°C [22]. If C. hyperboreus is a non-native species from the Skagerrak transported there by ocean currents, we expect it to be more sensitive to warming.
Contrary to expectations, we found that, in the Oslofjord, C. hyperboreus is more resistant to warming than M. longa and C. finmarchicus. Indeed, C. hyperboreus survival decreases above 16°C–20°C with no survival above 20°C–24°C. This is far greater than what is reported for Subarctic/Arctic C. hyperboreus that do not survive above 10°C–15°C [22,23]. However, comparisons with literature should be made with caution since we use a different protocol. The strength of our study is comparing the tolerance of C. hyperboreus with the other species occurring in Oslofjord that should be adapted to warmer conditions. Because of different abundances in the field, we had 7 and 3 times fewer C. hyperboreus per condition than M. longa and C. finmarchicus, respectively. Therefore, the probability of having a high proportion of individuals surviving after 5 days was much lower for C. hyperboreus, and yet this occurred between 0°C and 16°C, suggesting our results are conservative. There was a difference in the proportions of life stages between the three copepod species used in the experiment. In some copepod species, older individuals survive stress better than younger individuals [59–61]. In our study, there were more adults in M. longa (51%) than in C. hyperboreus (31%) and significantly more than in C. finmarchicus (9%). Therefore, C. hyperboreus may have faced a disadvantage in terms of survival. Yet, it survived MHW much better than M. longa, confirming the robustness of our observations.
We find that C. hyperboreus exhibits less DNA damage than C. finmarchicus, with both species exhibiting less DNA damage than M. longa. DNA damage occurs continuously in organisms owing to exogenous (e.g. UV, environmental stressors) or endogenous (production of reactive oxygen species (ROS) by metabolism) factors [62]. At the same time, organisms continually repair DNA to ensure their survival [62]. FMM determines the net amount of DNA damage, detected as single- and double-stranded DNA breaks and alkali labile sites, which result from both concomitant damage and repair [46,48]. This is a good indicator of environmental stress in marine invertebrates [46,48]. If C. hyperboreus cannot survive in the Skagerrak and is a non-native species transported by ocean currents, as suggested [24,27,32], one would expect the species to have high levels of DNA damage owing to stress. Conversely, C. hyperboreus exhibits overall less DNA damage than C. finmarchicus and M. longa, suggesting that its biology is adapted to life in the Oslofjord. Temperature can increase the rate of DNA damage either by increasing ROS production through metabolism or by decreasing DNA repair by altering enzyme activity [37,63]. Here, we indeed find that DNA damage increases with temperature in all three species without interspecific differences.
Our results suggest the presence of an isolated population of C. hyperboreus in the Skagerrak that is adapted to warmer temperatures than Arctic conspecifics. The literature on C. hyperboreus outside the Arctic is scarce but supports our hypothesis. First, C. hyperboreus from the Oslofjord has smaller genomes than in the Arctic, which is generally associated with adaptation to warmer temperatures [25]. Second, there is a discontinuity in the range of the species. Indeed, C. hyperboreus are negligible in the Norwegian and North Seas. However, abundances in the Skagerrak, off Kristiansand, can be as high as 3.2 g DW m^−2^ [28,29]. These abundances are similar to those in much of the Arctic Ocean, such as the Amundsen, Canada, Makarov and Nansen basins, as well as Svalbard waters, which are typically 2−3 g DW m^−2^ [33–36]. It is unlikely that C. hyperboreus would concentrate in Skagerrak only through advection via oceanic currents as it is often suggested [24,27,32]. All surface currents outflow from Skagerrak, the only current coming from the North is the Norwegian trench inflow in the deep (<200 m) [64]. The Norwegian trench inflow transports overwintering C. finmarchicus from the Norwegian sea into Skagerrak during the winter [65,66]. However, C. hyperboreus are considered unable to overwinter at such southern latitudes [24,27,32].
Calanus hyperboreus is a key species in Boreal and Arctic ecosystems as the main prey of many fish species and seabirds, and by storing carbon in the deep sea via overwintering migration [15,17,18]. Models predict that the species' range will shift northwards with OW, losing up to 70 km per decade in the coming years [14], with large consequences for ecosystems. Here we show that local adaptations to warmer waters probably exist in the Skagerrak and should be considered when projecting the future of C. hyperboreus. Further studies directly comparing the ecology, genetics, connectivity and stress tolerance threshold of C. hyperboreus between Skagerrak and Arctic populations would greatly improve our knowledge of local adaptations and global change. This is crucial since the existence of local adaptations can constitute a reservoir of resistant genotypes, which can benefit the persistence of species in the face of environmental changes [38,39].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sluijs A et al. 2006 Subtropical Arctic Ocean temperatures during the Palaeocene/Eocene thermal maximum. Nature 441, 610–613. (10.1038/nature 04668)16752441 · doi ↗ · pubmed ↗
- 2IPCC. 2021 Climate change 2021 – the physical science basis: working group I contribution to the sixth assessment report of the intergovernmental panel on climate change. Cambridge, UK: Cambridge University Press. (10.1017/9781009157896) · doi ↗
- 3Rantanen M, Karpechko Ay, Lipponen A, Nordling K, Hyvärinen O, Ruosteenoja K, Vihma T, Laaksonen A. 2022 The Arctic has warmed nearly four times faster than the globe since 1979. Commun. Earth Environ. 3, 10. (10.1038/s 43247-022-00498-3) · doi ↗
- 4Orton JH. 1920 Sea-temperature, breeding and distribution in marine animals. J. Mar. Biol. Assoc. UK 12, 339–366. (10.1017/S 0025315400000102) · doi ↗
- 5Frölicher TL, Fischer EM, Gruber N. 2018 Marine heatwaves under global warming. Nature 560, 360–364. (10.1038/s 41586-018-0383-9)30111788 · doi ↗ · pubmed ↗
- 6Cossins A. 2012 Temperature biology of animals. Berlin; Heidelberg, Germany: Springer Science & Business Media.
- 7Huey RB, Kingsolver JG. 1989 Evolution of thermal sensitivity of ectotherm performance. Trends Ecol. Evol. 4, 131–135. (10.1016/0169-5347(89)90211-5)21227334 · doi ↗ · pubmed ↗
- 8Pörtner HO. 2002 Climate variations and the physiological basis of temperature dependent biogeography: systemic to molecular hierarchy of thermal tolerance in animals. Comp. Biochem. Physiol. Mol. Integr. Physiol. 132, 739–761. (10.1016/S 1095-6433(02)00045-4)12095860 · doi ↗ · pubmed ↗