Simultaneous Detection of Four Madurella Species Using Loop-Mediated Isothermal Amplification (LAMP) for Eumycetoma Diagnosis
Isato Yoshioka, Ahmed Hassan Fahal, Doudou Sow, Satoshi Kaneko, Yugo Mori, Sayaka Ban, Takashi Yaguchi

TL;DR
This paper introduces a new LAMP method to detect four Madurella species causing eumycetoma, a tropical disease, enabling faster and simpler diagnosis.
Contribution
A novel LAMP primer set was developed for simultaneous detection of four Madurella species with high specificity and sensitivity.
Findings
The LAMP primer sets detected up to 1 pg of genomic DNA from all four Madurella species.
The method showed no cross-reactivity with other pathogenic fungi.
A selected primer set demonstrated improved reactivity for potential clinical use.
Abstract
Eumycetoma is a neglected tropical disease caused primarily by a Madurella mycetomatis infection, besides other related species. In this study, we designed a novel loop-mediated isothermal amplification (LAMP) primer set capable of simultaneously detecting four Madurella species (M. mycetomatis, M. pseudomycetomatis, M. tropicana, and M. fahalii). Genomic sequencing of M. pseudomycetomatis strain and comparative genome analysis revealed the candidate genes that were common among and specific to Madurella species. The 3 LAMP primer sets targeting these genes detected up to 1 pg of the genomic DNA of all 4 Madurella species, exhibiting no cross-reactivity toward other pathogenic fungi. Among these, one primer set showing better reactivity was selected as a candidate used for diagnosis. Therefore, we developed novel primer sets which enabled the simultaneous detection of four Madurella…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
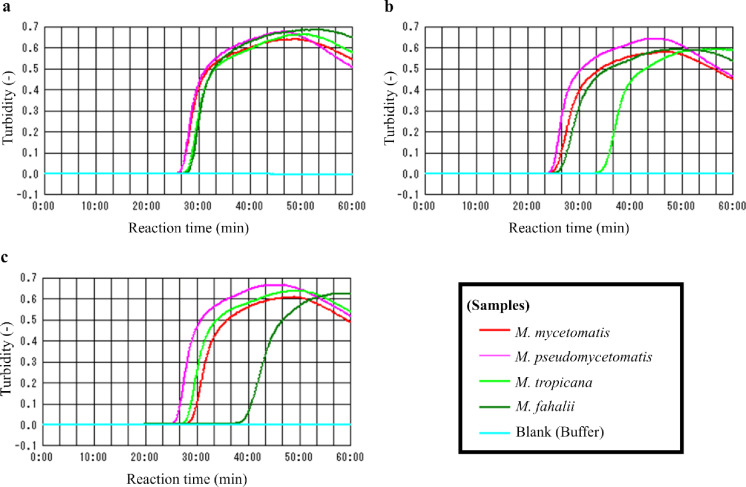 Figure 1
Figure 1- —Japan Agency for Medical Research and Development (AMED)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsActinomycetales infections and treatment · Infectious Diseases and Mycology · Fungal Infections and Studies
Short Communication
Mycetoma is a neglected tropical disease caused by microbial infection, resulting in chronic granulomatous inflammation of the subcutaneous tissue discharging grains [1, 2]. Although a wide range of microorganisms, including actinomycetes and fungi, have been reported as causative agents for this disease, the most frequently cited microbe in fungal mycetoma (eumycetoma) is Madurella mycetomatis [3, 4]. In addition, all of the known species belonging to the genus Madurella (M. mycetomatis, M. pseudomycetomatis, M. tropicana and M. fahalii) are known to cause eumycetoma [4]. Due to the presence of small and painless subcutaneous swellings as the initial symptoms of the slowly progressing mycetoma, misdiagnosis sometimes occurs and leads to high morbidity and difficulty in achieving a cure [5]. Therefore, a sensitive detection tool specific to Madurella is urgently needed for the early diagnosis of mycetoma, particularly in remote rural settings.
Considering this diagnostic scenario, various molecular methodologies have been developed over the years to detect the causative agent of mycetoma. For example, a polymerase chain reaction (PCR) assay specific to M. mycetomatis or Madurella species has been published [6, 7]. However, isothermal DNA amplification appears more feasible compared with PCR because an isothermal reaction does not require expensive instrumentation that would otherwise be needed for thermal cycling and/or fluorescence monitoring, both of which may not be available at a low-resource rural hospital [8]. Based on this rationale, we have previously constructed a loop-mediated isothermal amplification (LAMP) system targeting ribosomal DNA in Madurella species [9]. Although we have previously described the simple identification of M. mycetomatis and M. fahalii by a combination of three primer sets (three reactions) [9], the development of a simpler and more versatile system will contribute to the initial screening for Madurella by reducing diagnostic costs and enhancing throughput.
In this study, we describe the design of a novel primer set that can simultaneously detect all Madurella species in a single reaction. To achieve this goal, we conducted a comparative genome analysis that enabled us to design and evaluate primer sets targeting candidate genes common to Madurella species.
A total of 37 fungal strains (including 22 species) and five bacterial strains (including five species) known to cause mycetoma [3, 4] were used in this study, as listed in Table 1. These strains have been preserved at the Medical Mycology Research Center (MMRC; Chiba University, Japan) as an IFM collection through the National Bio-Resource Project (Japan) or at the Mycetoma Research Center (MRC) (Khartoum University, Khartoum, Sudan). Otherwise, other strains were obtained from the culture collections of the Westerdijk Fungal Biodiversity Institute (CBS; Netherlands), the Biological Resource Center, National Institute of Technology and Evaluation (NBRC; Japan), the American Type Culture Collection (ATCC; United States), the Fungal Genetics Stock Center (FGSC; United States), and the Japan Collection of Microorganisms, RIKEN BioResource Research Center (JCM; Japan).Table 1. Fungal and bacterial strains used in this studySpeciesStrain NoFungiMadurella mycetomatisIFM 46458IFM 68169 (= MRC No. 6)IFM 68172 (= MRC No. 14)IFM 68173 (= MRC No. 16)IFM 68174 (= MRC No. 21)IFM 68175 (= MRC No. 22)IFM 68176 (= MRC No. 26)IFM 68177 (= MRC No. 31)IFM 68178 (= MRC No. 33)IFM 68239 (= MRC No. 10)IFM 68240 (= MRC No. 16)IFM 68243 (= MRC No. 28)IFM 68244 (= MRC No. 32)Madurella pseudomycetomatisIFM 46460Madurella tropicanaCBS 201.38^T^Madurella fahaliiCBS 129176^T^IFM 68170 (= MRC No. 9)IFM 68171 (= MRC No. 13)IFM 68242 (= MRC No. 25)Aspergillus fumigatusAf293 (= FGSC A1100)Aspergillus nigerIFM 68248Aspergillus nidulansIFM 68245 (= MRC No. 4)Aspergillus terreusIFM 67509Chaetomium globosumCBS 148.51^T^Chaetomium rectangulareCBS 126778^T^Curvularia lunataIFM 64654Exophiala jeanselmeiIFM 67393Falciformispora senegalensisCBS 196.79^ T^Falciformispora tompkinsiiCBS 200.79Fusarium oxysporumIFM 61457Fusarium solaniIFM 67677Neocosmospora falciformisCBS 475.67Neotestudina rosatiiCBS 331.78Sarocladium kilienseCBS 158.61Scedosporium apiospermumIFM 66388Thermothielavioides terrestrisCBS 355.66 (= NBRC 9121)Trichophyton rubrumIFM 66913BacteriaActinomadura maduraeIFM 0585^ T^ (= JCM 7436^ T^)Actinomadura pelletieriIFM 0590^ T^ (= JCM 3388^ T^)Nocardia asteroidesIFM 10791Nocardia brasiliensisIFM 12361Streptomyces somaliensisIFM 11185
The bacterial and fungal genomic DNA samples were extracted using the benzyl chloride method, as described previously [9]. In addition, the genomic DNA samples of M. mycetomatis IFM 46458 and M. pseudomycetomatis IFM 46460 were further purified using ISOSPIN Plant DNA (Nippon Gene, Toyama, Japan) in accordance with the instructions of the manufacturer for the evaluation of LAMP primer sets. Additionally, the extracted genomic DNA samples of M. tropicana CBS 201.38^ T^ and M. fahalii CBS 129176^ T^ were obtained from the CBS. The DNA concentration of these DNA samples was determined by QuantiFluor ONE dsDNA System (Promega, Madison, WI, USA).
For genomic sequencing, M. pseudomycetomatis IFM 46460 was cultivated in Sabouraud medium, and its genomic DNA was extracted by the phenol–chloroform method and purified with a Genomic-tip 100/G column (Qiagen, Hilden, Germany) as described previously [10]. The extracted genomic DNA was sequenced by Genome-Lead Co., Ltd. (Kagawa, Japan) via the Illumina NovaSeq 6000 and Nanopore PromethION platforms (Oxford Nanopore Technologies [ONT], Cambridge, UK). The genome assembly and gene annotation were performed in accordance with the methods described previously, with some modifications [10]. Briefly, genome assembly was executed using the ONT long reads not mapped to the mitochondrial genome by NECAT software v.0.0.1 [11] and Flye software v.2.9.2 [12] with a genome size of 35 Mb; however, the genome coverage value of input reads used for Flye assembly was set to a coverage × 50 in comparison with the previous study. Finally, genome annotation was performed by using the Funannotate software v.1.8.15 pipeline (https://github.com/nextgenusfs/funannotate).
The genomic sequences of Madurella spp. and their closely related species, members of Chaetomiaceae and Podosporaceae [13, 14], were obtained from the DDBJ/EMBL/GenBank database through NCBI (https://www.ncbi.nlm.nih.gov/) and the JGI genome portal (https://genome.jgi.doe.gov/portal/). The details of the genomic sequences obtained from DDBJ/EMBL/GenBank are as follows: M. mycetomatis mm55 (accession: GCA_001275765.2) [15], Madurella fahalii IFM 68171 (accession: GCA_045866475.1) [10], Chaetomium globosum CBS 148.51 (accession: GCA_000143365.1) [16], Podospora anserina S mat + (accession: GCA_000226545.1) [17] and Mycothermus thermophilus UFV (accession: GCA_011316235.1) [18]. Additionally, the genomic sequences obtained from JGI were as follows: Thermothielavioides terrestris NRRL 8126 (ID: 1184802) [19] and Canariomyces arenarius (ID: 1018941) [20]. The orthologous genes were identified using annotated protein sequences through SwiftOrtho [21], with a minimum alignment coverage of 80% and a sequence identity cutoff of 50%. Subsequently, genes conserved exclusively among Madurella species were extracted from the SwiftOrtho results and further refined through an all-vs-all nucleotide-level comparison using VSEARCH [22], with a minimum sequence length of 500 bp and a nucleotide identity threshold of 90%. We used PrimerExplorer (https://primerexplorer.eiken.co.jp/e/) to design LAMP primer sets based on the genomic sequence of M. mycetomatis.
The LAMP reaction was performed in a 25 μL reaction volume containing template DNA, 40 pmol FIP and BIP, 20 pmol loop primers (LF and LB), and 5 pmol F3 and B3 in Loopamp DNA Amplification Reagent D (Eiken Chemical Co., Ltd; Japan). However, the reaction mixture of Primer #3 contained 20 pmol of three loop primers in comparison with the other primer sets (#1 and #2). Unless otherwise noted, the reaction was conducted at 66 °C for 60 min, and amplification was monitored in real-time by the change in turbidity in the LAMP reactions using LoopampEXIA (Eiken Chemical Co., Ltd; Japan). To determine the detection limit, genomic DNA solutions of M. mycetomatis IFM 46458 ranging from 100 pg to 100 fg were tested. In addition, 1 pg of the genomic DNAs of M. pseudomycetomatis IFM 46460, M. tropicana CBS 201.38 and M. fahalii CBS 129176 were used to assess both the sensitivity and specificity for Madurella species. Finally, 1 ng of genomic DNA was used per reaction tube to determine the optimal reaction temperature and to check specificity.
The chromosomal genome assembly of M. pseudomycetomatis IFM 46460 resulted in six scaffolds with a total length of 40,045,822 bp and an N50 value of 25,195,283 bp. Gene annotation identified 11,334 protein-coding genes, 180 tRNAs, and 79 rRNAs with a 99.6% BUSCO v.5.5.0 [23] score for eukaryote databases. A comparative genome analysis was conducted by using species evolutionarily close to the Madurella genus to identify genes common with and specific to the Madurella genus. A two-step screening protocol identified forty genes that were well-conserved across the genomes of Madurella genus members, as shown in Supplementary Table 1. Among these candidate genes, the LAMP primers we designed were specific for three genes (locus tag in M. mycetomatis mm55 [15]: MMYC01_204841, MMYC01_205181, MMYC01_210211), targeting gene regions with few or no mutations, as shown in Table 2.Table 2LAMP Primers used in this studyRegionSequence (5′ to 3′)Primer #1 (MMYC01_204841)F3AGCGGACTTGGCCTCATCB3GTAATCGTGGCTGCATTTCCAFIP (F1c/F2)CTCTCGTTGGCTCGGCAGAG-CGCCCCTCAAAGCTGGATBIP (B1c/B2)ACCGCCGATGCCTCGAATCCTA-TGTAGTTGTGGCGAATCTTTCCLFAGCTGCTCGGGCGGALBTCTCGGGCACCGGGTGPrimer #2 (MMYC01_205181)F3AGGGGCTCCCAGGAAATCB3TCGTCGCTGGAATCCTTCAFIP (F1c/F2)CAGTAGGCACAGTCCTCGATCCT-TCCATCAGCATCGCTAAACGBIP (B1c/B2)TTGGCATCAACATGCCCATGC-GATCTCCTCCTGGAGTCGGLFTCTTTGCCGCCCACGACATGLBTGTATGGCGAGAACGAAAAGGCPrimer #3 (MMYC01_210211)F3ATCGTCCGTCGTCCGCB3CGTAATCCTCAGCTCCCAFIP (F1c/F2)AGTTGCGCTGCAGCCTGAC-GACCGAGCAAGGTGACAAAGBIP (B1c/B2)CACTCGCTTGTCGAGGAACTCA-TCCTAACGTCGTCGAAAACALFGGGCGACCGCGTGTCLB1ACGACGCTGCTGCCGLB2AGAACGACGCCACCGC
The primer sets were tested by employing 100 pg of genomic DNA samples derived from M. mycetomatis IFM 46458 and M. fahalii CBS 129176 and running the LAMP reactions at temperatures ranging from 64 to 67 °C. The optimal temperature of the three primer sets was determined to be 66 °C, although they exhibited efficient amplification curves in all conditions, as shown in Supplementary Fig. 1. We determined the sensitivity of the three primer sets by employing 100 fg—100 pg of genomic DNA samples derived from M. mycetomatis IFM 46458. All three primer sets could detect up to 1 pg of genomic DNA, suggesting that they possess equal sensitivity, as shown in Supplementary Fig. 2. Subsequently, we evaluated the detection limits of other Madurella species. Although Primer #3 showed a reduced reactivity toward M. fahalii, these primer sets could also detect 1 pg of genomic DNA samples derived from three Madurella species (M. pseudomycetomatis, M. tropicana, M. fahalii) in approximately 45 min, as shown in Fig. 1.Fig. 1. The specificity of designed LAMP primers, a #1 (for MMYC01_204841), b #2 (for MMYC01_205181) and c #3 (for MMYC01_210211). 1 pg of the extracted genomic DNAs derived from Madurella spp. were used as templates. The red, magenta, yellow-green, and green plots represent the data for M. mycetomatis IFM 46458, M. pseudomycetomatis IFM 46460, M. tropicana CBS 201.38^ T^, and M. fahalii CBS 129176^ T^, respectively. The light blue plot indicates a blank reaction mixture, in which Tris–HCl buffer was used as a template instead of DNA
Since the three LAMP primer sets showed equal sensitivity towards Madurella, we conducted further tests to assess their specificity towards other pathogens and select the optimal primer set. We conducted LAMP reactions using 1 ng of genomic DNA from fungal and bacterial species known as the mycetoma causative agents, as listed in Table 1 [3, 9]. Primers #1, #2 and #3 showed no non-specific (unexpected) reaction toward pathogenic microorganisms other than the Madurella species*.* However, Primer #2 failed to amplify the genomic DNA of the M. fahalii strain (IFM 68242), while the other two primer sets successfully detected all tested strains belonging to the Madurella species.
We successfully developed three primer sets which could detect genomic DNA derived from four Madurella strains belonging to four species. However, Primer #2 failed to amplify one M. fahalii strain. As for the reactivity of the LAMP reactions at lower genomic DNA concentrations, the amplification rates of Primer #1 for four Madurella species were comparable, while Primer #3 exhibited lower reactivity for low-concentration genomic DNA samples of M. fahalii. These results suggest that Primer #1 is the first choice for its practical application to clinical diagnosis. Additionally, the detection limit of this primer set is comparable to that of LAMP primers designed in a previous study [9]. However, since our study only tested the extracted DNA from culture isolate, the LAMP reaction using a grain should be performed to evaluate the practical use in diagnosis in the future. Moreover, to facilitate diagnosis in rural settings, it is also necessary to adapt or extend simple detection methods that do not require expensive instruments, such as colorimetric and fluorescent assays.
In conclusion, as described above, a new primer set can detect all known Madurella species in a single reaction tube, whereas previous studies required two or three separate reactions. This advantage should enable medical workers to avoid complex operation protocols and to reduce the cost per experiment. Thus, this primer set can replace the previous version as a molecular diagnostic kit.
Supplementary Information
Below is the link to the electronic supplementary material.Supplementary file1 (DOCX 1999 KB)