Iron, copper and disulfide dysregulation: molecular crossroads of metabolic cell death in melanoma progression
Xinge Li, Yan Gao, Zhiyao Xing, Zichuan Liu, Xiangyang Zhang

TL;DR
This review explores how iron, copper, and disulfide imbalances drive metabolic cell death in melanoma and highlights new treatment strategies to overcome resistance.
Contribution
The paper integrates mechanisms of ferroptosis, cuproptosis, and disulfidptosis and proposes novel therapeutic strategies for melanoma.
Findings
Ferroptosis, cuproptosis, and disulfidptosis each have unique roles in melanoma progression and drug resistance.
Combining metabolic cell death with immune regulation and nanoparticle delivery improves therapeutic outcomes.
Key genes like SLC7A11, GPX4, and FDX1 are central to metabolic cell death mechanisms in melanoma.
Abstract
Melanoma is a highly aggressive malignant tumor arising from melanocytes, with its incidence and mortality rates continuously rising in recent years, posing a major global public health challenge. Although traditional targeted therapies and immune checkpoint inhibitors have significantly improved survival in some patients, primary and acquired resistance remain common, creating an urgent need for new treatment strategies. In recent years, metabolic cell death, ferroptosis, cuproptosis, and disulfidptosis, has shown unique advantages in melanoma research. Ferroptosis directly kills tumor cells through iron-mediated lipid peroxidation; cuproptosis relies on copper-induced mitochondrial protein aggregation to inhibit tumor proliferation; disulfidptosis arises from disulfide stress caused by glucose deprivation. This review provides a detailed analysis on the mechanisms and metabolic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
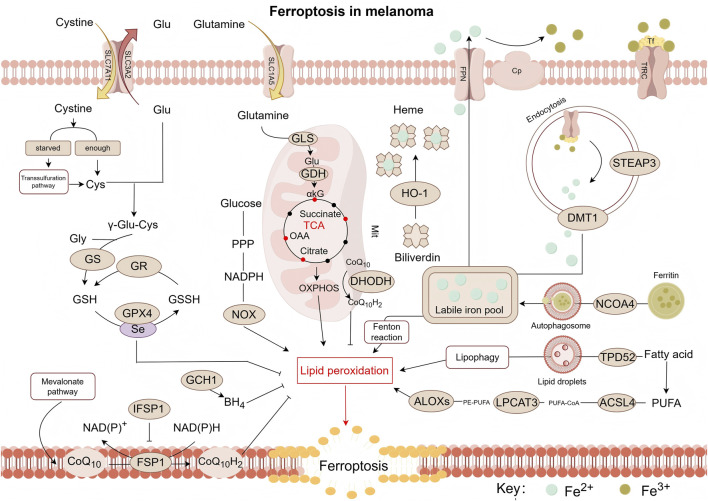 FIGURE 1
FIGURE 1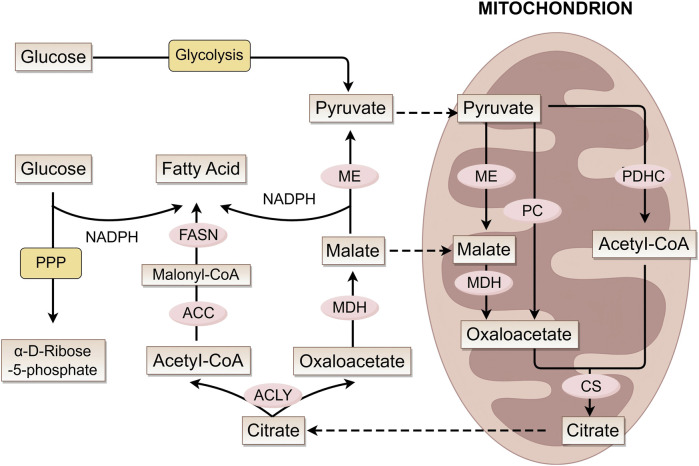 FIGURE 2
FIGURE 2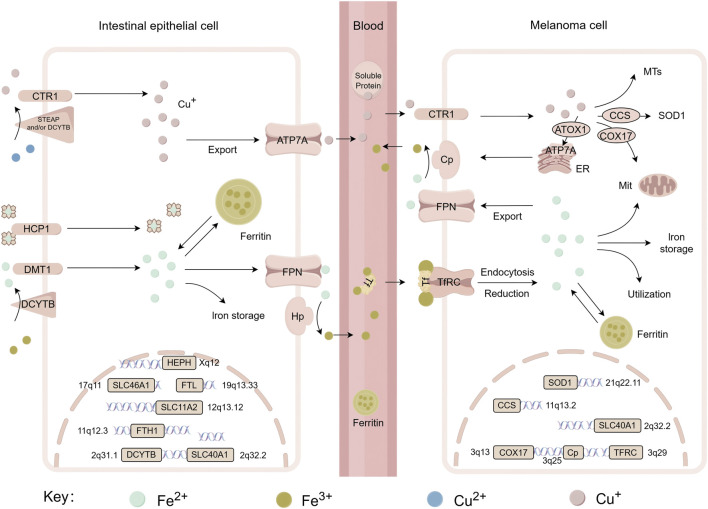 FIGURE 3
FIGURE 3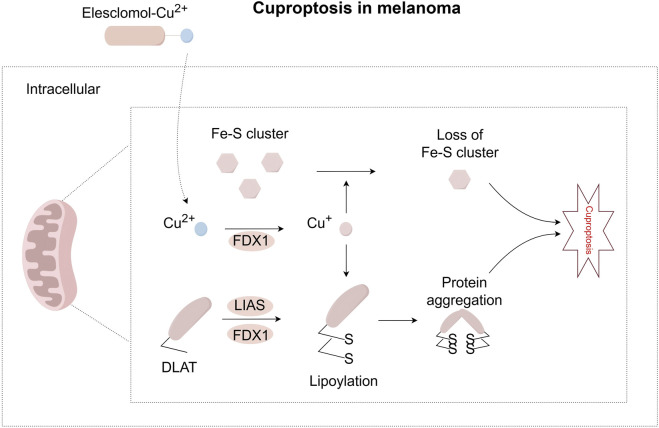 FIGURE 4
FIGURE 4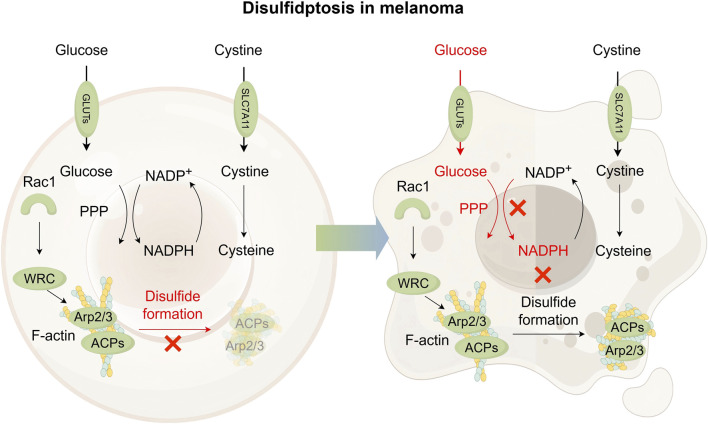 FIGURE 5
FIGURE 5| Feature | Ferroptosis | Cuproptosis | Disulfidptosis |
|---|---|---|---|
| Core Inducer | Iron ions (Fe2+) | Copper ions (Cu+) | Disulfide bonds |
| Morphological Features | Increased mitochondrial membrane density; Mitochondrial shrinkage; Reduction or disappearance of mitochondrial cristae; Outer membrane rupture ( | Mitochondrial shrinkage; Plasma membrane rupture; Endoplasmic reticulum damage ( | Cell membrane contraction; loss of contents; collapse of the F-actin network; Cytoskeleton collapse ( |
| Hallmark Metabolic Event | Lipid peroxidation of membrane phospholipid | Oligomerization of lipoylated TCA cycle proteins | Actin disulfide crosslinking |
| Key Defense Pathways | GPX4-GSH axis | Copper chaperones (ATOX1, CCS)Copper efflux pumps (ATP7A/B) | NADPH regeneration |
| Death Executor | Accumulation of lipid ROS → Loss of membrane integrity | Mitochondrial protein aggregation → Proteotoxic stress | F-actin contraction → Plasma membrane detachment |
| Category | Compound | Research Stage | Mechanism related to ferroptosis | Current application | Existing research on melanoma | Research material | |
|---|---|---|---|---|---|---|---|
| Ferroptosis | ACSL4 Indirect Regulation | Rosiglitazone | Clinical use | PPARγ agonist; indirectly inhibits ACSL4 expression ( | Type 2 diabetes | Promote the development of human melanoma in xenografts | A375 ( |
| Pioglitazone | Clinical use | PPARγ agonist; similar to Rosiglitazone ( | Type 2 diabetes | Interfer TLR4-dependent signaling pathways to exert protective effects against melanoma | B16F10 ( | ||
| LOX Inhibition | Zileuton | Clinical use | Inhibits LOX activity; reducing lipid peroxidation ( | Asthma treatment | — | — | |
| Antioxidant | Ferrostatin-1 | Preclinical | Radicals scavenging; iron chelation ( | Tool compound for mechanism studies | Inhibit melanoma ferroptosis | A375 ( | |
| Liproxstatin-1 | Preclinical | Clearing ROS; activates the Nrf2 pathway and restores GPX4 levels ( | Tool compound for mechanism studies | Inhibit melanoma ferroptosis | M21 ( | ||
| ROS Scavenging | PHOXNO | Preclinical | Nitrogen oxide; blocks Fenton reaction and ROS generation ( | Tool compound for mechanism studies | — | — | |
| Ferroptosis | Targeting | Erastin | Preclinical | Inhibits System Xc−; activates Nrf2/HO-1 pathway in cervical cancer cells ( | Tool compound for mechanism studies | Induce melanoma ferroptosis | A375 G361 ( |
| Sorafenib | Clinical use | Inhibits System Xc−; suppresses BXIP/SCD axis in HCC ( | Advanced HCC, RCC | Induce melanoma ferroptosis | B16F10 ( | ||
| Sulfasalazine (SAS) | Clinical use | Inhibits System Xc−; increases Fe2+ and ROS levels in Neuroblastoma with MYCN amplification ( | Inflammation | Decrease glutathione content, enhance susceptibility to radiation therapy | B16F10 ( | ||
| Targeting | RSL3 | Preclinical | Reduces GPX4 expression; increases ROS ( | Tool compound for mechanism studies | Induce melanoma ferroptosis | A375 G361 ( | |
| ML162 | Preclinical | Inhibits GPX4; induces ROS accumulation ( | Tool compound for mechanism studies | — | — | ||
| Targeting | iFSP1 | Preclinical | Inhibits FSP1; reduces CoQ10 synthesis ( | Tool compound for mechanism studies | — | — | |
| Simvastatin | Clinical use | Inhibit HMGR, block MVA pathway and CoQ10/GPX4 biosynthesis in Triple-negative breast cancer ( | Anti | Combine simvastatin with tanshinone Ⅰ to regulate the expression of PARP1 and inhibit melanoma | A375 ( | ||
| Targeting | Cisplatin | Clinical use | Depletes GSH | Solid tumors | Anti melanoma | Patients with melanoma | |
| Acetaminophen (APAP) | Clinical use | Metabolite NAPQI depletes GSH; induces ferroptosis in hepatocytes ( | Pain/fever | — | — | ||
| Targeting | Ferumoxytol | Clinical use | Increases Fe2+ levels ( | Anemia in chronic kidney disease | Enhance immune effect of macrophages on melanoma | B16F10 ( | |
| FINO2 | Preclinical | Oxidizes iron, induces lipid peroxidation ( | Tool compound for mechanism studies | — | — | ||
| Salinomycin | Preclinical | Induce iron loading in the lysosomes, induces ROS ( | Broad-spectrum anti-bacterial agent; | Obviate liver metastasis of uveal melanoma | 92.1 Mel270 | ||
| Cuproptosis Inhibitors | Targeting | Ammonium tetrathiomolybdate | Phase II clinical trials | Copper chelating agents are also a type of sulfide donor ( | Breast cancer | — | — |
| Penicillamine | Preclinical | Copper chelating agents ( | Tool compound for mechanism studies | Induce Noxa (PMAIP1)-dependent mitochondrial apoptosis | A375 G361 ( | ||
| Cuproptosis | Targeting | Disulfiram | Preclinical | The active form of DSF transports copper to intracellular compartments ( | Tool compound for mechanism studies | Induce copper-dependent reactive oxygen species stimulation | B16F10 ( |
| Elesclomol | Preclinical | Highly lipophilic copper ion carrier ( | Tool compound for mechanism studies | Load copper and induce cuproptosis | B16 ( | ||
| Zinc Pyrithione | Market exit | Copper ion carrier ( | Antibacterial action | Increase the zinc content within melanoma cells and induce autophagy | Primary culture ( | ||
| Disulfidptosis | Targeting | Erastin | Preclinical | Block the cystine uptake mediated by SLC7A11 ( | Tool compound for mechanism studies | Induce ferroptosis (the main function) | — |
| Targeting disulfide bond | DL-dithiothreitol | Preclinical | Maintain molecular structure ( | Tool compound for mechanism studies | — | — | |
| Disulfidptosis | Targeting | Diethyl maleate | Preclinical | Upregulate the activity of system Xc− ( | Tool compound for mechanism studies | — | — |
| Targeting glucose | BAY-876 | Preclinical | Glucose transporter 1 (GLUT1) inhibitor ( | Tool compound for mechanism studies | — | — | |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA regulation and disease · melanin and skin pigmentation · Ferroptosis and cancer prognosis
1 Introduction
Melanoma, a highly aggressive malignant tumor originating from melanocytes in the skin or other anatomical sites (e.g., ocular uvea, mucosa, meninges), has seen a persistent rise in global incidence and mortality, emerging as a major public health challenge. Epidemiological projections indicate 100,640 new cases and 8,290 deaths in the United States in 2024 (Long et al., 2023), with global incidence expected to 510,000 cases by 2040 (Ubellacker et al., 2020). Notably, the disease primarily affects Caucasian populations, with ultraviolet (UV) radiation exposure identified as the primary risk factor (Teixido et al., 2021). While early-stage localized melanoma is often curable through surgical resection (Tyrell et al., 2017), the prognosis for metastatic melanoma remains poor, with distant metastatic cases exhibiting a 5-year survival rate of less than one-third (Tao et al., 2025), underscoring the complexity of clinical management.
From a molecular pathological perspective, the aggressive nature of melanoma is closely linked to its mutation. Approximately 50%–60% of cases are linked to the BRAF V600E mutation (Anestopoulos et al., 2022; Marzagalli et al., 2019), together with other common mutations involving KRAS (Kirsten rat sarcoma viral oncogene homolog), NRAS (neuroblastoma RAS viral oncogene homolog), HRAS (Harvey rat sarcoma viral oncogene), CDKN2B (cyclin-dependent kinase inhibitor 2B), PTEN (phosphatase and tensin homolog), TERT (telomerase reverse transcriptase), and p53 (Ta et al., 2023). Especially, the metastatic potential of melanoma is associated with its unique metabolic adaptability: the lymphatic system promotes the distant dissemination of tumor cells by providing oxidative stress protection (Ubellacker et al., 2020). Additionally, the presence of immunosuppressive cells (e.g., regulatory T cells) in the tumor microenvironment (TME) and changes in the extracellular matrix are known to accelerate the progression of the disease (Xu et al., 2021).
Although targeted therapies and immunotherapies have significantly changed the treatment for advanced melanoma, their clinical applications still face multiple challenges. Compared with monotherapy, kinase inhibitors targeting BRAF mutations (e.g., vemurafenib, dabrafenib) combined with MEK inhibitors (e.g., trametinib) have improved median overall survival (OS) and progression-free survival (PFS) rates in patients with unresectable advanced metastatic BRAF-V600-mutant melanoma (Robert et al., 2019; Dhillon, 2016). Nevertheless, acquired resistance remains inevitable. 1mmune checkpoint inhibitors (ICIs), such as anti-PD-1 agents (pembrolizumab) and anti-CTLA-4 agents (ipilimumab), have achieved long-term survival in a subset of patients. However, a quite proportion of melanoma patients exhibit primary or acquired resistance (e.g., increased PD-L1 expression, impaired antigen presentation, or T-cell inactivation) resulting in 40%–65% failure in anti-PD-1 monotherapy and more than 70% failure in anti-CTLA-4 therapy (Gide et al., 2018). Additionally, chemotherapy and radiotherapy show limited effectiveness against metastatic melanoma and often lead to significant adverse effects.
Cell death is a fundamental biological process that maintains homeostasis in multicellular organisms. However, its dysregulation influences the pathogenesis and progression of diseases such as cancer (Hotchkiss et al., 2009). Traditionally, cell death is divided into two categories: non-regulated (accidental) cell death and regulated cell death. The former occurs passively due to external factors (e.g., physical injury or chemical toxicity) and lacks clear intracellular signaling pathways (Green and Victor, 2012). In contrast, regulated cell death is an orderly process governed by specific molecular programs. (Galluzzi et al., 2018; Tang et al., 2019).
Recent advances have revealed a distinct class of regulated cell death, which arises from metabolic imbalances caused by nutrient depletion or metal ion overload. This form of cell death–metabolic cell death–has often been referred to as “cell sabotage” (Green and Victor, 2012). It includes mechanisms like ferroptosis, cuproptosis, disulfidptosis, etc. (Mao et al., 2024). While the potential physiological roles of these “sabotage” mechanisms remain debated, a thorough analysis of their regulatory networks could lead to a better understanding of disease progression and the development of novel cancer therapies (Green and Victor, 2012). More and more emerging evidences have disclosed the pivotal role of copper, iron, and disulfide homeostasis in melanoma pathogenesis and therapeutic resistance. This review focuses on the molecular interplay in metabolic cell death including ferroptosis, cuproptosis and disulfidptosis. How dysregulation of these pathways leads to melanoma initiation, metastasis, and resistance to conventional therapies. We also evaluate their therapeutic potential, focusing on the interactions between metabolic stress and immune responses in melanoma, thereby providing critical insights for the development of novel therapeutic strategies.
2 Ferroptosis in melanoma: an iron-dependent cell death driven by lipid peroxidation
Thirteen years ago, Dixon’s lab proposed the concept of a unique form of regulated, iron-dependent cell death driven by lipid peroxidation, along with the term “Ferroptosis.” Three key research areas converged to establish the foundational understanding of the field of “ferroptosis”: (i) metabolic mechanisms, (ii) regulation of reactive oxygen species (ROS), and (iii) iron homeostasis (Stockwell, 2022), as illustrated in Figure 1.
Pathway map of mechanisms associated with ferroptosis in melanoma. Ferroptosis is a form of cell death triggered by lipid peroxidation, which primarily occurs on polyunsaturated fatty acid-containing phospholipids (PUFA-PLs). Notably, PE-PUFA exhibit high susceptibility to ferroptosis. The synthesis of PE-PUFA is regulated by enzymes such as ACSL4, LPCAT3, and ALOXs. Ferroptosis relies on iron-mediated Fenton reactions, making the process highly sensitive to alterations in iron metabolism. This involves multiple steps including iron uptake (e.g., via transferrin), storage (e.g., via ferritin), release (e.g., via DMT1), and efflux (e.g., via ferroportin). To counteract lipid peroxidation and prevent ferroptosis, cells primarily depend on the GPX4 pathway as well as GPX4-independent pathways, such as those mediated by CoQ10H2, DHODH, and BH4—which constitute key antioxidant defense mechanisms. ROS, reactive oxygen species; Glu, glutamic acid; Cys, cysteine; GS, glutamine synthetase; GR, glutathione reductase; GSH, glutathione (Reduced); GSSH:glutathione (Oxidized); GPX4, glutathione peroxidase 4; GCH1, GTP cyclohydrolase 1; BH4:tetrahydrobiopterin; CoQ10, coenzyme Q10; CoQ10H2, reduced coenzyme Q10; FSP1, ferroptosis suppressor protein 1; IFSP1, ferroptosis suppressor protein 1; NOX, NADPH oxidase; PPP, pentose phosphate pathway; αkG, α-ketoglutaric acid; OAA, oxaloacetic acid; HO-1, Heme oxygenase 1; ALOXs, arachidonate lipoxygenases; LPCAT3, lysophosphatidylcholine Acyltransferase 3; DHODH, dihydroorotate dehydrogenase; ACSL4, acyl-CoA synthetase long-chain family member 4; TPD52, tumor protein D52; NCOA4, nuclear receptor coactivator 4; PUFA, polyunsaturated fatty acid; PE, phosphatidylethanolamine; CoA, coenzyme A; DMT1, divalent metal transporter 1; STEAP3, six-transmembrane epithelial antigen of prostate 3; Cp, ceruloplasmin; Tf, transferrin; TfRC, transferrin receptor; FPN/SLC40A1, ferroportin; SLC3A2, recombinant solute carrier family 3, member 2; SLC7A11, recombinant solute carrier family 7, member 11; ASCT2/SLC1A5, solute carrier family 1, member 5; GLS:glutaminase; GDH:glutamate dehydrogenase.
2.1 Lipid metabolism and ferroptosis induction in melanoma
Ferroptosis is closely linked to dysregulated lipid metabolism in melanoma, in which polyunsaturated fatty acid (PUFA) metabolism serves as the central molecular mechanism. Figure 2 shows the process of fatty acid formation in melanoma cells. Mitochondrial acetyl-CoA combines with oxaloacetate to form citrate via citrate synthase (CS) (Koundouros and Poulogiannis, 2020; Chhimpa et al., 2023), exported via citrate carrier (CIC). Notably, during metabolic stress such as hypoxia, the synthesis of acetyl-CoA preferentially originates from acetate. And melanoma, particularly brain metastases with poor prognosis, exhibits increased dependency on above-mentioned acetate, a weakness specific to BRAF-mutant tumors (Kamphorst et al., 2014; Mashimo et al., 2014; Lakhter et al., 2016). Cytosolic ATP-citrate lyase (ACLY) cleaves citrate into acetyl-CoA and oxaloacetate. Subsequent oxaloacetate conversion to malate (via MDH) and pyruvate (via ME) generates NADPH for biosynthesis (Simmen et al., 2020).
Fatty acid metabolic pathways and key enzymes. This figure illustrates the interconnected mechanisms of fatty acid synthesis, mitochondrial metabolism (involving the conversion of intermediate metabolites such as pyruvate, acetyl-CoA, and citrate), and glucose metabolic pathways. It demonstrates how these distinct metabolic processes are linked through key metabolites and enzymes (e.g., ME, FASN, ACLY), working in concert to coordinately regulate cellular material and energy metabolism. ME, malic enzyme; MDH, malate dehydrogenase; PC, pyruvate carboxylase; PDHC, pyruvate dehydrogenase complex; CS, citrate synthase; ACC, acetyl-CoA carboxylase; FASN, fatty acid synthase; ACLY, ATP citrate lyase.
Fatty acid (FA) synthesis initiates with acetyl-CoA carboxylase (ACC) producing malonyl-CoA. Then, the fatty acid synthase (FASN) complex iteratively elongates the chain to produce palmitate (C16:0) (Wedan et al., 2024). Li and Kapur et al. discovered that ACC and FASN are significantly upregulated in human melanoma (Kapur et al., 2005; Li et al., 2016). Long-chain fatty acid (LCFA) elongation occurs mainly in the endoplasmic reticulum (ER), with desaturation being mediated by stearoyl-CoA desaturase (SCD) (Kubota and Espenshade, 2022). Research indicates that SCD activity may play a key role in melanoma phenotype switching: under low SCD conditions, ER stress is induced, suppresses microphthalmia-associated transcription factor (MITF), and promotes melanoma dedifferentiation (Vivas-García et al., 2020).
In melanoma ferroptosis, the oxidative modification of polyunsaturated PUFAs incorporated into membrane phospholipids plays a decisive role. Under oxidative stress, Reactive Oxygen Species (ROS) selectively attack hydrogen atoms from PUFA double bonds, triggering lipid peroxidation chain reactions (Yan et al., 2021). Studies reveal that phosphatidylethanolamine (PE)-bound PUFAs, particularly PE-AA and PE-AdA, exhibit marked sensitivity to ferroptosis (Fang et al., 2023), with their abundance positively correlating with acyl-CoA synthetase long-chain family member 4 (ACSL4) activity. Meanwhile, ACSL3, which belongs to the same family as ACSL4, is also associated with the poor prognosis of melanoma (Chen et al., 2016). One study indicates that the abundant oleic acid in lymphocytes protects melanoma cells from ferroptosis in an ACSL3-dependent manner. Notably, enzymatic oxidation of PUFAs is catalyzed by lipoxygenases (LOXs, particularly 15-LOX1/2) (Bouchaoui et al., 2023). This process is antagonized by ferroptosis inhibitors such as glutathione peroxidase 4 (GPX4), forming a key regulatory node that maintains redox balance in ferroptosis control.
2.2 GPX4 and its role in suppressing ferroptosis in melanoma
ROS, which include hydrogen peroxide (H_2_O_2_), hydroxyl radicals (·OH), singlet oxygen (^1^O_2_), and superoxide (·O_2_ ^−^), among others (Bedard and Krause, 2007), function as key signaling mediators in redox processes and are critically involved in both the initiation and progression of ferroptosis. The mitochondrial electron transport chain is considered major contributor to ROS generation. During coenzyme Q10 (CoQ_10_) mediated electron transfer, approximately 1%–2% of electrons leak during their transport from Complex I and II to Complex III (Gutierrez-Mariscal et al., 2021). Under specific conditions, these leaked electrons react with molecular oxygen to generate superoxide radicals (·O_2_ ^−^) (Parascandolo and Laukkanen, 2019), later converted to H_2_O_2_ by SOD. H_2_O_2_ can then react with free Fe^2+^ through the Fenton reaction, yielding highly reactive hydroxyl radicals (·OH). In BRAF-mutant melanoma, BRAF inhibitors upregulate oxidative phosphorylation (OXPHOS), enhancing mitochondrial ROS accumulation and cellular susceptibility to ferroptosis (Haq et al., 2013; Schöckel et al., 2015). Additionally, the NADPH oxidase (NOX) family specifically catalyzes the generation of O_2_ ^−^ via transmembrane electron transfer. NOX2 activation stimulates ROS production through a calcium signaling pathway dependent on the Ryanodine receptor (RyR) (Prosser et al., 2011).
The cystine/GSH/GPX4 axis represents a central defense mechanism against ferroptosis. System Xc^-^, which is a heterodimeric transmembrane transporter consisting of the light chain xCT (SLC7A11) and heavy chain 4F2hc (SLC3A2) (Koppula et al., 2018), facilitates the exchange of cystine and glutamate at a 1:1 ratio (Liu et al., 2021). It has been observed that BRAF inhibitor-resistant melanomas display increased reliance on glutamine and activate the NRF2 pathway, increasing xCT expression and GSH levels to evade ferroptosis (Khamari et al., 2018). Conversely, dedifferentiated melanomas are characterized by lower basal GSH levels and greater susceptibility to ferroptosis (Tsoi et al., 2018). Cystine within the cells is reduced to cysteine for glutathione (GSH) biosynthesis. GSH exists in reduced (GSH) and oxidized (GSSG) forms, dynamically balanced by glutathione peroxidase 4 (GPX4) and glutathione reductase (GR) (González-Domínguez et al., 2020). Among mammalian GPX family members (GPX1-GPX8), GPX4 is uniquely capable of clearing membrane lipid hydroperoxides. GSH biosynthesis is regulated in a stepwise manner: γ-glutamylcysteine ligase (GCL) catalyzes the conjugation of glutamate and cysteine to form γ-glutamylcysteine (γ-Glu-Cys) (Oppenheimer et al., 1979), followed by glycine addition via glutathione synthetase (GS) to produce GSH (Gasmi et al., 2024). In addition to System Xc^−^-mediated cystine uptake, mammalian cells utilize the transsulfuration pathway to derive cysteine from methionine. Studies indicate that elevated tumor microenvironmental methionine levels may disrupt GSH homeostasis through this pathway (Kamphorst et al., 2015).
2.3 Iron regulation and sensitivity to ferroptosis in melanoma
Iron, as an essential trace metal element in living organisms, participates in a wide range of critical biochemical processes, including oxygen transport, DNA synthesis and repair, and electron transfer in mitochondria (Galaris et al., 2019). From a chemical perspective, iron exhibits valence states ranging from −2 to +7. However, biological systems mainly utilize the +2 [ferrous, Fe (II)] and +3 [ferric, Fe (III)] redox states (Bayır et al., 2023). In melanoma, iron metabolism drives ferroptosis through multiple pathways. Heme iron is taken up by enterocytes via heme carrier protein 1 (HCP1) on their apical membrane as an intact porphyrin complex. Conversely, absorption of non-heme iron requires reduction of Fe^3+^ to Fe^2+^, which is catalyzed by duodenal cytochrome B (DCYTB), followed by transport across the membrane through divalent metal transporter 1 (DMT1/SLC11A2) (Dutt et al., 2022). Figure 3 illustrates the iron transport process, while the copper transport mechanism will be detailed in the next chapter.
Systemic transport of iron and copper, related genes and their chromosomal localization. This figure shows the metabolic processes of iron (Fe) and copper (Cu) in intestinal epithelial cells and melanoma cells, including the uptake, transport, and storage of metal ions, as well as the roles of associated functional proteins such as CTR1, DMT1, Ferritin and so on. It also displays the chromosomal locations of key genes involved in these metabolic pathways—including SLC46A1, FTL, SOD1, etc., thereby revealing the molecular mechanisms and regulatory gene networks underlying copper and iron metabolism across different cell types. DCYTB, duodenal cytochrome b; DMT1/SLC11A2, divalent metal transporter 1; HCP1/SLC46A1, heme carrier protein 1 (HCP1); STEAP, six-transmembrane epithelial antigen of prostate; CTR1/SLC31A1, copper transport protein 1; Hp/HEPH, hephaestin; FPN/SLC40A1, ferroportin; Cp, ceruloplasmin; FTL, ferritin light chain; FTH, ferritin heavy chain 1; TFRC, transferrin receptor; ATOX1, antioxidant 1; CCS, copper chaperone for superoxide dismutase; COX17, cytochrome c oxidase copper chaperone; SOD1, superoxide dismutase 1; ATP7A, copper-transporting ATPase 1; MTs, metallothioneins.
Iron efflux from intestinal epithelial cells depends on the coordinated activity of ferroportin (FPN/SLC40A1) and hepcidin (Hp) at the basolateral membrane. Iron is extruded into the extracellular space by FPN, after which it is oxidized to Fe^3+^ by Hp, enabling its binding to transferrin (TF). The transferrin-iron complex in plasma is internalized by target cells via transferrin receptor (TfR)-mediated endocytosis (Tang et al., 2021). This process involves vesicular acidification, iron dissociation, reduction of Fe^3+^ catalyzed by STEAP3 (six-transmembrane epithelial antigen of the prostate 3), and final transport into the cytoplasm via DMT1.
In melanoma, the labile iron pool (LIP) expansion promotes Fenton reactions, which produce hydroxyl radicals that initiate lipid peroxidation, a key feature of ferroptosis. Excess iron is normally stored by ferritin, but ferritin degradation through ferritinophagy releases iron back into the pool, accelerating ferroptosis in melanoma (Gao et al., 2016). Crucially, iron regulatory protein 1 (IRP1) is upregulated by ferroptosis inducers (e.g., erastin, RSL3) in A375 and G361 melanoma cells, promoting ferroptosis by modulating TfR, FPN to increase intracellular iron (Yao et al., 2021).
Iron metabolism within the skin also plays a critical role in melanoma development. Disruption of iron balance induces oxidative stress in the skin microenvironment, accelerating inflammation and melanoma risk. Iron lack impairs skin metabolism, while excess iron promotes ferroptosis through ROS accumulation, representing a targetable weakness in melanoma treatment.
2.4 Alternative antioxidant pathways in ferroptosis resistance
Recent advancements in ferroptosis regulation research have revealed multiple novel antioxidant defense systems independent of the classic GSH-GPX4 pathway, offering new targeted strategies for cancer therapy. Non-GPX4 dependent pathways also play crucial roles in ferroptosis resistance, together constituting a defense system that operates across multiple cellular compartments.
Ferroptosis suppressor protein 1 (FSP1), primarily localized in lipid droplets and plasma membranes (Bersuker et al., 2018), acts as a key regulatory factor independent of the glutathione system. It uses NAD(P)H to reduce ubiquinone (CoQ_10_) to ubiquinol (CoQ_10_H_2_), which can directly neutralize lipid radicals and thus suppress lipid peroxidation (Bersuker et al., 2019; Doll et al., 2019). Emerging evidence indicates FSP1 can also exert ferroptosis resistance through mechanisms not involving CoQ10 by promoting membrane repair via the ESCRT-III complex (Dai et al., 2020). The STARD7 protein controls both the production and spatial distribution of CoQ10: its mitochondrial form synthesizes CoQ_10_, while the cytosolic form delivers it to various membranes, helping to form a trans-membrane antioxidant system (Deshwal et al., 2023).
Within mitochondria, DHODH and SQRDL constitute another protective layer. DHODH, a key enzyme in the pyrimidine synthesis pathway, reduces mitochondrial CoQ_10_ to CoQ_10_H_2_ and collaborating with mitochondrial GPX4 to combat mitochondrial specific lipid peroxidation. When GPX4 activity is compromised, DHODH maintains mitochondrial redox homeostasis through compensatory upregulation (Mao et al., 2021). SQRDL utilizes selenite as an electron donor to catalyze mitochondrial CoQ_10_ reduction. This pathway not only participates in antioxidant effects but also modulates ferroptosis susceptibility through mitochondrial electron transport chain regulation (Zarrinpar, 2022; Gui et al., 2021). It has been experimentally shown that combined inhibition of both DHODH and mitochondrial GPX4 induces extensive mitochondrial lipid peroxidation and irreversible ferroptosis, underscoring the functional synergy within this dual mitochondrial antioxidant system.
Furthermore, GCH1 (GTP cyclohydrolase 1), which is the rate-limiting enzyme for synthesizing tetrahydrobiopterin (BH4), inhibits ferroptosis through GPX4-independent mechanisms. As a potent free radical scavenger (Soula et al., 2020; Kraft et al., 2020), BH4 directly neutralizes lipid peroxidation radicals. In tumor cells lacking GPX4, the GCH1-BH4 pathway is often activated as a compensatory survival mechanism, becoming essential for maintaining cell viability.
3 Cuproptosis in melanoma: a mitochondrial damage activated by copper overload
In 2022, Tsvetkov et al. discovered a new type of regulated cell death that depends on copper and is induced by mitochondrial protein aggregation, which they named “cuproptosis”.
3.1 Dysregulated copper homeostasis promotes cuproptosis in melanoma
Both Systemic and cellular copper levels are tightly controlled to prevent toxicity. Dietary Cu^2+^ is reduced to Cu^+^ by STEAP or DCYTB and then imported through the high-affinity copper transporter CTR1 (SLC31A1) (Figure 3). CTR1 is overexpressed in melanoma biopsies compared to normal tissue (Mason, 1979; Lv et al., 2022; Georgatsou et al., 1997). DMT1 provides an alternative uptake when CTR1 is insufficient. Inside the cell, copper distribution is guided by specific chaperones: COX17 delivers copper to mitochondria (Horng et al., 2004; Cobine et al., 2006; Banci et al., 2008), CCS supplies copper to superoxide dismutase 1 (SOD1) for redox defense (Bertinato and L'Abbé, 2003; Prohaska et al., 2003), and ATOX1 is responsible for transferring copper to Cu-ATPases (ATP7A/B) in the trans-Golgi network (TGN) and supports the synthesis of cuproenzymes such as ceruloplasmin (Hamza et al., 2003). Meanwhile, the Cu-ATPases ATP7A (ubiquitous) and ATP7B (liver-specific) also act as the major transporters for exporting cellular copper (Lutsenko et al., 2007). Excess copper is stored by metallothioneins (MTs) or removed from the cell primarily through ATP7A and ATP7B (La Fontaine and Mercer, 2007; Palmgren and Nissen, 2011). Importantly, disruptions in copper export or storage trigger cytotoxic copper accumulation, directly linking cuproptosis in melanoma cells (Schmidt et al., 2018; Tsvetkov et al., 2022). Copper chelating active substance (e.g., D-penicillamine) induce phorbol-12-myristate-13-acetate-induced protein 1 (PMAIP1) expression, which upregulates NOXA protein, a necessary condition for melanoma cell death, highlighting the potential of targeting copper homeostasis as a therapeutic strategy (Qiao et al., 2012).
3.2 FDX1 and lipoylated proteins in cuproptosis activation
Distinct from ferroptosis, cuproptosis is directly triggered by copper overload and exhibits unique resistance to classical cell death inhibitors (Tsvetkov et al., 2022). Melanoma shows particular sensitivity to copper toxicity due to its metabolic characteristics: 35%–50% of wild-type, BRAF-mutant, and patient-derived melanoma cells rely heavily on oxidative phosphorylation (OXPHOS) for energy, making them sensitive to copper-induced mitochondrial damage (Fischer et al., 2018). The mechanism of cuproptosis is closely tied to mitochondrial function, as it can be suppressed via electron transport chain inhibitors (Tsvetkov et al., 2022). A key player is ferredoxin 1 (FDX1), which is highly expressed in melanoma and other cancers (Lu et al., 2023). FDX1 reduces Cu^2+^ to more toxic Cu^+^ and activates mitochondrial protein lipoylation by interacting with lipoic acid synthetase (LIAS) (Dreishpoon et al., 2023). Lipoylation, a post-translational modification, be disrupted during cuproptosis (Tsvetkov et al., 2022; Rowland et al., 2018). Notably, copper binds directly to lipoylated DLAT, triggering its oligomerization. This is thought to produce toxic protein aggregates that lead to cell death (Tsvetkov et al., 2022). Interestingly, high FDX1 expression in melanoma is linked to improved response to anti-PD-L1 immunotherapy (Lu et al., 2023), but FDX1 knockdown inhibits the in vitro proliferation of melanoma cells (Liu et al., 2022). Copper toxicity also damages iron-sulfur (Fe-S) cluster integrity. Recent studies have found that treating cells with a copper ionophore resulted in FDX1-dependent loss of Fe-S cluster proteins (Walshe, 2007), Figure 4 visually presents this process.
Pathway map of mechanisms associated with cuproptosis in melanoma. Extracellular copper enters cells through copper ionophores, such as elesclomol. Inside the cell, FDX1 reduces Cu2+ to Cu+. FDX1 also works with LIAS to support the lipoylation of certain metabolic enzymes, such as DLAT. When Cu+ binds directly to lipoylated proteins, it causes proteins to form oligomers. This abnormal process can lead to a toxic function that results in cuproptosis. At the same time, Cu+ can lead to the loss of iron-sulfur clusters in a process dependent on FDX1. These combined effects cause proteotoxic stress and eventually lead to cell death. FDX1, Ferredoxin 1; LIAS, Lipoic Acid Synthetase.
4 Disulfidptosis in melanoma: a novel cell death mechanism triggered by disulfide stress
Disulfidptosis, identified in 2023, is a type of regulated cell death that occurs in cells with high SLC7A11 expression when glucose is scarce. Upon glucose starvation, cells with high expression of SLC7A11 experience rapid NADPH depletion and abnormal accumulation of insoluble disulfides, leading to disulfidptosis, as shown in Figure 5 (Liu et al., 2020; Goji et al., 2017; Liu X. et al., 2023; Joly et al., 2020). Importantly, this death mechanism differs from apoptosis, necroptosis and ferroptosis, as evidenced by insensitivity to their inhibitors and absence of classical markers like caspase-3 cleavage, cystine crystal formation or ATP depletion (Liu X. et al., 2023; Elmonem et al., 2016; Pereira et al., 2015). Cystine removal rescues these cells from glucose starvation-induced death (Liu et al., 2020; Goji et al., 2017), while thiol oxidizing agents worsen it (Liu X. et al., 2023). The glycolysis inhibitor 2-deoxyglucose (2DG) unexpectedly reduces cell death by shifting glucose analogs into the PPP to replenish NADPH (Liu et al., 2020; Zhang et al., 2014), confirming that NADPH supply not glycolytic is the key determinant. Disulfide-reducing agents (e.g., N-acetyl cysteine, tris (2-carboxyethyl) phosphine) restore NADPH levels and prevent cell death, further supporting disulfide overload as the main cause (Liu et al., 2020).
Pathway map of mechanisms associated with disulfidptosis in melanoma. In cells exhibiting high SLC7A11 expression, cystine uptake is significantly elevated, followed by its reduction to cysteine. This reduction process depends on NADPH as a key reducing agent, which is primarily supplied through the pentose phosphate pathway (PPP) using glucose. Abundant glucose is essential to prevent abnormal intracellular cystine accumulation and avoid disulfide bond formation within actin cytoskeleton proteins, thereby supporting cell survival in SLC7A11-high contexts. Under glucose-deficient conditions, limited NADPH availability leads to excessive buildup of cystine and other disulfide species. This promotes aberrant disulfide bonding in actin cytoskeletal proteins, resulting in the disruption of the actin network and ultimately inducing disulfidptosis. It is suggested that Rac1-WRC-Arp2/3-mediated branched actin polymerization may provide a structural environment conducive to disulfide bond formation among actin filaments, thereby promoting disulfidptosis. GLUT, glucose transporter; Rac1, Rac family small GTPase 1; ACPs, actin cytoskeleton proteins; WRC, Rac1-wave regulatory complex.
ROS scavengers show different effectiveness among various cell lines, indicating that ROS may influence but not initiate disulfidptosis (Liu et al., 2020; Joly et al., 2020). Further mechanistic insights reveal that glucose-starved cells with high expression of SLC7A11 leads to abnormal disulfide bonding within actin cytoskeleton proteins (ACPs), which results in the breakdown of the actin network, detachment of the plasma membrane, and ultimately disulfidptosis cell death (Liu D. et al., 2023). The Rac1-WAVE regulatory complex (WRC) pathway plays a critical role by activating Arp2/3-mediated branched actin polymerization and lamellipodia formation. This branched structure provides a site for disulfide bonding. Disulfidptosis is diminished upon WRC disruption and enhanced by Rac1 activation (Liu D. et al., 2023; Ibarra et al., 2005; Alekhina et al., 2017).
Correlatively, the movement and invasion of melanoma cells strongly rely on changes in the actin cytoskeleton. Although the WRC-Arp2/3 pathway is a main controller of this process, melanoma cells show a high level of flexibility. The co-activator YAP, which promotes melanoma growth and spread, controls the Arp2/3 subunit ARPC5. This subunit is essential for keeping focal adhesions stable and supporting the ability of melanoma cells to invade (Lui et al., 2022). This YAP-ARPC5 axis represents an alternative pathway influencing Arp2/3 activity and actin dynamics in melanoma. In studies using B16F1 melanoma cells, it was found that lamellipodia-like structures (LLS) driven by Arp2/3 and regulated by Rac/Cdc42 can form without the WRC. This suggests there is more complexity in how the actin network is controlled in melanoma cells (Kage et al., 2022). These results indicate that melanoma cells may have other or backup ways to reshape their actin structures, which might change how they respond to disulfidptosis caused by WRC activation.
5 Interactions between three metabolism-linked cell death pathways
Ferroptosis, cuproptosis, and disulfidptosis are programmed cell death pathways triggered by metabolic stress. They form a dynamic antagonistic-synergistic network through shared key molecular hubs. Table 1 provides a comparative analysis of their characteristics, while Table 2 lists common or potential inducers and inhibitors for these three forms of programmed cell death.
5.1 The metabolic resource competition paradox among ferroptosis, cuproptosis, and disulfidptosis
Ferroptosis and disulfidptosis exhibit competitive regulatory features in NADPH generation and metabolic control. Ferroptosis relies on the enrichment of GSH precursors by system Xc^−^ and the antioxidant capacity of the GPX4-GSH system. In contrast, disulfidptosis arises from an imbalance between cystine intake driven by SLC7A11 and the use of NADPH. When glucose is limited, the PPP is unable to provide enough NADPH, which affects the activity of GPX4. The competition for metabolic resources between these two forms of cell death is notable. Specifically, high levels of SLC7A11 increase resistance to ferroptosis but also increase the risk of disulfidptosis due to excessive NADPH depletion. In addition, GPX4 also functions as an intracellular copper chelator to control copper levels inside the cell. Meanwhile, research has been shown that glutathione depletion induced by buthionine sulfoximine sensitizes cancer cells to cuproptosis (Mao et al., 2024; Tsvetkov et al., 2022). This competition at the metabolic level may help cells maintain a balance for cell fate selection.
5.2 Mitochondrial damage and disrupted metal homeostasis in ferroptosis, cuproptosis, and disulfidptosis
The interaction between cuproptosis and disulfidptosis mainly occurs at key points in mitochondrial metabolism. Copper ions induce abnormal aggregation of lipoylated DLAT in mitochondria via the FDX1-LIAS pathway, while NADPH deficiency in disulfidptosis may worsen mitochondrial function, forming a harmful cycle. The copper release mechanism involving ATP7A/B in cuproptosis shares similarities with the iron export process controlled by FPN in ferroptosis. At the same time, the stability of iron-sulfur clusters inside the cell affects whether cuproptosis is triggered, while ferroptosis also relies on Fe-S cluster biogenesis (Lee and Roh, 2023). This suggests that interfering with metal transport systems could impact multiple types of cell death simultaneously.
From an evolutionary perspective, the redox reactions involving metal ions naturally conflict with the precise control of cell metabolism especially in the tumor environment. To support rapid growth, cancer cells often accumulate metal ions in excess, but this makes them more sensitive to the activation of multiple death signals.
5.3 Molecular crosstalk among tumor microenvironment, ferroptosis, cuproptosis, and disulfidptosis
Ferroptosis, cuproptosis and disulfidptosis share certain common features in immune regulation, yet each exhibits distinct biological characteristics and mechanisms of action. Within the tumor microenvironment (TME), they collectively influence anti-tumor immune responses through metabolic reprogramming, immune cell polarization, and regulation of immune checkpoint expression.
The interaction between ferroptosis and the TME is complex. The sensitivity of immune cells to ferroptosis varies greatly. CD8^+^ T cells play a crucial role in triggering ferroptosis in cancer cells: IFNγ produced by CD8^+^ T cells suppresses the Xc^−^ system and upregulates ACSL4 expression, promoting ferroptosis in tumor cells (Dixon et al., 2012; Wang et al., 2019; Liao et al., 2022). However, some studies indicate that T cells themselves are also vulnerable to ferroptosis. Tumor-associated macrophages (TAMs) exhibit high plasticity and can differentiate into either immunostimulatory M1 or immunosuppressive M2 phenotypes. M2 macrophages, due to their lower antioxidant capacity, are more possible to ferroptosis than M1 macrophages (Kapralov et al., 2020; Luo et al., 2021). Inducing ferroptosis in M2 macrophages alleviates the immunosuppressive microenvironment, and modulating the M1/M2 macrophage ratio enhances the responsiveness to PD-1 therapy (Jiang et al., 2021), thereby improving the efficacy of cancer immunotherapy. NK cells also play a central role in anti-tumor immunity. NK cells inactivation in the TME are associated with oxidative stress, and activation of the transcription factor NRF2 has been shown to restore NK cells function (Sun et al., 2016; Poznanski et al., 2021). As the most potent antigen-presenting cells, dendritic cells (DCs) are essential for activating naive T cells and initiating T cell (Marmonti et al., 2022). Both the GPX4 inhibitor RSL3 and the lipid peroxidation product 4-hydroxynonenal (4-HNE) can impair dendritic cells (DCs) function (Cubillos-Ruiz et al., 2015). The effect of ferroptotic cancer cells on DCs may depend on the stage of ferroptosis. In early stages, cancer cells promote DCs maturation; however, as ferroptosis advances, this ability declines. Nonetheless, these late-stage ferroptotic cells remain susceptible to efficient phagocytosis by DCs (Efimova et al., 2020).
Cuproptosis modulates immune responses through copper-dependent mechanisms. Elevated copper levels in tumor tissues not only promote tumor proliferation and angiogenesis (Garber, 2015; Ge et al., 2022) but also specifically upregulate PD-L1 expression (Zhang et al., 2024), promoting immune escape. The copper transporter CTR1 is associated with infiltration of multiple immune cells (Wang J. et al., 2023) and correlates with poor prognosis in cancers such as breast cancer and melanoma (Lv et al., 2022; Wu et al., 2023). Copper chelators can reverse copper-induced immunosuppression and enhance infiltration of CD8^+^ T and NK cells (Cheng et al., 2022).
As a newly identified form of cell death, the direct link between disulfidptosis and the immune microenvironment remains unclear. However, its metabolic basis overlaps significantly with immune regulation. Key metabolites such as glucose, lactate, and cystine not only influence the occurrence of disulfidptosis but also strongly modulate immune cell function (Mi et al., 2024). For instance, high glucose consumption by tumor cells leads to nutrient deprivation in the TME and accumulation of lactate, which acidifies the TME. This acidic environment impairs T cell cytolytic activity and cytokine production (Apostolova and Pearce, 2022), while also promoting macrophage polarization toward the M2 phenotype and altering regulatory T cell (Treg) metabolism to sustain their function under low glucose conditions (Watson et al., 2021; Han et al., 2023). Furthermore, tumor cell expression of GLUT1 negatively correlates with CD8^+^ T cell infiltration. Inhibiting GLUT1 may not only induce disulfidptosis but also increase CD8^+^ T cell infiltration and reduce PD-L1 levels (Singer et al., 2011). Cystine deficiency similarly destroys T cell function (Wang et al., 2019), suggesting that modulating metabolite levels may together induce disulfidptosis and remodel the immune microenvironment.
These three forms of metabolic cell death interact with the TME through the release of and response to immune signaling molecules, as well as via metabolic competition. They share common involvement in metabolic reprogramming and immune checkpoint regulation, and all hold potential for combination with immunotherapy. Their distinctions lie in the fact that ferroptosis is closely linked to immune cell regulation, cuproptosis depends more on metal ion homeostasis and specific protein expression, while disulfidptosis is more intimately associated with glycolytic pathways. Current understanding of their immune effects is largely inferred from metabolic substrate competition, and specific mechanisms need further verification.
6 Therapeutic prospects of using metabolic cell death in melanoma
A complex network of molecular interactions and conflicting resource among ferroptosis, cuproptosis, and disulfidptosis creates multiple weakness in melanoma cells. However, effectively using these death mechanisms requires understanding the metabolic strategies of melanoma cells, especially when facing treatment resistance.
6.1 Strategies to reverse ferroptosis resistance in melanoma
Ferroptosis regulation in melanoma is closely related to changes in metabolism that lead to treatment resistance. Studies reveal that the unique lipid microenvironment of the lymphatic system protects metastatic melanoma cells from ferroptosis. Compared to blood, lymphatic fluid contains more glutathione and oleic acid. Oleic acid increases resistance to ferroptosis and supports the spread of cancer cells by activating the ACSL3-related signal (Ubellacker et al., 2020). BRAF inhibitor-resistant melanoma cells show changes in lipid metabolism, with lower levels of saturated fatty acids and higher levels of mono/polyunsaturated fatty acids. Targeting cholesterol esterification enzymes ACAT2 or SOAT helps regain drug sensitivity (Vergani et al., 2022). Transcriptional networks further enforce ferroptosis resistance in melanoma: EZH2 copy number amplification silences KLF14, upregulating SLC7A11 to enhance glutathione synthesis (Du et al., 2024), while APOE reduces polyunsaturated fatty acids and upregulates GPX4 (More et al., 2024). Resistance is similarly mediated by SREBP2-induced transferrin transcription, which reduces intracellular iron pools, ROS, and lipid peroxidation (Hong et al., 2021). These metabolic changes create a safe environment, allowing melanoma cells to avoid ferroptosis. Propafenone promotes mitochondrial HMOX1 expression by activating JNK/JUN signal, induces iron accumulation and ROS eruption, works together with RSL3 to promote ferroptosis in melanoma, Similarly, the CDK4/6 inhibitor palbociclib enhances the efficacy of the ferroptosis inducer auranofin by inducing cell senescence and depleting glutathione (GSH) and NADPH, This suggests that metabolic interventions as a strategy to overcome melanoma heterogeneity (Fan et al., 2024).
6.2 Amplify the therapeutic effect of melanoma by combined targeting of ferroptosis and immune activation
Multiple nanoplatforms combine the two functions of metabolic control and immune modulation. TPL@TFBF releases Fe^3+^ for Fenton reactions while inhibiting Nrf2, triggering ferroptosis/pyroptosis and releasing DAMPs to enhance CD8^+^ T cell infiltration (Wang S. et al., 2024). Similarly, the TCFI nanoplatform combines photodynamic therapy with ferroptosis to induce immunogenic cell death. Concurrently, interferon-γ secretion suppresses system xc^−^ activity, forming a positive environment between ferroptosis and antitumor immunity (Hou et al., 2023).
Post-translational modifications also provide potential targets to overcome resistance. Studies show that the balance between SUMOylation and phosphorylation of STAT1 is crucial for ferroptosis. A 108-amino acid polypeptide from circular RNA circPIAS1 interacts with the SUMO E3 ligase Ranbp2 to increase STAT1 SUMOylation, which in turn reduces its phosphorylation. This modification blocks the SLC7A11/GPX4 pathway and weakens interferon-γ-induced ferroptosis, ultimately leading to resistance against immune checkpoint inhibitors (ICB). This mechanism supports the idea of combining anti-circPIAS1 and PD-1 inhibitors in melanoma treatment (Zang et al., 2024). Another study identifies that the ubiquitin ligase Nedd4 induces resistance to ferroptosis by promoting VDAC2/3 degradation, a process inversely regulated by the FOXM1-Nedd4-VDAC2/3 (Yang et al., 2020). These research above suggest that using ferroptosis inducers together with agents that disrupt redox balance or promote immunogenic cell death can create combination therapies.
6.3 Innovative strategies for cuproptosis and disulfidptosis in melanoma treatment and the prognostic value of related genes
Cuproptosis and disulfidptosis are recently identified forms of metabolic cell death, but they show strong potential for melanoma therapy. Nanotechnology platforms are being developed to trigger tumor cell cuproptosis or disulfidptosis, aiming to improve treatment outcomes and solve shortcomings of traditional methods. For example, the nanocarrier ACM@MCHS-CuMOF@Dox, combining Mesoporous Carbon Hollow Spheres (MCHS) loaded with Copper-based Metal-Organic Frameworks (CuMOFs) and Doxorubicin (Dox), can downregulate FDX1 to induce apoptosis and cuproptosis, significantly inhibiting proliferation and migration of melanoma A375 cell line in vitro (Zhang et al., 2025); the triboelectric-field cuproptosis induction patch (TIP) utilizes a portable electric field to induce cuproptosis, overcoming limitations of traditional electrostimulation and effectively inhibiting postoperative melanoma recurrence (Chen et al., 2025). Another study developed a copper oxide nanoplateform (ES@CuO), which is absorbed by tumor cells and degrades to release Cu^2+^ triggering cuproptosis, significantly inhibiting B16 melanoma growth in mice, while promoting CD8^+^T cell infiltration and inflammatory factor secretion. Combining it with PD-1 immunotherapy further enhances the antitumor immune response (Lu et al., 2024); The nanoimmunoagonist pLCGM-OVA links cuproptosis with the cGAS-STING pathway to stimulate dendritic cell maturation and strengthen cytotoxic T lymphocyte activity against tumors (Li et al., 2025). Additionally, the ternary heterojunction (HACT) generates reactive oxygen species (ROS) and releases copper ions through sonodynamic therapy, inducing oxidative stress and cuproptosis while exhibiting high tumor-targeting specificity (Huang et al., 2024); drug-loaded nucleic acid nanomedicine (SNAMA) effectively inhibits tumor growth in primary and metastatic uveal melanoma models via GSH release and disulfidptosis activation (Tian et al., 2025).
Key genes associated with cuproptosis and disulfidptosis also exhibit clinical significance. Among cuproptosis-related genes (CRGs), LIPT1 is identified as an independent prognostic factor, positively correlating with PD-L1 expression while negatively regulating Treg cell infiltration (Lv et al., 2022); PPIC is a promoter of melanoma progression, enhancing cell invasiveness while suppressing CD8^+^T cell activation (Zhou et al., 2024). YAP1, a core gene in the Hippo pathway, positively correlates with FDX1 expression in the A2058 cell line, and impacts prognosis through modulation of M2 macrophage and Treg infiltration (Lv et al., 2024). Prognostic models constructed based on cuproptosis-related genes (CRGs) and disulfidptosis-related genes (DRGs) (e.g., CRSS score, lncRNA signature, 2-DRL prognostic model, etc.) enhance prediction accuracy, immunotherapy benefit rate, and tumor microenvironment (TME) status assessment capability (Liu D. et al., 2023; Cheng et al., 2024; Li et al., 2024; Lei et al., 2024; Zhao et al., 2023; Zhou et al., 2022; Yang et al., 2022; Zhu et al., 2024). These findings confirm that CRGs and DRGs have the potential to alter the TME and enhance treatment responsiveness, offering critical strategies for personalized precision medicine (Hu et al., 2023; Chen et al., 2022; Sun et al., 2023; Huang et al., 2023; Yang et al., 2024).
7 Conclusion and perspectives
Ferroptosis, cuproptosis, and disulfidptosis represent three key forms of metabolic cell death that offer novel therapeutic strategies for melanoma. This review highlights their dynamic metabolic competition network in melanoma cells, interconnected through mitochondrial metabolism and metal ion homeostasis regulation. For example, GPX4 depletion not only induces ferroptosis but also functions as an intracellular copper chelator, influencing cuproptosis initiation. Overexpression of SLC7A11 creates a paradoxical balance between ferroptosis resistance and disulfidptosis risk. Also, all three pathways profoundly affect the tumor microenvironment (TME). On the other hand, melanoma resistance mechanisms are closely linked to metabolic reprogramming, including the protection of the lipid microenvironment and the regulation of ACAT2/SOAT, SLC7A11, APOE, and other related genes. To address treatment resistance, the review integrates a variety of innovative anti-melanoma strategies that rely on ferroptosis, cuproptosis, and disulfidptosis, including nanoplatforms such as ES@CuO, TPL@TFBF, and ACM@MCHS-CuMOF@Dox. This review also identifies prognostic models with strong clinical predictive value, such as the CRSS score, lncRNA signature, and 2-DRL prognostic model. These findings suggest that precise regulation of metabolic cell death, redox balance, and interactions with immune checkpoints may overcome melanoma heterogeneity and drug-resistance bottlenecks.
Although these cell death mechanisms share common features, being induced by imbalances in intracellular metal ions or metabolic intermediates, and highlighting the complex interplay between cellular metabolism and death regulation, their research for melanoma therapy still faces significant limitations. Ferroptosis, being the earliest discovered among them, has relatively well-established detection methods, including lipid peroxidation probes, iron level assays, and glutathione metabolism markers. In contrast, detection methods for the more recently identified cuproptosis and disulfidptosis remain preliminary. Current approaches rely on copper ion measurement, FDX1/LIAS protein detection, F-actin morphology observation, SLC7A11 expression, and NADP+/NADPH ratio analysis. The lack of highly reliable and specific biomarkers severely limits accurate assessment and clinical application under pathological conditions, posing a major bottleneck for translational research.
Most existing studies are based on retrospective clinical data, and there is a lack of prospective experiments about melanoma cell sensitivity to different death, particularly under clinically relevant conditions such as untreated and treated (including resistant) conditions. Although metabolic cell death inducers such as copper ionophores (e.g., Elesclomol) and disulfiram have been studied in other cancers and non-cancer (e.g., NCT06635252, NCT05210374), confirming certain translational potential, no active clinical trials currently focus on melanoma patients.
Although targeting metabolic cell death has made some progress in melanoma, clinical translation still faces multiple challenges. Most inducers suffer from poor stability, low solubility, and limited bioavailability, which greatly restrict their clinical use. Developing novel inducers with better pharmacokinetic properties or repurposing FDA-approved anticancer drugs with cell death-inducing activity may partially solve these limitations. Nanomaterials, with their tunability, biocompatibility, and targeting capabilities, offer potential breakthroughs in melanoma treatment, though their actual efficacy and safety still require to be systematically verified. Furthermore, since different cell types within the tumor microenvironment exhibit different sensitivities to these death pathways, non-specific inducers or inhibitors may interfere with other cells, leading to experimental bias. Thus, the absence of cell-specific delivery strategies remains a major limitation.
Future research should focus on solving these critical gaps: first, deepening the mechanistic understanding of cuproptosis and disulfidptosis to clarify their molecular pathways and regulatory networks; second, developing highly specific and sensitive biomarker detection systems to support clinical diagnosis and treatment monitoring; and finally, innovating drug delivery strategies to improve targeting and safety. High-throughput functional screening and artificial intelligence approaches may accelerate the discovery of novel compounds targeting these three death modalities. Subsequent pharmacological studies should confirm their targeted delivery capabilities to enhance safety and efficacy. Targeting metabolic cell death holds promise as a new therapeutic approach in melanoma therapy, especially for patients who do not respond to conventional treatments. Ultimately, these efforts will improve our understanding of the mechanisms of cell death in melanoma and promote the clinical translation of related therapeutic strategies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alekhina O.Burstein E.Billadeau D. D. (2017). Cellular functions of WASP family proteins at a glance. J. Cell Sci. 130 (14), 2235–2241. 10.1242/jcs.199570 28646090 PMC 5536917 · doi ↗ · pubmed ↗
- 2Anestopoulos I.Kyriakou S.Tragkola V.Paraskevaidis I.Tzika E.Mitsiogianni M. (2022). Targeting the epigenome in malignant melanoma: facts, challenges and therapeutic promises. Pharmacol. Ther. 240, 108301. 10.1016/j.pharmthera.2022.108301 36283453 · doi ↗ · pubmed ↗
- 3Apostolova P.Pearce E. L. (2022). Lactic acid and lactate: revisiting the physiological roles in the tumor microenvironment. Trends Immunol. 43 (12), 969–977. 10.1016/j.it.2022.10.005 36319537 PMC 10905416 · doi ↗ · pubmed ↗
- 4Banci L.Bertini I.Ciofi-Baffoni S.Hadjiloi T.Martinelli M.Palumaa P. (2008). Mitochondrial copper(I) transfer from Cox 17 to Sco 1 is coupled to electron transfer. Proc. Natl. Acad. Sci. U. S. A. 105 (19), 6803–6808. 10.1073/pnas.0800019105 18458339 PMC 2383975 · doi ↗ · pubmed ↗
- 5Bayır H.Dixon S. J.Tyurina Y. Y.Kellum J. A.Kagan V. E. (2023). Ferroptotic mechanisms and therapeutic targeting of iron metabolism and lipid peroxidation in the kidney. Nat. Rev. Nephrol. 19 (5), 315–336. 10.1038/s 41581-023-00689-x 36922653 · doi ↗ · pubmed ↗
- 6Bedard K.Krause K. H. (2007). The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol. Rev. 87 (1), 245–313. 10.1152/physrev.00044.2005 17237347 · doi ↗ · pubmed ↗
- 7Bersuker K.Peterson C. W. H.To M.Sahl S. J.Savikhin V.Grossman E. A. (2018). A proximity labeling strategy provides insights into the composition and dynamics of lipid droplet proteomes. Dev. cell 44 (1), 97–112.e 7. 10.1016/j.devcel.2017.11.020 29275994 PMC 5764092 · doi ↗ · pubmed ↗
- 8Bersuker K.Hendricks J. M.Li Z.Magtanong L.Ford B.Tang P. H. (2019). The Co Q oxidoreductase FSP 1 acts parallel to GPX 4 to inhibit ferroptosis. Nature 575 (7784), 688–692. 10.1038/s 41586-019-1705-2 31634900 PMC 6883167 · doi ↗ · pubmed ↗