Isolation, Identification, and Antimicrobial Susceptibility of Exiguobacterium mexicanum from a Giraffe
Fei Gao, Qunchao Liang, Rui Zong, Yuqing Xie, Chenxu Zhao, Yurong Yang, Linyang Yu, Dongliang Li, Hong Duan, Wenjuan Du, Yongtao Li

TL;DR
Scientists isolated a new strain of Exiguobacterium mexicanum from a dead giraffe and tested its response to various antibiotics.
Contribution
First report of E. mexicanum isolated from a giraffe, with detailed antimicrobial susceptibility data for this species in this host.
Findings
HN-1 strain of E. mexicanum was isolated from multiple organs of a deceased giraffe.
The strain showed susceptibility to several antibiotics but resistance to oxacillin, penicillin, ciprofloxacin, and linezolid.
Abstract
Exiguobacterium mexicanum (E. mexicanum) is a facultative anaerobic, Gram-positive bacterium within the genus Exiguobacterium. Although it is commonly found in diverse environments—including soil, water, and extreme habitats such as hot springs and permafrost—this species is rarely linked to infections in humans or animals. This study reports the isolation and characterization of a potentially pathogenic strain of E. mexicanum from a giraffe, along with its antimicrobial susceptibility profile, to inform clinical treatment strategies. The pathogenicity of Exiguobacterium species in wildlife remains poorly understood, with very few documented infections and limited data on host range, virulence factors, and resistance patterns. This gap underscores the significance of the current isolate, suggesting that E. mexicanum may represent an emerging pathogen in giraffes. Notably, despite the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
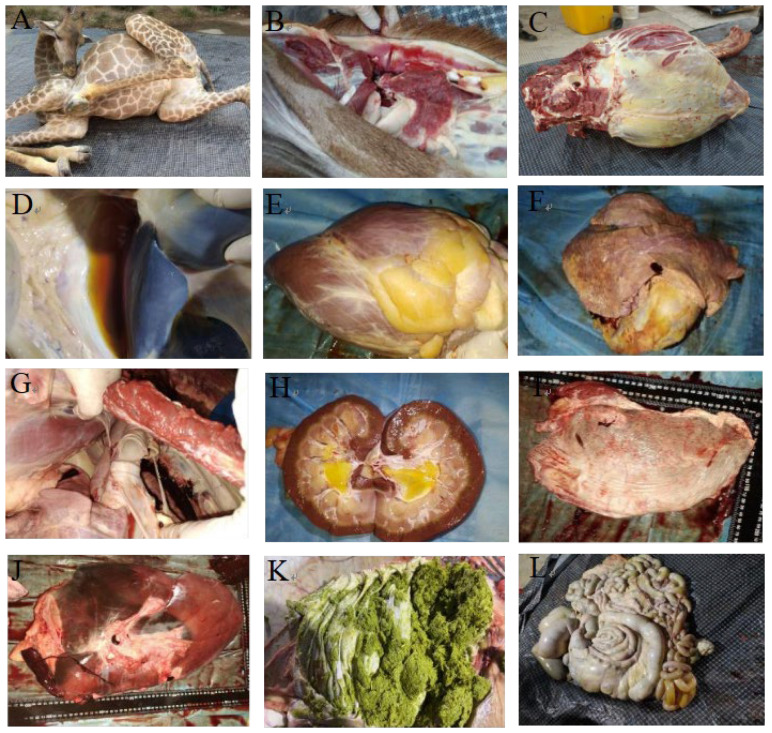 Figure 1
Figure 1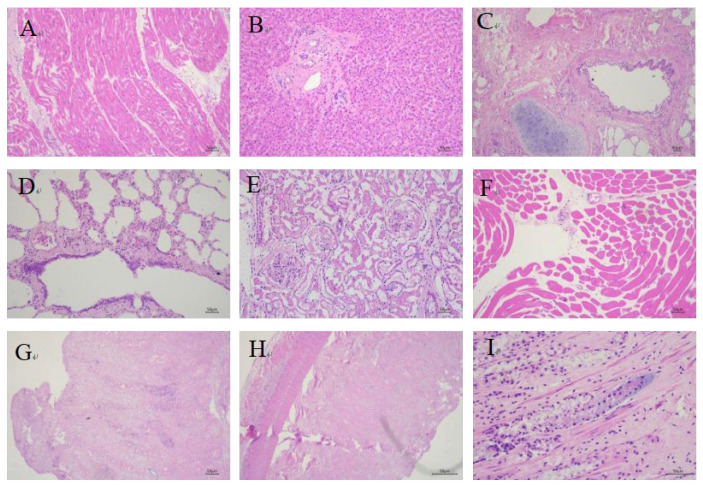 Figure 2
Figure 2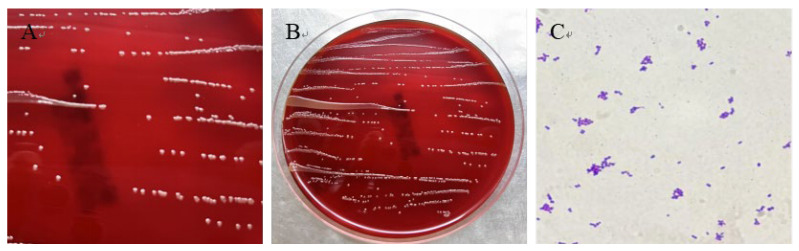 Figure 3
Figure 3 Figure 4
Figure 4- —National Natural Science Foundation of China (NSFC)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEssential Oils and Antimicrobial Activity · Probiotics and Fermented Foods · Aquaculture disease management and microbiota
1. Introduction
Giraffes (Giraffa camelopardalis), the world’s tallest terrestrial animals and largest ruminants [1], face a variety of health challenges in both wild and captive environments stemming from variations in diet [2], environmental conditions, climate [3], and susceptibility to infectious diseases. Captive giraffes are particularly vulnerable to bacterial infections [4,5,6], as exemplified by cases of Mycobacterium avium subsp. paratuberculosis identified in wild ruminants at a Mexican zoo [7], Fusobacterium necrophorum causing severe bilateral necrotic lesions near the tongue base and oral commissures in giraffes at Ordos Zoo [8], and Mycobacterium bovis associated with tuberculosis in giraffes at Greater Kruger National Park [9]. Given the dramatic decline in giraffe populations in recent decades, effective conservation strategies are crucial for ensuring their long-term survival [10].
E. mexicanum is a metabolically versatile bacterium exhibiting noteworthy biological characteristics and potential applications [11,12]. As a member of the Exiguobacterium genus, known for its ability to thrive across broad temperature and pH ranges [9], E. mexicanum was initially isolated and identified as one of two Gram-positive strains from brine shrimp (Artemia) cysts [13]. Subsequent isolations include strain A-EM from Atlantic deep-sea hydrothermal vents [14] and strain HA2 from Ilam Mountain in Iran, which demonstrates growth at 0–25 °C and tolerance up to 5% NaCl [15]. The genus has also been identified in the hyperarid Atacama Desert of Chile [16] and petroleum-contaminated permafrost [17], highlighting E. mexicanum’s adaptability to extreme environmental conditions. While direct evidence of E. mexicanum pathogenicity in animals or humans remains limited, other Exiguobacterium species are recognized as opportunistic pathogens with the potential to infect immunocompromised individuals [18]. Reported cases have linked the genus to community-acquired pneumonia and bacteremia, including a 2017 report of Exiguobacterium sp. isolated from both the blood and bronchoalveolar lavage fluid of a pneumonia patient [11]. Although the specific pathogenic potential of E. mexicanum requires further elucidation, the established pathogenicity of related species warrants continued investigation into its potential health implications.
2. Materials and Methods
2.1. Case Reports and Pathological Examination
In May 2025, a 605 kg female giraffe (ID: 41) in poor body condition died unexpectedly at a zoo in Zhengzhou, Henan Province, China. On the evening of 18 May (after 19:00), the giraffe showed reduced standing time, frequent recumbency, obvious weakness during ambulation, and an increased respiratory rate. It retained its appetite, consuming the provided foliage, but refused to drink water. On 19 May, the giraffe attempted to stand twice, each attempt lasting approximately two minutes, presenting with an unsteady gait, deep and rapid respiration, lethargy, dull eyes, and excretion of dark brown, tea-colored urine. At 11:35, it exhibited limb paddling and neck weakness, and unfortunately died despite emergency treatment. A comprehensive necropsy was subsequently performed, including a thorough pathological examination of all organs. Fresh tissue samples were sent to the Veterinary Diagnostic Laboratory of Henan Agricultural University for further analysis. Samples were collected from visceral organs (heart, liver, spleen, lungs, and kidneys) using sterile scalpels, scissors, and inoculating loops for subsequent histopathological and laboratory studies.
2.2. Bacterial Isolation and Morphological Observation
Under sterile conditions in a biosafety cabinet, samples from the heart, liver, spleen, and kidney were inoculated onto LB agar and blood agar plates (Biocell, Zhengzhou, China) using a sterile inoculating loop. The plates were then incubated at 37 °C for 16–24 h to facilitate colony observation. Single colonies exhibiting uniform morphology were selected and repeatedly streaked to ensure pure cultures displaying consistent colony size and morphology. These purified single colonies were subsequently inoculated into LB liquid medium for culture expansion and preserved on agar plates at 4 °C. Gram staining was performed on the purified bacterial strains, followed by microscopic examination to confirm cell morphology and Gram reaction.
2.3. Molecular Identification
Genomic DNA was extracted from pure bacterial colonies using a DNA purification kit (Tiangen Biotech, Beijing, China), with the extracted DNA stored at −20 °C until use as PCR templates. Molecular identification was performed through sequencing and phylogenetic analysis of the 16S rRNA and recA genes. The 16SrRNA gene was amplified using universal primers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-GGCTACCTTGTTACGACTT-3′) [19]. The recA gene was amplified with primers recA-F (TATCGACTTCTGCCGTCTTGAAC) and recA-R(GTCATCGAAGTGTACGGACCTG).
PCR reactions were performed in a 25 µL volume, comprising 12.5 µL Premix TaqTM, 1 µL each of the forward and reverse primers, 2 µL of DNA template, and 8.5 µL of sterile ddH_2_O. The amplification protocol consisted of an initial denaturation step at 95 °C for 5 min, followed by 30 cycles of denaturation at 95 °C for 30 s, annealing at 59 °C for 30 s, and extension at 72 °C for 20 s, with a final extension at 72 °C for 10 min. PCR products were verified by agarose gel electrophoresis and subsequently submitted to Shangya Biotechnology Co., Ltd. (Henan, China) for sequencing. The resulting sequences were compared to sequences in GenBank using the BLAST algorithm (National Center for Biotechnology Information, NCBI, https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 20 August 2025). Phylogenetic trees, based on both 16S rRNA and recA gene sequences, were constructed using the neighbor-joining method implemented in MEGA 7.0 software, with bootstrap analysis performed using 1000 replicates.
2.4. Antibiotic Susceptibility Testing
A bacterial suspension adjusted to a 0.5 McFarland standard (100 μL) was evenly spread over Mueller-Hinton agar plates (HOPEBIO, Qingdao, China) using a sterile spreader. Antimicrobial susceptibility test disks (OXOID) were placed on the agar surface with sterile forceps, with three replicate plates per disk type. After allowing the plates to stand for 3 min, they were inverted and incubated at 37 °C for 24 h. The diameter of each inhibition zone was measured using a vernier caliper, and results were expressed as the mean value. Since the Clinical and Laboratory Standards Institute (CLSI) has not established interpretive criteria for Exiguobacterium spp., the CLSI criteria for staphylococci were adopted for this study, with Staphylococcus aureus ATCC 25923 as the reference strain, in accordance with CLSI document M100 [20]. This approach was chosen because Exiguobacterium and staphylococci are both Gram-positive bacteria, permitting the use of similar AST methodologies, and staphylococcal criteria are well-validated and widely accepted in clinical and research settings. However, it should be noted that Exiguobacterium and Staphylococcus are phylogenetically distant, and intrinsic differences in membrane permeability, efflux pumps, and natural resistance mechanisms may limit the direct applicability of staphylococcal breakpoints.
3. Results
3.1. Clinical and Pathological Findings in the Deceased Giraffe
Clinically, the affected giraffe exhibited marked lethargy and emaciation. Postmortem examination revealed multi-organ pathological changes: hemorrhage in the cervical lymph nodes (Figure 1B); gelatinous subcutaneous edema in the inguinal region and axilla of the right forelimb; and yellow discoloration of the sternal subcutaneous musculature (Figure 1C). Additionally, the ribs were fragile, with a small amount of brownish-yellow effusion in the peritoneal cavity (Figure 1D). The heart (6.5 kg) showed pale myocardium, yellow gelatinous deposits on the epicardium, scattered petechiae, and extensive thrombi in the cardiac chambers (Figure 1E). The lungs (6.5 kg) presented a marbled appearance (Figure 1F), with widened interlobular septa, localized congestion, pale margins, white tracheobronchial froth, and enlarged hilar lymph nodes (Figure 1G). The kidneys (left: 0.8 kg; right: 1.1 kg) displayed uneven coloration, with yellow gelatinous material in the renal pelvis (Figure 1H). The spleen (0.9 kg) was notably pale (Figure 1I), while the liver (6.15 kg) showed mottled discoloration, accompanied by congestion, firm texture, and generalized atrophy (Figure 1J). Other lesions included varying degrees of enlargement of pancreatic lymph nodes; gastric tympany with mucosal sloughing and greenish frothy contents (Figure 1K); abomasal hemorrhage with rice-sized ulcers; orange-yellow fluid in the intestinal lumen (Figure 1L); petechiae in the cecum and colon; and mucoid rectal feces. Collectively, these pathological findings are consistent with a diagnosis of acute bacterial septicemia complicated by multi-organ failure
3.2. Histopathological Findings in the Deceased Giraffe
Histopathological examination of the deceased giraffe revealed significant pathological changes, characterized by multi-organ atrophy, extensive necrosis, and inflammatory cell infiltration in multiple tissues. Myocardial fibers showed dissolution and disorganization, with partial cardiomyocyte degeneration and necrosis manifested as pyknosis or loss of nuclei (Figure 2A). The liver exhibited atrophied hepatocyte cords with minimal bile pigment deposition, collagen fiber proliferation in the portal triads, and sparse lymphocytic infiltration (Figure 2B). Bronchial mucosal epithelium was desquamated with loss of cilia, accompanied by the peribronchial accumulation of lymphocytes and plasma cells (Figure 2C). Scattered erythrocytes, neutrophils, desquamated epithelial cells, and exudates were observed in pulmonary vessels, along with perivascular fibrin deposition, alveolar macrophages, dust cells, and squamous metaplasia of bronchiolar mucosa (Figure 2D). Renal lesions included degenerative and necrotic tubular epithelial cells with fibrinous casts in the lumens, atrophied glomeruli with capsular degeneration and exudate accumulation, and interstitial inflammatory cell infiltration (Figure 2E). Skeletal muscles displayed atrophy and structural disorganization (Figure 2F). Transmural necrosis was present in the gastric wall, with abundant rod-shaped bacteria observed in the mucosal glands (Figure 2G). Intestinal tissues showed transmural necrosis with disintegration of mucosal epithelium and glandular architecture, blurred cellular boundaries, and nuclear loss (Figure 2H), along with dense aggregates of rod-shaped bacteria in the intestinal glands accompanied by peripheral tissue necrosis (Figure 2I). These histopathological changes indicate severe pyogranulomatous inflammation and extensive tissue necrosis, suggesting that the pathogen is likely an invasive pathogenic bacterium capable of inducing a fatal septic shock-like syndrome.
3.3. Bacterium Isolation and Microscopic Examination
Sterilely collected tissue samples were inoculated onto blood agar plate (Biocell, Zhengzhou, China) and LB agar plates for bacterial isolation and culture. A total of 18 bacterial isolates were isolated from heart, spleen, lung, kidney, and liver tissues. One strain, found across multiple tissues, was designated as HN-1. HN-1 colonies were observed to be round, smooth, pale yellow, and raised in morphology (Figure 3A,B). Gram staining identified the newly isolated HN-1 strain as Gram-Positive, displaying a short, rod-shaped morphology with blunt ends, and arranged singly or in pairs (Figure 3C). These characteristics, indicative of systemic infection, suggest that strain HN-1 possesses biological properties consistent with septicemia pathogens.
3.4. Strain HN-1 Was Identified as E. mexicanum
HN-1 strain underwent further characterization through 16S rRNA and recA gene sequence analysis. Amplification yielded 16S rRNA and recA gene fragments of 1446 bp (GenBank accession number: PX248038) and 575 bp (GenBank accession number: PX255259), respectively. The agarose gel electrophoresis of the PCR products (Figure 4A) shows three lanes for both 16S rRNA and recA gene amplicons, corresponding to bacterial colonies isolated from three independent culture plates, confirming the consistency of the amplification results across different isolates of the same strain. Subsequent BLAST analysis against the NCBI database revealed high sequence similarity between strain HN-1 and various Exiguobacterium strains. Specifically, the 16S rRNA sequence exhibited 99.73–99.86% similarity, while the recA gene sequence showed 98.26–100% similarity. Furthermore, mass spectrometry confirmed that the bacterial strains isolated from different samples were all identified as E. mexicanum. Phylogenetic trees, constructed using both 16S rRNA and recA gene sequences from HN-1 and relevant reference strains, provided further insight into its taxonomic position. The 16S rRNA phylogenetic tree (Figure 4B) demonstrated that HN-1 clustered exclusively with E. mexicanum OK135833. Likewise, the recA phylogenetic tree (Figure 4C) indicated that strain HN-1 formed a distinct clade encompassing all Exiguobacterium species included in the analysis. Integrating morphological characteristics with the results of 16S rRNA and recA gene sequence analyses, as well as confirmation by mass spectrometry, the bacterial strain HN-1 was conclusively identified as E. mexicanum.
3.5. Antimicrobial Susceptibility Detection
To investigate the antimicrobial susceptibility profile of E. mexicanum HN-1, the disk diffusion method was used to determine the susceptibility of this strain to 14 antibiotics. The results showed that strain HN-1 exhibited resistance to oxacillin, penicillin, ciprofloxacin, and linezolid, which was characterized by small inhibition zones. In contrast, the strain was susceptible to levofloxacin, clindamycin, chloramphenicol, trimethoprim, rifampicin, tetracycline, minocycline, gentamicin, erythromycin, and doxycycline (Table 1). These findings not only clarify the resistance characteristics of this strain to certain antibiotics but also provide potential therapeutic options for clinical infections caused by Exiguobacterium mexicanum HN-1.
4. Discussion
Global outbreaks of zoonotic diseases have become increasingly frequent in recent decades, posing escalating threats to public health and wildlife conservation [21]. Advances in diagnostic technologies have facilitated the identification of emerging zoonoses—infections caused by bacteria, parasites, viruses, and fungi that originate in animals and are transmitted to humans from wild or domestic reservoirs [18,22]. Within this context, the genus Exiguobacterium, a group of Gram-positive bacteria renowned for their exceptional adaptability (including thermotolerance, psychrotolerance, alkaliphily, and halotolerance) [23,24], has garnered increasing attention due to its underrecognized pathogenic potential. Although reports of Exiguobacterium-related infections remain scarce, existing evidence underscores their clinical significance. For instance, E. aurantiacum has been isolated from blood cultures of bacteremia patients, including those with multiple myeloma, highlighting the heightened susceptibility of immunocompromised individuals [25]. In China, Exiguobacterium sp. A1b/GX59 has been associated with community-acquired pneumonia and bacteremia, with the pathogen detected in both blood and bronchoalveolar lavage fluid [11]. These cases align with our findings but also reveal a critical knowledge gap: prior to this study, no cases of E. mexicanum infection had been reported in giraffes, nor had systemic infection leading to fatal septicemia been documented. This novelty establishes our work as a pivotal step in expanding the known host range of E. mexicanum—previously limited to environmental niches and human clinical cases—to include large ruminants, particularly captive wildlife.
E. mexicanum, an opportunistic pathogen ubiquitous in soil and water [26,27], likely infects giraffes through multiple transmission routes such as ingestion of contaminated feed or water, skin wounds, or respiratory exposure [28]. This is consistent with the systemic distribution of strain HN-1 across the heart, liver, spleen, lungs, and kidneys observed in this case. Captive giraffes, already vulnerable to stress-induced immune impairment due to overcrowding or poor sanitation [29], may face elevated risks, as evidenced by the severe pathological lesions—including multi-organ necrosis, thrombus formation, and inflammatory infiltration—identified in our study. Notably, this contrasts with prior research on E. mexicanum, which has focused primarily on its role in environmental bioremediation (e.g., nitrogen removal in saline wastewater) [30,31,32] rather than its pathogenicity, underscoring the need to reevaluate its ecological and clinical importance. Currently, antibiotic therapy remains the primary treatment option due to its broad applicability and rapid efficacy [33]. In terms of antimicrobial susceptibility, our findings reveal a distinct profile compared to related species. While six E. aurantiacum strains isolated from human blood cultures showed susceptibility to all the tested antibiotics [34], whereas E. mexicanum HN-1 showed resistance to oxacillin, penicillin, ciprofloxacin, and linezolid, and susceptibility to levofloxacin, clindamycin, chloramphenicol, trimethoprim, rifampicin, tetracycline, minocycline, gentamicin, erythromycin, and doxycycline. This resistance pattern may reflect local antibiotic usage practices [35] and also emphasizes the importance of conducting species-specific antimicrobial susceptibility testing to avoid ineffective treatments. For the management of captive wildlife, these data provide actionable guidance: prioritizing the use of susceptible antibiotics such as levofloxacin, clindamycin, and gentamicin, while avoiding resistant drugs like penicillin and oxacillin, may improve the cure rate of future cases.
Collectively, this study addresses key gaps in our understanding of E. mexicanum pathogenicity. By documenting the first case of E. mexicanum infection in giraffes, we confirm that this bacterium can infect large herbivores, thereby expanding its known host range. The observed antibiotic resistance further highlights the need for targeted surveillance in captive settings, where the risk of zoonotic spillover may be amplified. Moving forward, investigations into the bacterium’s virulence factors (e.g., adhesion proteins or toxin production) and the links between environmental reservoirs and animal or human infections will be critical. For zoos and wildlife facilities, our work underscores the urgency of enhancing biosecurity measures—including pathogen monitoring, stress reduction, and prudent antibiotic use—to mitigate risks posed by this emerging pathogen, ultimately safeguarding both animal welfare and public health [36,37,38].
5. Conclusions
This study reports on the first case in which E. mexicanum was isolated and identified, from a deceased giraffe, with strain HN-1 exhibiting resistance to penicillin, oxacillin, ciprofloxacin, and linezolid while being susceptible to most other tested antibiotics. The systemic distribution of HN-1 in multiple visceral organs and the severe pathological changes observed suggest its potential role in the giraffe’s fatal septicemia. However, it is important to acknowledge that 18 bacterial strains were isolated from the animal, and only HN-1 was characterized. Thus, we cannot rule out the possibility of a mixed infection, where other unstudied strains may have contributed to the pathogenesis. Future research should focus on the comprehensive identification of all isolates, assessment of their pathogenicity in combination, and exploration of virulence factors to clarify the exact etiology of such infections in giraffes. These findings provide a foundation for understanding E. mexicanum infections in large ruminants and offer guidance for clinical treatment in captive wildlife settings.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bashaw M.J. Tarou L.R. Maki T.S. Maple T.L. A survey assessment of variables related to stereotypy in captive giraffe and okapi Appl. Anim. Behav. Sci.20017323524710.1016/S 0168-1591(01)00137-X 11376840 · doi ↗ · pubmed ↗
- 2Kearney C.C. Ball R.L. Hall M.B. Effects of altering diet carbohydrate profile and physical form on zoo-housed giraffe Giraffa camelopardalis reticulata J. Anim. Physiol. Anim. Nutr.20241081119113310.1111/jpn.1395738590078 · doi ↗ · pubmed ↗
- 3Saito M. Matsunaga M. Fukuizumi H. Nakamichi M. Kinoshita K. Factors affecting captive female giraffe stress response: Male presence, small enclosure, and low temperature Zoo Biol.20234263264310.1002/zoo.2177437154219 · doi ↗ · pubmed ↗
- 4Passos L.F. Garcia G. Young R.J. Comparing the bacterial communities of wild and captive golden mantella frogs: Implications for amphibian conservation P Lo S ONE 201813 e 020565210.1371/journal.pone.020565230379861 PMC 6209184 · doi ↗ · pubmed ↗
- 5Diaz J. Reese A.T. Possibilities and limits for using the gut microbiome to improve captive animal health Anim. Microbiome 202138910.1186/s 42523-021-00155-834965885 PMC 8715647 · doi ↗ · pubmed ↗
- 6Schmidt D.A. Barbiers R.B. Ellersieck M.R. Ball R.L. Koutsos E.A. Griffin M.E. Grobler D. Citino S.B. Bush M. Serum Chemistry Comparisons Between Captive and Free-ranging Giraffes (Giraffa camelopardalis)J. Zoo Wildl. Med.201142333910.1638/2009-0218.122946367 · doi ↗ · pubmed ↗
- 7Hernández-Reyes A.L. Chávez-Gris G. Maldonado-Castro E. Alcaraz-Sosa L.E. Díaz-Negrete M.T. First identification of Mycobacterium avium subsp. paratuberculosisin wild ruminants in a zoo in Mexico Vet. World 20221565566110.14202/vetworld.2022.655-66135497960 PMC 9047120 · doi ↗ · pubmed ↗
- 8Wang J. Wang Y. Ding Y. Suljid J. Wang W. Oral and pulmonary necrobacillosis in a juvenile reticulated giraffe J. Vet. Diagn. Investig.20213334534710.1177/104063872098781833446090 PMC 7953106 · doi ↗ · pubmed ↗