Antimicrobial Susceptibility of Campylobacter spp. Isolated from Cattle in Mongolia
Erdenebat Bulgan, Zolzaya Byambajav, Batsukh Naranchimeg, Batsaikhan Chantsal, Tsognemekh Bolormaa, Badrakh Sandagdorj, Purevdorj Nyam-Osor, Eisaku Kikuchi, Akio Suzuki, Jirachaya Toyting-Hiraishi, Toyotaka Sato, Motohiro Horiuchi

TL;DR
This study reports the first characterization of Campylobacter species in cattle in Mongolia, finding diverse strains with distinct antibiotic resistance patterns compared to chicken isolates.
Contribution
The first characterization of Campylobacter spp. in cattle in Mongolia, revealing unique antimicrobial resistance patterns and genetic diversity.
Findings
Thirty-five Campylobacter isolates were identified, including 23 C. jejuni, with substantial genetic diversity observed.
Cattle C. jejuni isolates showed antimicrobial resistance patterns distinct from chicken isolates, with most being susceptible to tetracycline and ciprofloxacin.
Sequence type ST61, associated with cattle and sheep, was identified among the isolates.
Abstract
This study aimed to assess prevalence and antimicrobial resistance of Campylobacter spp. in cattle in Mongolia. Thirty-five Campylobacter spp., including 23 C. jejuni, 7 C. hyointestinalis, 4 C. fetus, and 1 C. lari, were isolated. Multilocus sequence typing of C. jejuni cattle isolates revealed substantial genetic diversity. The antimicrobial resistance patterns of the C. jejuni cattle isolates completely differed from those of previously reported chicken isolates; excluding one ciprofloxacin-resistant isolate, all C. jejuni isolates were susceptible to tetracycline and ciprofloxacin. To the best of our knowledge, this is the first report on the characterization of Campylobacter spp. in cattle in Mongolia. Poultry and cattle are the major reservoirs of Campylobacter infection in humans. However, no information is available on Campylobacter spp. in cattle in Mongolia. Thus, this study…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
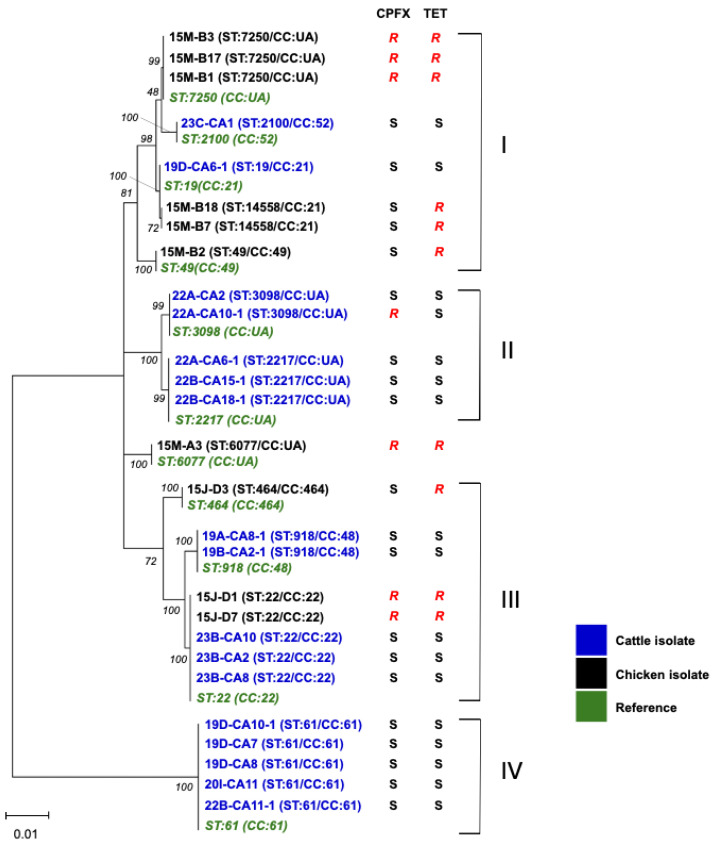 Figure 1
Figure 1- —International Collaborative Research
- —Ministry of Education, Culture, Sports, Science, and Technology, Japan
- —JICA Agriculture Studies Networks for Food Security (Agri-Net)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSalmonella and Campylobacter epidemiology · Veterinary medicine and infectious diseases · Viral gastroenteritis research and epidemiology
1. Introduction
Campylobacter infection is a leading cause of foodborne bacterial diseases worldwide. Campylobacter spp. colonize in the intestines of homoiothermal animals asymptomatically in most cases except for C. fetus infection in cattle, which causes bovine genital campylobacteriosis. Among the Campylobacter spp., C. jejuni and C. coli are the predominant causes of campylobacteriosis in humans. Additionally, species such as C. fetus, C. lari, and C. hyointestinalis are also known to be pathogenic to humans [1,2]. The primary clinical symptoms of Campylobacter infection are diarrhea, abdominal pain, fever, headache, nausea, and vomiting. Although most cases are self-limited within 3–6 days without any treatment, macrolides and fluoroquinolones (FQs) are prescribed to patients with severe or prolonged symptoms. Campylobacter infection is recognized as a risk factor for Guillain-Barré syndrome (GBS), an autoimmune demyelinating polyneuropathy [3]. Nearly 30% of GBS cases are associated with antecedent infection of Campylobacter [4,5], and about 0.1% of infected individuals develop GBS [6].
Livestock such as poultry and ruminants are major attributed sources of Campylobacter infection in humans, and companion animals and environments including surface water are also sources of infection [7,8]. As the route of infection can vary by region depending on differences in livestock breeding conditions, livestock hygiene management, food hygiene practice, food culture, and the coexisting environment between humans and animals, the identification of the source and infection route to humans is important for implementing effective measures against campylobacteriosis.
In Mongolia, meat is thoroughly cooked before consumption; thus, it is unlikely that undercooked meat poses a risk of infection. However, there are other risks of foodborne campylobacteriosis such as ingestion of traditional dairy products made from unpasteurized animal milk, secondary contamination during processing and along the food chain, the spread of ready-to-eat foods such as salads, the westernization of dietary habits, and the use of surface water in rural areas.
We recently reported the high antimicrobial resistance (AMR) ratios of Campylobacter spp. in chickens in Mongolia. All isolates were resistant to tetracycline (TET), and more than half were resistant to FQs. Half of the C. coli isolates exhibited muti-drug resistance, including resistance to erythromycin (EM) [9]. Because WHO assigned EM and ciprofloxacin (CPFX) as critically important antimicrobials for treating severe campylobacteriosis in humans, the presence of macrolide- and/or FQ-resistant Campylobacter spp. in food-producing animals, foods, and environments represents a public health concern. However, no data on the existence and AMR of Campylobacter spp. in other livestock and contamination of Campylobacter spp. in food chain are available. Additionally, campylobacteriosis in humans has not been officially reported to date, probably because of the insufficient diagnostic capacity.
More than 5 million cattle, 25 million sheep, and 23 million goats are raised in Mongolia. Meat is one of the primary food products in Mongolia, and beef and mutton are widely consumed. The annual consumption is approximately 134,000 tons for beef and 275,000 tons for sheep and goat meat [10]. Additionally, Mongolia has a unique culture of using animal milk and consumes 160,000 tons of milk per year. These facts imply that ruminants are possible sources of Campylobacter infection in humans. Therefore, in the current study, we characterized Campylobacter spp. isolated from cattle to provide scientific data for investigating the zoonotic transmission route of Campylobacter spp. in Mongolia.
2. Materials and Methods
2.1. Sampling
In total, 100 rectal swabs were collected from 10 dairy farms in Ulaanbaatar, Mongolia (Table 1). Most samples were collected from the Songinokhairkhan district, a cattle farm-concentrated area in Ulaanbaatar. Rectal swabs were collected using sterile cotton swabs. Each swab was directly placed in 5 mL of Brucella broth (Becton Dickinson, Franklin Lakes, NJ, USA) supplemented with Modified Preston Campylobacter Supplement (Oxoid, Basingstoke, UK) and 5% lysed horse blood (LHB, Nippon Bio-test Laboratories Inc., Asaka, Japan; the supplemented broth is hereafter termed Preston broth) and kept cooled during transportation to the laboratory.
2.2. Isolation and Identification of Campylobacter spp.
Preston broth containing the rectal swabs was incubated for 24 h at 42 °C under microaerobic conditions using AnaeroPacks-MicroAero (Mitsubishi Gas Chemical Co., Inc., Tokyo, Japan) to enrich thermophilic Campylobacter spp. Then, one loop of the broth was streaked on Campylobacter Blood-Free Selective Agar Base (Oxoid) with Charcoal Cefoperazone Deoxycholate Agar (CCDA) selective supplement (Oxoid; hereafter termed mCCDA), and the mCCDA plates were incubated for 48 h at 37 °C under microaerobic conditions. Typical Campylobacter colonies on the mCCDA plates were picked up using sterilized toothpicks and inoculated into Campylobacter Blood-Free Selective Agar Base without CCDA supplement to analyze bacterial growth under microaerobic and anaerobic conditions using AnaeroPacks-MicroAero and AneroPak-Anaero (Mitsubishi Gas Chemical Co., Inc.), respectively.
The suspected Campylobacter colonies were cultured in 2 mL of Brucella broth for 48 h at 37 °C under microaerobic conditions, and DNA was extracted from the cultures using CICA Geneus DNA Extraction Reagent (Kanto Chemical Co., Inc., Tokyo, Japan) according to the supplier’s instructions. Species-specific multiplex PCR targeting cdtC gene [11], which can distinguish C. jejuni, C. coli, and C. fetus, was performed as described previously [9]. Campylobacter spp. were also confirmed by matrix-assisted laser desorption/ionization (MALDI) time-of-flight mass spectrometry using the MALDI Biotyper Compass (Bruker Daltonics, Bremen, Germany).
2.3. Antimicrobial Susceptibility Test
The antimicrobial susceptibility of the isolates was analyzed using the E-test and broth dilution method. E-test strips of TET, EM, nalidixic acid (NA), and ciprofloxacin (CPFX) were purchased from BioMérieux (Marcy-l’Etoile, France), and E-test was performed as descrived previously [9]. The minimum inhibitory concentrations (MIC) for NA were also determined using the broth dilution method according to the CLSI guidelines. Antimicrobial susceptibility was determined according to CLSI M45 third edition, with resistant breakpoints of EM (≥32 μg/mL), CPFX (≥4 μg/mL), and TET (≥16 μg/mL), and according to Japanese Veterinary Antimicrobial Resistance Monitoring System (JVARM) for NA (≥16 μg/mL).
2.4. Genetic Analysis
Nucleotide sequencing of the gyrA gene to analyze codon 86 amino acid substitution for estimating FQ susceptibility and 16S rDNA for species identification, was performed as described elsewhere [9]. Multi-locus sequence typing (MLST) was performed to identify sequence types (STs) and clonal complexes (CCs) of the C. jejuni isolates. Seven housekeeping genes (aspA, glnA, gltA, glyA, pgm, tkt, and uncA) were amplified by PCR using the primer sets listed in the Campylobacter MLST database [12], and the amplified DNA fragments were purified using a FastGene Gel/PCR Extraction Kit (Nippon Genetics Co., Ltd., Tokyo, Japan). Nucleotide sequences of the amplified fragments were determined using the BigDye Terminator 3.1 Cycle Sequencing Kit and 3130-Avant Genetic Analyzer (Applied Biosystems, Waltham, MA, USA). Phylogenetic analyses were conducted using Molecular Evolutionary Genetics Analysis version 11 [13]. The nucleotide sequences of aspA (409 base pairs [bp], nucleotide [nt] 669–1077), glnA (452 bp, nt 247–698), gltA (398 bp, nt 321–719), glyA (403 bp, nt 392–794), tkt (438 bp, nt 247–684), uncA (482 bp, nt 676–1157), and pgm (498 bp, nt 421–1118) were used for the phylogenetic analyses.
3. Results
3.1. Isolation and Identification of Campylobacter spp.
Between 2019 and 2023, we visited 10 dairy farms (9 farms in Songinokhairkhan district were within a 40-km radius of central Ulaanbaatar, while farm 20I in Baganuur district was 130 km away from central Ulaanbaatar), and collected 10 rectal swabs from cattle in each farm. A total of 35 Campylobacter spp. were isolated (Table 1), including 23 C. jejuni (65.7%) and 4 C. fetus (11.4%) identified by multiplex PCR. The remaining isolates were identified as C. hyointestinalis (n = 7, 20.0%) by 16S rDNA sequencing analysis and MALDI Biotyper analysis and C. lari (n = 1, 2.9%) by MALDI Biotyper analysis. No C. coli was isolated in the current study. All C. jejuni and C. fetus isolates were also confirmed by MALDI Biotyper analysis. Of the 10 farms, C. jejuni was isolated from 9 farms, whereas in farm 19C, only C. hyointestinalis was isolated. In farms 19D, 20I, and 23B, only C. jejuni was isolated, whereas more than one Campylobacter spp. including C. jejuni, C. fetus, C. hyointestinalis, or C. lari were isolated from six farms (19A, 19B, 22A, 22B, 23A, and 23C). Two different Campylobacter spp., C. jejuni and C. hyointestinalis, were isolated from cattle ID CA2 in farm 19B and cattle ID CA11 in farm 22B (Table 2).
3.2. AMR of Campylobacter spp. Isolates
We previously reported that C. jejuni isolates from chickens in Mongolia exhibited extremely high AMR ratios; all the isolates were resistant to TET and half of the isolates were resistant to NA and FQs [9]. Different from the antimicrobial resistance patterns of C. jejuni isolates from chickens, most of the C. jejuni isolates from cattle in Mongolia were sensitive to TET and CPFX; among the 23 C. jejuni isolates, only one isolate (22A-CA10-1) was resistant to CPFX and NA. Nucleotide sequence analysis of gyrA of isolate 22A-CA10-1 confirmed that nucleotide substitution at nt257 (ACA to ATA) resulted in a Thr-to-Ile amino acid substitution at codon 86, which is associated with FQ resistance in C. jejuni [14]. Consistent with the susceptibility to CPFX, other C. jejuni isolates carried Thr at codon 86 (Table 2). C. hyointestinalis, C. fetus, and C. lari are known to be intrinsically resistant to NA [15,16,17], and consistent with this, all three Campylobacter spp. isolated in this study were resistant to NA (Table 2).
3.3. MLST and Phylogenetic Analysis
For MLST, 17 and 10 C. jejuni isolates from cattle (this study) and chickens [9] in Mongolia, respectively, were used. The 17 cattle isolates were classified into 7 STs (ST19, ST22, ST61, ST918, ST2100, ST2217, and ST3098) (Table 2), suggesting the diversity of C. jejuni in cattle in Mongolia. Twelve C. jejuni isolates from cattle with ST19, ST22, ST61, ST918, and ST2100 belonged to CC19, CC21, CC22, CC48, and CC52, respectively. The CCs of the remaining 5 isolates with ST2217 or ST3098 were not identified. The chicken isolates 15M-B7 and 15M-B18 were assigned to ST14448 (CC21), a new ST (Figure 1).
Figure 1 shows phylogenetic relationships of the isolates from cattle and chickens based on the seven genes used for MLST analysis. Clusters I and III comprised isolates from chickens and cattle, whereas clusters II and IV solely contained cattle isolates. Within cluster II, ST3098 and ST2217 have only been reported in cattle or beef, with the exception of a single human blood isolate, suggesting the association of these STs with cattle [12]. Cluster III included ST22 isolates from both chickens and cattle; however, the cattle isolates were susceptible to TET and CPFX but chicken isolates were resistant to these antibiotics. Cluster IV only included ST61, which is known as a cattle/sheep-associated ST worldwide [8]. ST61 C. jejuni were isolated from different farms, suggesting a wide distribution among cattle in Mongolia.
4. Discussion
AMR monitoring of C. jejuni in humans and food-producing animals conducted by member states of the European Unions revealed extremely high ratios of TET and CPFX resistance in three animal species with sufficient isolate numbers including broilers (ranging from 24.1 to 84.6% and 38.0 to 97.3%, respectively), fattening turkeys (18.8–76.3% and 34.1–93.2%, respectively), and cattle under 1-year old (35.3–92.6% and 45.7–79.3%, respectively), across all nations excluding Nordic countries [18]. Increased trends of CPFX-resistant C. jejuni both in human and livestock have been reported in European countries and globally [19,20]. Because EM and CPFX are assigned as critically important antimicrobials for treating severe campylobacteriosis in humans [21], the presence of macrolide- and/or FQ-resistant Campylobacter spp. in food-producing animals, foods, and environments represents a public health concern. We previously reported that C. jejuni isolated from chickens in Mongolia showed extremely high AMR ratios; all isolates were resistant to TET and nearly half were resistant to FQs [9]. However, this trend was not observed in C.jejuni isolates from cattle in this study, as all isolates were susceptible to TET and CPFX, excluding one isolate with CPFX-resistance (22A-CA10-1) carried a Thr-to-Ile substitution at codon 86 in gyrA. This unique difference in TET and FQ resistance between chickens and cattle suggests that the transmission of Campylobacter spp. between the two livestock species is rare in Mongolia. In the questionnaire survey, we could not obtain any evidence of FQ use on chicken and dairy farms. It has been noted that no fitness burden is required for FQ-resistant C. jejuni to colonize the chicken intestine [22,23]. Similarly, it is reported that FQ-resistant C. jejuni might have a fitness advantage over FQ-sensitive isolates in colonizing the cattle intestine [24,25]. Thus, although the current prevalence of FQ-resistant C jejuni is low in cattle, AMR monitoring, including that of Campylobacter spp. in cattle, should be strengthened to detect trends as quickly as possible.
ST61 C. jejuni is known to be associated with cattle and sheep [8] and associated with CPFX sensitivity significantly [26]. Consistent with these findings, ST61 was only found in cattle isolates, all of which were sensitive to CPFX. Although undercooked beef is not consumed often because of the Mongolian food culture, C. jejuni in cattle can spread to humans via cross-contaminated foods during processing and along the food chain, surface water contaminated with cattle feces, and the consumption of raw milk and dairy products. Indeed, Campylobacter spp. are considered as one of the major hazards associated with raw drinking milk [27]. In Japan, where undercooked poultry is the main attribution source of food-borne campylobacteriosis, ST61 C. jejuni caused an outbreak by consumption of raw cow milk [28]. Raw ruminant milk is a potential risk of Campylobacter infection in humans in Mongolia because it is used for a variety of traditional dairy products such as cream and cheese.
One of the limitations in this study is that most of the samples were collected in the Songinokhairkhan district of Ulaanbaatar, a region where many dairy farms reside. Genetic diversity was observed among the C. jejuni isolates from this district; however, continued analysis of a larger number of Campylobacter spp. isolated from different regions will be required to clarify the characteristics of Campylobacter spp. in cattle in Mongolia.
Although there are no official statistics on campylobacteriosis in humans in Mongolia, a genetic fragment of Campylobacter spp. has been detected by PCR in human diarrhea samples of unknown cause (personal communication, Dr. Tundev Odgerel at National Center for Communicable Disease, Mongolia). At present, no information on the attribution source of human campylobacteriosis is available. Future molecular genetic studies of isolates from humans, animals, foods, and environments under the One Health concept will uncover the exposure route of Campylobacter spp. to humans.
5. Conclusions
We recently reported the high antimicrobial resistance ratios of C. jejuni in chickens in Mongolia; all isolates were resistant to TET, and nearly half were resistant to FQs. However, this study revealed that the antimicrobial resistance patterns of the C. jejuni cattle isolates completely differed from those of chicken isolates. This is the first report on the characterization of Campylobacter spp. in cattle in Mongolia. Data on Campylobacter spp. in food-producing animals will be valuable for investigating potential sources and infection routes to humans.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Jansen A. Stark K. Kunkel J. Schreier E. Ignatius R. Liesenfeld O. Werber D. Göbel U.B. Zeitz M. Schneider T. Aetiology of community-acquired, acute gastroenteritis in hospitalised adults: A prospective cohort study BMC Infect. Dis.2008814310.1186/1471-2334-8-14318940017 PMC 2596151 · doi ↗ · pubmed ↗
- 2Kaakoush N.O. Castaño-Rodríguez N. Mitchell H.M. Man S.M. Global Epidemiology of Campylobacter Infection Clin. Microbiol. Rev.20152868772010.1128/CMR.00006-1526062576 PMC 4462680 · doi ↗ · pubmed ↗
- 3Blaser M.J. Engberg J. Clinical Aspects of Campylobacter jejuni and Campylobacter coli Infection 3rd ed. Nachamkin I. Szymanski C.M. Blaser M.J. ASM Press Washington, DC, USA 2008
- 4Poropatich K.O. Walker C.L. Black R.E. Quantifying the association between Campylobacter infection and Guillain-Barré syndrome: A systematic review J. Health Popul. Nutr.20102854555210.3329/jhpn.v 28i 6.660221261199 PMC 2995022 · doi ↗ · pubmed ↗
- 5Rees J.H. Soudain S.E. Gregson N.A. Hughes R.A. Campylobacter jejuni infection and Guillain-Barré syndrome N. Engl. J. Med.19953331374137910.1056/NEJM 1995112333321027477117 · doi ↗ · pubmed ↗
- 6Keithlin J. Sargeant J. Thomas M.K. Fazil A. Systematic review and meta-analysis of the proportion of Campylobacter cases that develop chronic sequelae BMC Public Health 201414120310.1186/1471-2458-14-120325416162 PMC 4391665 · doi ↗ · pubmed ↗
- 7Mughini-Gras L. Pijnacker R. Coipan C. Mulder A.C. Fernandes Veludo A. de Rijk S. van Hoek A. Buij R. Muskens G. Koene M. Sources and transmission routes of campylobacteriosis: A combined analysis of genome and exposure data J. Infect.20218221622610.1016/j.jinf.2020.09.03933275955 · doi ↗ · pubmed ↗
- 8Sheppard S.K. Dallas J.F. Strachan N.J. Mac Rae M. Mc Carthy N.D. Wilson D.J. Gormley F.J. Falush D. Ogden I.D. Maiden M.C. Campylobacter genotyping to determine the source of human infection Clin. Infect. Dis.2009481072107810.1086/59740219275496 PMC 3988352 · doi ↗ · pubmed ↗