Mosquito Feeding Preference, Infectivity Rates, and Knockdown Resistance Within the Wild Population of Anopheles arabiensis in Jabi Tehnan District, Northwest Ethiopia
Alemnesh Hailemariam Bedasso, Sisay Dugassa, Jimma Dinsa Deressa, Geremew Tasew Guma, Getachew Tolera Eticha, Mesay Hailu Dangisso, Eliningaya J. Kweka, Habte Tekie

TL;DR
This study examines the feeding habits, infectivity, and insecticide resistance in Anopheles arabiensis mosquitoes in Ethiopia to guide malaria control strategies.
Contribution
The study provides new insights into the species composition, blood meal preferences, and knockdown resistance allele frequencies of Anopheles mosquitoes in Jabi Tehnan District.
Findings
Anopheles arabiensis was the dominant malaria vector species in the study area.
A significant proportion of An. arabiensis mosquitoes showed knockdown resistance gene mutations.
Most blood meals of An. arabiensis were of human origin.
Abstract
Background: In recent decades, malaria vector species distribution and insecticide resistance have taken new colonization steps across Africa. Understanding the malaria vector insecticide resistance status, blood meal source, and species composition is of paramount importance in designing evidence-based vector control strategies. This study assessed the blood meal sources, sporozoite (infectivity) rate, and knockdown resistance allele’s frequency in female Anopheles arabiensis in chosen villages of Jabi Tehnan District, Northwest Ethiopia. Methods: The host-seeking and resting Anopheles gambiae s.l. were collected using human landing catches (HLCs), CDC light traps (CDC-LTs), pyrethrum spray catches (PSCs), and pit shelters (PSs) both indoors and outdoors. The analysis of both blood meal sources and circumsporozoite proteins was performed using enzyme-linked immunosorbent assay (ELISA).…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
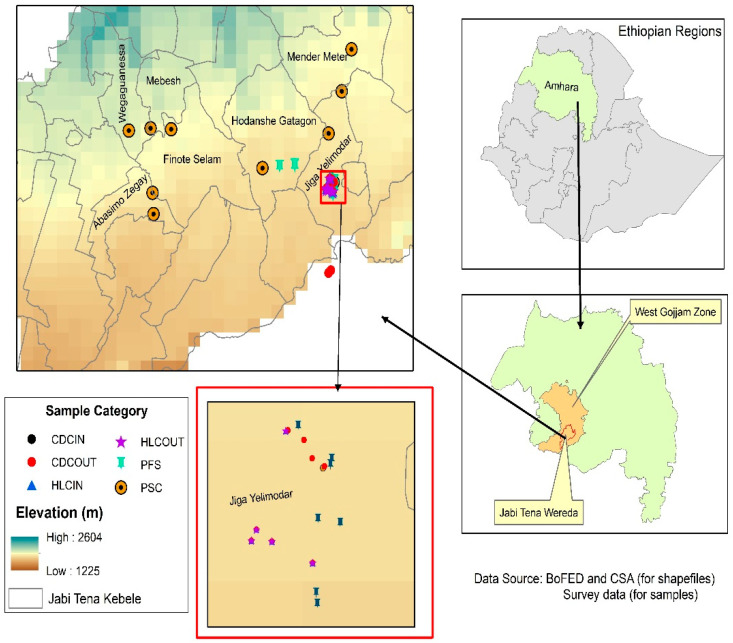 Figure 1
Figure 1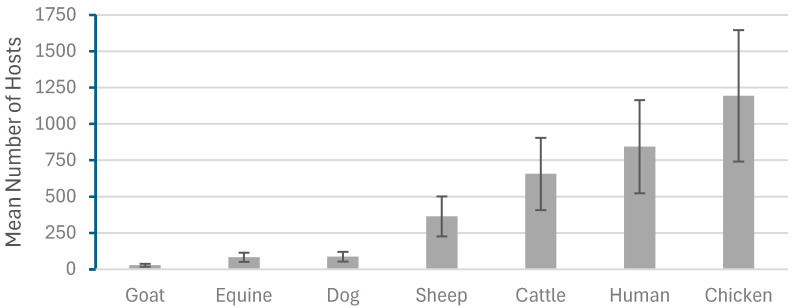 Figure 2
Figure 2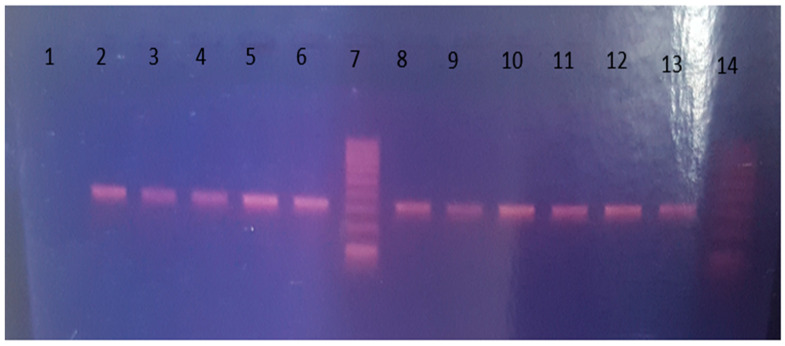 Figure 3
Figure 3- —Ministry of Health (Global fund)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Insect Pest Control Strategies · Mosquito-borne diseases and control
1. Introduction
Anopheles arabiensis and An. gambiae s.s. are the major malaria vectors in the Afrotropical region, which can survive in a range of altitudes, temperatures, and humidity conditions [1,2] and are sympatric in many regions [2,3], while those in the Anopheles funestus group serve as secondary vectors [4]. The mosquito species that breed in freshwater habitats are sympatrically distributed with other species of the An. gambiae complex [2]. An. gambiae s.l. is dominant in the humid parts of Africa with freshwater, while An. arabiensis is more widespread in the dry savannah habitats of Africa [5]. Contrarily to the findings of ecological studies that have shown that arabiensis and An. gambiae breed in clean water, recently, they have been found to survive in polluted habitats [6,7,8]. The accurate identification of malaria vector species is critical for designing and deploying efficacious tools for vector control [9]. The morphological keys developed by Gillies and Coetzee [1,10] and rigorous molecular analysis via a polymerase chain reaction (PCR) are used for the identification of sibling species in the An. gambiae and An. funestus complexes [11,12] and are important for effective malaria vector control programs [12,13].
Apart from species identification and host preference, it is important to understand the vectorial capacity and malaria transmission dynamics of Anopheles species [3,14]. Anopheles mosquitoes have a wide range of preferred hosts, including humans and animals such as cattle, sheep, horses, pigs, dogs, cats, birds, and reptiles, as well as other vertebrates and reptiles [15,16]. Anthropophilic Anopheles mosquitoes usually feed on humans, whereas zoophagic Anophelines feed mainly on bovines [17,18]. However, others are opportunistic, feeding on both humans and other animals depending on which host species is available [19].
The blood-feeding behavior of host-seeking female Anopheles mosquitoes is one of the drivers of the transmission of Plasmodium parasites in human populations during endemic malaria transmission seasons [20]. The human blood index (HBI), expressed as the proportion of mosquito blood meals that are of human origin, is an important indicator of malaria transmission and is used to estimate the human biting rate to measure the vectorial capacity and intensity of malaria transmission [21].
The sporozoite infectivity rates of mosquitoes are utilized to better understand malaria transmission intensity, vector species importance, and intervention impact [22]. The Plasmodium falciparum circumsporozoite protein for P. vivax-210 or P. vivax-247 is also used [23]. These phenomena are of paramount importance in malaria vector intervention evaluation and impact assessment [9].
Anopheles arabiensis is a main vector species of malaria in Ethiopia which has developed resistance to multiple insecticides. This is probably a result of the long-term use of indoor residual spraying (IRS) and insecticide-treated bed nets (ITNs) and their use in agriculture [24,25]. The increased multiple insecticide resistance of the main malaria vectors has raised concerns regarding insecticide-based malaria vector control interventions [26]. Insecticide resistance has been detected using molecular assays, which give early notice of resistance emergence and the underlying mechanism [3,27,28,29]. This has improved the precision of the insecticides chosen [30,31].
The relationship between the knockdown resistance (kdr) mutation, blood meal sources, and Plasmodium infection in Ethiopia is not well understood. These parameters are of paramount importance in designing appropriate approaches for effective vector control tools and programs. In Ethiopia, previous studies have identified insecticide resistance to An. arabiensis in multiple insecticide classes [32]. However, data are scarce in Jabi Tehnan District. Insecticide-based methods for controlling malaria vectors have proven to be successful in many parts of Africa [33]. Yet, because of the rise in insecticide resistance among the main malaria vectors, the fear of insecticide-based malaria vector control tools failing has grown. One of the mechanisms of the development of resistance is the knockdown resistance (kdr) mutation, which transforms Leucine (TTA) into Phenylalanine (TTT) at position 1014 of the voltage-gated sodium channel (VGSC) [32,34].
This study investigated malaria vector species diversity, blood meal source, and Plasmodium circumsporozoite rates to address challenges facing African and Ethiopian malaria control initiatives, such as insecticide resistance [35]. Establishing effective techniques for managing insecticide resistance and maintaining the effectiveness of chemical-based vector control tools can be aided by a well-coordinated national database on knockdown resistance, malaria vector species diversity and dynamics, blood meal sources, and Plasmodium circumsporozoite rates. These must be understood to create effective vector control strategies and procedures and to manage residual malaria transmission with particular care.
This study provides new insights compared with previous Ethiopian studies because it focuses on Jabi Tehnan District, a location with unique ecological, epidemiological, and intervention-related contexts that have not been extensively characterized. The district’s agricultural practices, climate variability, and rapid changes in land use create distinct vector habitats and transmission dynamics that differ from those previously documented in other regions of Ethiopia. Furthermore, the study period coincides with the intensified implementation of malaria control interventions, which included the expanded coverage of indoor residual spray (IRS), long-lasting insecticidal nets (LLINs), and new policy plans on the diagnostics and availability of first-line anti-malaria treatments. Conducting this research during a time of significant environmental change and evolving intervention strategies allows for the identification of emerging entomological and epidemiological patterns that may not have been captured in earlier studies, thereby filling a critical knowledge gap for localized malaria elimination efforts.
2. Materials and Methods
2.1. Description of Study Area and Study Design
This study was conducted in Jabi Tehnan District, a malaria-endemic area in West Gojjam Zone, Northwest Ethiopia, that is located at a distance of 387 km from Addis Ababa and 176 km southwest of Bahir Dar City (Figure 1). This area lies between 10°24′36″ and 10°55′48″ N latitude and 37°4′12″ and 37°30′36″ E longitude. This study was conducted at an altitude ranging from 1345 to 2572 masl [36]. A considerable part of the area lies in ranges closer to 2000 masl where malaria is endemic. The area receives a rainfall of about 1250 mm per annum, and the mean minimum and maximum temperatures are 14 °C and 32 °C, respectively. The rainfall distribution in the area is unimodal, and the rainy season lasts from June to mid-September [37]. The district’s total population is 225,769, of which 112,341 are males, and 113,428 are females [36,37,38].
A total of five villages were selected in Jabi Tehnan District for this study: Jiga Yelimodar (37°21′36″ E; 10°40′48″ N; altitude: 1833 m), Hodanshe Gatagon (37°19′12″ E; 10°42′00″ N; altitude: 1866 m), Abasimo Zegay (37°13′12″ E; 10°39′00″ N; altitude: 1806 m), Mender Meter (37°21′36″ E; 10°45′00″ N; altitude: 1947 m), and Mebesh (37°14′24″ E; 10°43′48″ N; altitude: 2037 m). These villages were purposively selected based on documented malaria endemicity [36,37], the representation of a range of altitudes (1806–2037 m), and ecological settings to capture variation in malaria transmission, accessibility for field surveys and logistical feasibility, and their relevance to ongoing malaria control interventions in the district.
Households were selected based on several ecological, structural, and social criteria. Proximity to mosquito breeding sites was a key factor, with priority given to houses located near known or potential Anopheles larval habitats such as streams, ponds, irrigation canals, and rice fields to maximize mosquito collection efficiency. Housing structures of varying construction types (e.g., thatched versus corrugated iron roofs, mud versus cement walls, and presence of eaves) were included to assess the influence of building characteristics on mosquito entry and resting behavior. Socio-environmental factors, including household practices and environmental features such as livestock presence, water storage practices, and vegetation cover, were also considered given their potential effect on mosquito density and human–vector contact. To avoid spatial clustering and ensure ecological representativeness, sentinel houses were distributed across each village to capture different microhabitats. All community members who provided consent and were accessible to the research team were voluntarily recruited for this study.
2.2. Survey of Vertebrate Hosts
Data on the population of vertebrate hosts residing in the study sites were provided [39]. Similar information was gathered from this study for potential hosts, including bovines, goats, dogs, and chickens [40].
2.3. Adult Mosquito Sampling Methods and Processing Procedures
The collection of adult mosquitoes was conducted from September 2016 to August 2017, using four WHO-recommended sampling techniques: pyrethrum spray catches (PSCs), human landing catches (HLCs), Centers for Disease Control (CDC) light traps, and pit shelters (PSs) [41]. Generally, these sampling methods were used for the collection of adult mosquitoes indoors and outdoors.
Five houses were selected in each sentinel village to conduct human landing catches (HLCs) and ten houses each for collection using pyrethrum spray catches (PSCs) and CDC light traps (CDC-LTs). Houses were chosen via simple random sampling from a list of all occupied households in the village after stratification by proximity to known larval habitats (within 500 m vs. >500 m), ensuring the representation of both near- and far-from-breeding-site households. The same houses were used throughout the study period unless a household became unavailable, in which case a pre-randomized replacement was enrolled.
The number of houses per method was determined by balancing spatial coverage, ethical considerations, logistical feasibility, and statistical power. HLCs were conducted at five houses per village to maximize the coverage of biting activity across settlements while minimizing collector workload and limiting exposure risk (see Ethics). PSCs and CDC-LTs were conducted at ten houses per village to provide robust estimates of indoor resting densities and trap-based abundance, complementing HLC data. Collections using different methods were conducted concurrently where possible to enable direct comparisons. The chosen sampling scheme (5 houses for HLCs and 10 houses for PSCs and CDC-LTs per village) lies within the range commonly applied in entomological surveys in Ethiopia and elsewhere (typically 3–6 houses per method per village) while providing sufficient replication across sites to ensure representative estimates of mosquito density and behavior.
2.4. Human Landing Catches (HLCs)
In each study site, five houses were randomly selected as sentinel houses for human landing catches. In each of the five houses, indoor and outdoor mosquito collections were carried out from 6:00 pm to 6:00 am for four successive nights. HLCs were carried out monthly. In each village, mosquitoes were collected from four houses, each separated by approximately 100 m. The weather data were recorded in each house that received IRS [42].
Human landing catches (HLCs) were conducted by trained collectors who were provided with mefloquine chemoprophylaxis, rather than by untrained village volunteers. This approach ensured both the safety of participants and the reliability and consistency of mosquito collection data. HLCs were conducted by an adult man exposing his lower limbs and collecting landing mosquitoes using an aspirator [41]. Each mosquito collected stayed in a station for 45 min, and there was a 15 min break for resting. To obtain hourly biting densities, the catches for each hour were divided by 0.75 [43]. In each house, a group of 4 data collectors was deployed. Once the team members arrived at the collection household, they were divided into two groups of two collectors each. One group was set to trap indoors, while the other group was outdoors. Outdoor mosquito sampling groups sat between 8 and 10 m apart from each other in all sentinel houses selected for indoor mosquito collection. There were two collection shifts: one team of collectors worked from 18:00 to 00:00 h, followed by the second team working from 01:00 to 06:00 h.
In each hour, volunteers rotated between indoors and outdoors to avoid positional bias. A full dose of mefloquine was provided as chemoprophylaxis to the volunteers who participated in human landing collections. Daily follow-up was also performed on all volunteers for any malaria symptoms and prompt access to treatment with effective anti-malarial drugs if any of the collectors were screened and then found to have malaria parasites. Captured mosquitoes were placed in labeled paper cups at hourly intervals. Thus, a total of 24 labeled paper cups (12 labeled cups for indoor and 12 labeled cups for outdoor collection) were used for each night collection per house. At the end of each collection night, all paper cups with mosquitoes were brought to the field identification center where the identification of mosquitoes to the species level using taxonomic keys [1,10] and their physiological status, unfed (UF), fed (F), half-gravid (HG), or gravid (G), was performed [41].
2.5. Pyrethrum Spray Catches
In each sentinel village, ten houses were randomly chosen for pyrethrum spray catches (PSCs). Mosquitoes were sampled from 6:00 a.m. to 7:00 a.m. from each house for two consecutive nights in all ten selected houses. Before PSCs, all the things and food in the room were covered with white sheets, and all animals were removed from the room where the collection took place. A commercially available pyrethroid-based aerosol (Baygon aerosol, SC. Johnson & Son Inc., Racine, WI, USA) was sprayed in the entire space of the room, and the house was then closed for 15 min after spraying. After fifteen minutes, all the knocked-down mosquitoes on the white sheets were collected carefully with forceps and placed in paper cups [41]. All Anopheles mosquitoes collected on the white cotton sheet were identified to species using taxonomic keys, and the abdominal stage of each mosquito was also determined as unfed, freshly fed, half-gravid, or gravid [41].
2.6. CDC Light Traps (CDC-LTs)
CDC miniature light traps (Gladwick St., Rancho Dominguez, CA, USA, and J.W. Hock Ltd., Gainsville, FL, USA) were powered by a GS premium high-power (6N11-2D; 6v-11Ah/10Hr Japan storage battery Co., Ltd., Kyoto, Japan), rechargeable battery. The battery was set near an occupied bed at a height of 1.5 above the floor from 18:00 to 06:00 h to sample indoor host-searching mosquitoes [41]. For outdoor host-seeking mosquito sampling, a CDC light trap was also set in the vicinity (within 2 m) of sentinel houses from 18:00 to 06:00 h. Ten houses with paired traps indoors and outdoors in the same village were selected with varied vicinities to breeding. Mosquitoes were collected indoors and outdoors from 18:00 to 06:00 h from each house using CDC light traps. Mosquito collection bags were retrieved from traps in the morning between 06:00 h and 07:00 h. Mosquitoes were identified using morphological features [1,10] and classified based on abdominal status (blood-fed, empty (unfed), semi-gravid, and gravid) [41].
2.7. Pit Shelter Collections
Outdoor resting mosquitoes were collected monthly between 06:00 and 09:00 h from ten pit shelters made in selected house compounds. A rectangular pit was dug in the ground (1.5 m in depth, 1.2 m in length, and 1 m in width). Mosquitoes were sampled monthly using a mouth-suction aspirator according to the method stated in the WHO entomological manual in 1975 [41]. The captured mosquitoes were anesthetized using chloroform. The pit shelters were covered with an untreated net during collection to prevent mosquitoes from escaping [41].
2.8. Molecular Identification of Anopheles gambiae s.l.
DNA was extracted from the legs and wings of the identified female Anopheles mosquitoes using the DNeasy Blood and Tissue Kit using the manufacturer’s protocol (Qiagen^®^, Sigma Aldrich, St. Louis, MO, USA). The extracted DNA was stored at −20 °C until it was used for molecular analysis. Then, it was subjected to molecular identification using a species-specific polymerase chain reaction (PCR) assay following the methods of Scott and others [12], at the Molecular Biology Laboratory of Tropical and Infectious Diseases Research Centre (TIDRC), Sekoru, Jimma University. Briefly, PCR amplification was performed in a final reaction of 20 uL using a set of each primer with a 0.25 µM final concentration (UN: 5′-GTGTGCCCCTTCCTCGATGT-3′) and species-specific primers for An. gambiae s.l (GA: 5′-CTGGTTTGGTCGGCACGTTT-3′), An. arabiensis (AR: 5′-AAGTGTCCTTCTCCA TCCTA-3′), 7.5 µL nuclease-free water, and 1 µL template DNA. The PCR program was set for an initial step at 94 °C/10 min, 30 cycles of 94 °C/30 s, 50 °C/30 s, 72 °C/30 s, and final extension at 72 °C/5 min. Then, the amplicon was loaded on 2% agarose gel stained with 1% ethidium bromide, and a 100 bp reference DNA ladder was used. Finally, the band size of PCR products was compared to the An. arabiensis-susceptible colony strain used as a positive control for confirmation during visualization on a Bio-Rad-UV-gel documentation system.
2.9. Detection of Knockdown Resistance Gene Mutation
An analysis of the knockdown resistance gene (L1014F) mutation was carried out on 251 An. arabiensis (PCR-identified) samples as described in previous reports conducted in different areas with species identifications [44,45]. Briefly, genomic DNA extracted from individual mosquitoes was genotyped using allele-specific primers [44]. The amplicon was run on a 2% agarose gel and stained with 3 µL of 1% ethidium bromide. The resulting fragments (bands) were interpreted as 293 bp internal control, 195 bp resistant, and 137 bp susceptible/wild-type mosquitoes [13,44]. Susceptible An. arabiensis strains were taken from the Sekoru insectary colony of Jimma University, Tropical and Infectious Diseases Research Center, Ethiopia, and used as a control.
2.10. Analysis of Mosquito Blood Meal Source
The origins of the blood meals of all freshly fed mosquitoes and a subset of half-gravid Anopheles mosquitoes (An. arabiensis) were identified using ELISA. Adult mosquitoes were collected from a malaria-endemic village, and blood-fed specimens were analyzed following the method of Beier et al. [46].
Briefly, the abdomen of each freshly blood-fed mosquito was separated from the head–thorax and homogenized in phosphate-buffered saline (PBS) using a pestle in a 1.5 mL Eppendorf tube. The homogenate was diluted 1:50 with PBS, and 50 μL of the diluted sample was added to U-shaped microplate wells. After incubation at room temperature for 3 h, the homogenate was discarded, and the wells were washed twice with PBS–Tween 20. Subsequently, 50 μL of peroxidase-conjugated human antibodies and 50 μL of phosphatase-conjugated bovine antibodies were added to the respective wells. Plates were then covered and incubated for 1 h at room temperature, followed by three washes with PBS–Tween 20. Finally, 100 μL of ABTS peroxidase substrate solution was added, and absorbance was measured using an ELISA reader at 405–415 nm after 30 min. Positive controls consisted of cattle and human blood dried on Whatman paper, while negative controls included unfed An. arabiensis from an insectary colony at ALIPB.
2.11. Determination of Plasmodium Sporozoite Rates
The head and thorax of preserved An. gambiae s.l. mosquitoes were carefully separated from the abdomen and processed for the detection of the circumsporozoite protein for both P. falciparum and P. vivax using an enzyme-linked immunosorbent assay (CSP-ELISA) [47]). The wavelength absorbance was measured at 405 nm using a UV-assisted spectrophotometer ELISA reader (SpectraMax i3x (Syngene International Ltd., Baltimore, MD, USA)). The sporozoite rate was estimated as the number of mosquitoes with sporozoites divided by the number of females examined multiplied by 100 [48].
2.12. Data Analysis
The human blood index (HBI) and bovine blood index (BBI) were calculated as the ratio of blood-fed mosquitoes that had fed on humans and cattle, respectively, relative to the total number of mosquitoes tested. The mixed blood meal source was determined as the ratio of blood-fed mosquitoes that had fed on both human and cattle blood to the total tested represented as a percentage. The unknown blood meal source represented the proportion of blood-fed mosquitoes with unidentified blood origins, also expressed as a percentage [14]. The sporozoite rate (SR) was calculated as the number of mosquitoes positive for circumsporozoite protein (CSP) antigens divided by the total number examined, expressed as a percentage [48,49].
3. Results
3.1. Survey of Vertebrate Hosts in Study Site
A total of 19,520 potential vertebrate hosts were recorded in the study villages (Table 1), including cattle, goats, sheep, dogs, chickens, equines, and humans, which varied statistically significantly (F = 23.74, df = 6, p > 0.001) (Figure 2).
3.2. Anopheles Mosquito Species Compositions and Abdominal Status Using Different Sampling Techniques
A total of 5098 female Anopheles mosquitoes belonging to five species were collected using four sampling techniques: CDC light traps (1423, 27.9%), human landing catches (HLCs) (1690, 33.2%), pyrethrum spray catches (PSCs) (1635, 32.1%), and pit shelters (PSs) (350, 6.9%). Of these mosquitoes, 57.2% (n = 2915) were sampled indoors using PSCs, HLCs, and CDC-LTs, while 42.8% (n = 2183) were sampled outdoors using HLCs, CDC-LTs, and PSs (Table 2).
The examination of abdominal status showed that the majority of mosquitoes were unfed (2792, 54.8%), followed by freshly fed (2126, 41.7%), gravid (147, 2.9%), and half-gravid (33, 0.6%). Species-specific analysis indicated that An. arabiensis was the most abundant (2982, 58.5% of the total), with a high proportion of unfed and freshly fed females, suggesting active host-seeking and blood-feeding behavior. Other species collected included An. coustani (774, 15.2%), An. cinereus (561, 11.0%), An. pharoensis (605, 11.9%), and An. funestus (176, 3.5%).
The distribution of abdominal status across sampling methods showed that HLCs captured the highest number of host-seeking (unfed) mosquitoes outdoors, while PSCs and CDC-LTs captured primarily indoor-resting freshly fed mosquitoes. Pit shelters mainly collected outdoor-resting mosquitoes with a mix of those of unfed and freshly fed status (Table 2). These patterns highlight the behavioral diversity of vector species and emphasize the importance of using multiple sampling methods to accurately assess malaria transmission potential.
3.3. Molecular Identification of Anopheles gambiae s.l. Mosquitoes
Among the 530 An. gambiae s.l. samples analyzed using species-specific PCR, 509 (96.03%) were An. arabiensis, while the remaining 21 (3.97%) of the samples were not amplified (unknown) (Figure 3).
3.4. Detection of Knockdown Resistance Gene Mutation
The presence of the L1014F (West African kdr) allelic mutation was investigated in 251 PCR-confirmed An. arabiensis mosquitoes, of which 84.5% (n = 212) were successfully amplified, and 15.5% (n = 39) were unamplified. The knockdown resistance gene distribution consisted of 1.4% (n = 3) heterozygous resistant (RS), 16.3% (n = 38) homozygous resistant (RR), and 68.13% (n = 171) homozygous susceptible (SS) genotypes, and 15.54% (n = 39) were undetected (NB) (Table 3).
3.5. Blood Meal Sources of An. arabiensis Mosquitoes
The human blood index (HBI) and bovine blood index (BBI) and the mixed-blood-fed and unknown blood meal sources of An. arabiensis are presented in Table 4. Out of 1475 freshly fed field-caught An. arabiensis, 360 (24.4%) mosquitoes were tested for single-host blood meals, mixed blood meals, and unknown blood meal sources. For blood meal analysis, the following vertebrate antisera were used: human and bovine. These were chosen based on the common domestic hosts present in the study villages, allowing for the determination of the host-feeding patterns of Anopheles arabiensis. Among the tested mosquitoes, 13.1% (n = 47) of blood meals were of human origin, 8.9% (n = 32) were bovine, and 5.8% (n = 21) were mixed (human and bovine), while the majority, 72.2% (n = 260), were from unidentified sources. Mosquitoes collected with pyrethrum spray catches (PSCs) exhibited a relatively higher HBI (17.7%, n = 23) and BBI (9.2%, n = 12) compared to those captured using human landing catches (HLCs), CDC light traps (CDC-LTs), and pit shelters (PSs).
3.6. Plasmodium Circumsporozoite Protein Detection in Anopheles Mosquitoes
A total of 500 An. arabiensis and 20 An. pharoensis mosquitoes were assessed for the presence of Plasmodium falciparum and P. vivax circumsporozoite protein (CSP). No mosquitoes tested positive for P. falciparum CSP; therefore the entomological inoculation rate (EIR) was not calculated.
4. Discussion
The findings of this study showed that understanding blood meal sources, the detection of knockdown genes, and species composition is of paramount importance for the effective design of the malaria vector control programs and tools to be used. The understanding of mosquito host choice behavior during blood meal analysis was found to be crucial in providing information on how vectors transmit pathogens between vertebrates. In natural settings, mosquitoes exhibit varied feeding succession due to host availability and attractiveness [50]. The feeding patterns and host associations of mosquitoes are influenced by both environmental and biological factors [51]. Female mosquitoes ingest vertebrate blood to produce eggs and then transmit pathogens during blood feeding, and those that feed on multiple hosts like humans and other hosts accelerate malaria transmission by shortening the incubation period [52]. In Ethiopia, the primary vector feeding on human blood is An. arabiensis, with great implications for malaria transmission, followed by An. stephensi, and these vectors must be tackled to control and eliminate them [53]. In female An. arabiensis, after blood feeding, their abdomens become engorged, which affects their flight capacity, host-seeking behavior, and patterns, while half-gravid mosquitoes prefer resting sites like suitable water resources to lay eggs [54]. In this study, we performed laboratory investigations on 500 An. arabiensis and 20 An. pharoensis mosquitoes to determine the mosquito infectivity rate, which refers to the proportion of those carrying infectious plasmodium sporozoites, using the sandwiched enzyme linked immunosorbent assay method. None of the analyzed mosquitoes were positive for either Plasmodium falciparum or Plasmodium vivax infections, considering the limitations of the method, and due to this, we were unable to determine the entomological inoculation rate (EIR). However, this may not represent the real situation of the study area, and therefore large-scale surveillance is recommended as vector density and malaria parasite incidence are resurging.
This study provides a comprehensive assessment of Anopheles mosquito species composition, abdominal status, host-feeding behavior, and infection potential in Jabi Tehnan District, Northwest Ethiopia. A total of 5098 female mosquitoes were collected, with An. arabiensis being the most abundant (2982, 58.5%), followed by An. coustani (15.2%), An. pharoensis (11.9%), An. cinereus (11.0%), and An. funestus (3.5%). An analysis of abdominal status revealed that most mosquitoes were unfed (54.8%) or freshly fed (41.7%), indicating active host-seeking and recent human–vector contact, while half-gravid and gravid females were relatively sparse, suggesting pre-oviposition behavior.
Indoor versus outdoor collections showed that 57.2% of mosquitoes were captured indoors using PSCs, CDC light traps, and HLCs, while 42.8% were collected outdoors using HLCs, CDC-LTs, and pit shelters. This indicates the presence of both endophagic/endophilic and exophagic/exophilic vectors. These patterns highlight the importance of integrated vector control strategies that target multiple ecological niches and both indoor and outdoor biting populations.
The laboratory testing of 500 An. arabiensis and 20 An. pharoensis mosquitoes revealed no CSP-positive mosquitoes; hence the calculation of the entomological inoculation rate (EIR) was not possible. This absence may reflect seasonality, limited sample size, or assay sensitivity, rather than a true lack of malaria transmission. Coupled with the high abundance of host-seeking and recently fed mosquitoes, these findings suggest that transmission potential remains in the area. Blood meal behavior further demonstrates that vectors feeding on multiple hosts, including humans, can accelerate malaria transmission by shortening the parasite incubation period.
The 17.9% frequency of resistance alleles suggests moderate insecticide resistance in the local vector population, comparable to other mid-altitude Ethiopian sites (10–25%) and lower than in some East African regions (>40%) [55]. A 15.5% PCR amplification failure rate is unlikely to significantly bias this estimate if the failures were randomly distributed, but ongoing monitoring is essential to detect changes that may compromise LLIN and IRS effectiveness.
The findings of this study highlighted the areas to focus on to strengthen indoor interventions such as LLINs and IRS. Scaling up blood meal and sporozoite monitoring will help guide timely and evidence-based interventions. Overall, this study underscores the need for integrated, evidence-based vector control strategies that account for species diversity, feeding behavior, abdominal status, insecticide resistance, and both indoor and outdoor transmission dynamics to effectively reduce malaria risk in the district.
This study identified five Anopheline species morphologically. These are An. arabiensis, An. funestus, An. pharoensis, An. coustani, and Anopheles cinereus, which were found to be similar to those in earlier studies in other parts of the region [56,57]. Among these, the An. arabiensis species was confirmed by a molecular test to be able to be used to determine the frequency of insecticide resistance. The large-scale use of insecticide-treated nets (ITNs) and indoor residual spraying is associated with an increased number of cases, and management led to a remarkable reduction in malaria cases and vector control up to early 2019 in Ethiopia [58]. In most African countries, following the massive use of insecticides, insecticide-resistant vectors are most commonly seen in malaria-endemic settings [59]. According to a study by Karunaratne and others, the metabolism and longevity of insecticide-resistant An. gambiae are lower than those of the susceptible strain, and the resistant stain showed a higher level of oxygen reactive species (ROS), which are key factors in determining oxidative stress. The major effects of insecticide resistance in mosquitoes are metabolic changes, altered target sites, and cuticular and behavioral resistance [60]. Overall, mosquitoes that develop the capacity to cope with oxidative stress are likely to live longer. According to an experimental investigation study by Oliver and Brooke, evaluating the effects of oxidative stress on the longevity of both An. arabiensis and An. funestus bearing, respectively, kdr and cytochrome P450 mechanisms demonstrates that these species live longer and that cytochrome P450 activity seems to be more protective against oxidative stress [61]. In our study, among the An. arabiensis species confirmed by a molecular test, 251 genomic isolates were tested for kdr, and the frequency was determined, as indicated in Table 3. Thus, we detected L1014F (West African kdr) allelic variant point mutations in 17.9% of the An. arabiensis mosquitoes genotyped, and over 80% were the susceptible strain. This study provides valuable insights into and information about the mosquito infectivity rate, blood-feeding patterns, mosquito density, and allelic frequency of insecticide resistance in the study setting.
The reported 17.9% frequency of the resistance allele in the study area indicates a moderate level of insecticide resistance within the local Anopheles population. This value is comparable to previous findings in mid-altitude Ethiopian sites, where resistance allele frequencies ranged from 10% to 25% [62,63], and is lower than the frequencies reported in certain high-transmission areas of East Africa, such as western Kenya and Tanzania, where frequencies often exceed 40–50% [64,65]. The observed allele frequency suggests that while resistance is present, it has not yet reached fixation in the local population. Continuous monitoring is warranted, as rising resistance levels could compromise the effectiveness of indoor-based interventions such as LLINs and IRS. Moreover, these findings emphasize the need for integrated vector management strategies, including the rotation of insecticides and complementary control measures, to mitigate further selection pressure.
This study found that 15.5% of all specimens failed to be amplified to the species level during molecular analysis, which could be due to factors such as degraded DNA, insufficient template quantity, or suboptimal PCR conditions. Also, the reported presence of the invasive mosquito species An. stephensi might be the cause as it is already widely reported in Ethiopia [66,67,68,69]. These “not amplified” samples may introduce potential bias in estimating resistance allele frequencies if their distribution differs from that of successfully amplified samples. For instance, if non-amplified mosquitoes disproportionately carried the resistance allele or belonged to a specific species, the reported 17.9% allele frequency could underestimate or overestimate the true prevalence. However, assuming the failures occurred randomly across species and collection sites, the impact on overall allele frequency estimates is likely minimal. Nevertheless, reporting the amplification failure rate and considering it in data interpretation are important to provide an accurate assessment of resistance in the local vector population.
Overall, we recommend that combination efforts targeting vector bionomics and parasite clearance and the large-scale molecular surveillance of insecticide resistance are very important for the management of vector and malaria parasite incidence, which is resurging in Ethiopia.
5. Conclusions
This study gives valuable insights into malaria transmission dynamics and vector control strategies. Blood-feeding patterns influence malaria transmission dynamics as An. arabiensis is the primary vector feeding on human blood in Ethiopia. Thus, insecticide-resistant vectors are common in malaria-endemic regions, and large-scale molecular surveillance is recommended for accurate vector density, insecticide resistance, and malaria incidence assessment.
To strengthen future blood meal analyses and reduce the proportion of unidentified samples, studies should expand the panel of tested hosts to include locally relevant species, adopt molecular approaches such as cytochrome b or 16S rRNA sequencing for higher sensitivity, and improve field preservation protocols. These efforts, complemented by contextual household or ecological data on host availability, would provide a more accurate understanding of vector feeding behavior.
A substantial proportion of blood meals in this study were classified as “unknown”, which limits our ability to fully characterized host-feeding patterns. Several factors may have contributed to this. First, the antiserum panel used (human, bovine, goat, chicken) did not cover all vertebrate species present in the study area—for example, dogs, sheep, donkeys/horses, and a range of peri-domestic or wild mammals (rodents, small carnivores) were not tested and could account for a proportion of the unidentified meals. Second, blood meal degradation (due to the time between mosquito capture and sample preservation or suboptimal storage/transport conditions) can reduce the sensitivity of immunoassays and produce indeterminate results. Third, mixed blood meals may cause weak or ambiguous reactivity in ELISA-based assays, and cross-reactivity between closely related species can mask true host identity. Finally, the technical limitations of immunoassays (limits of detection, lot variability in antisera) can also increase the “unknown” fraction.
Because the unknown category is non-random, the observed host-feeding proportions likely underrepresent certain hosts and therefore should be interpreted cautiously. Conclusions about the relative importance of humans versus domestic animals as blood meal sources may be biased if a sizeable portion of anthropophagy or zoophagy is hidden within the unknowns. In particular, vector control recommendations that assume a low level of feeding on certain domestic animals should be made tentatively if these animals were not included in the assay panel.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Coetzee M. Key to the females of Afrotropical Anopheles mosquitoes (Diptera: Culicidae)Malar. J.2020197010.1186/s 12936-020-3144-932054502 PMC 7020601 · doi ↗ · pubmed ↗
- 2Coetzee M. Craig M. Le Sueur D. Distribution of African malaria mosquitoes belonging to the Anopheles gambiae complex Parasitol. Today 200016747710.1016/S 0169-4758(99)01563-X 10652493 · doi ↗ · pubmed ↗
- 3Kweka E.J. Mazigo H.D. Lyaruu L.J. Mausa E.A. Venter N. Mahande A.M. Coetzee M. Anopheline mosquito species composition, kdr mutation frequency, and parasite infectivity status in northern Tanzania J. Med. Entomol.20205793393810.1093/jme/tjz 24531923308 · doi ↗ · pubmed ↗
- 4Mustapha A.M. Musembi S. Nyamache A.K. Machani M.G. Kosgei J. Wamuyu L. Ochomo E. Lobo N.F. Secondary malaria vectors in western Kenya include novel species with unexpectedly high densities and parasite infection rates Parasit. Vectors 20211425210.1186/s 13071-021-04748-933980273 PMC 8117294 · doi ↗ · pubmed ↗
- 5Sinka M.E. Bangs M.J. Manguin S. Rubio-Palis Y. Chareonviriyaphap T. Coetzee M. Mbogo C.M. Hemingway J. Patil A.P. Temperley W.H. A global map of dominant malaria vectors Parasit. Vectors 201256910.1186/1756-3305-5-6922475528 PMC 3349467 · doi ↗ · pubmed ↗
- 6Hinne I.A. Attah S.K. Mensah B.A. Forson A.O. Afrane Y.A. Larval habitat diversity and Anopheles mosquito species distribution in different ecological zones in Ghana Parasit. Vectors 20211419310.1186/s 13071-021-04701-w 33827667 PMC 8025514 · doi ↗ · pubmed ↗
- 7Mireji P.O. Keating J. Hassanali A. Impoinvil D.E. Mbogo C.M. Muturi M.N. Nyambaka H. Kenya E.U. Githure J.I. Beier J.C. Expression of metallothionein and alpha-tubulin in heavy metal-tolerant Anopheles gambiae sensu stricto (Diptera: Culicidae)Ecotoxicol. Environ. Saf.201073465010.1016/j.ecoenv.2009.08.00419735939 PMC 2783303 · doi ↗ · pubmed ↗
- 8Mireji P.O. Keating J. Hassanali A. Mbogo C.M. Nyambaka H. Kahindi S. Beier J.C. Heavy metals in mosquito larval habitats in urban Kisumu and Malindi, Kenya, and their impact Ecotoxicol. Environ. Saf.20087014715310.1016/j.ecoenv.2007.03.01217532467 PMC 2673497 · doi ↗ · pubmed ↗