First Investigation of Grass Carp Reovirus (GCRV) Infection in Amphioxus: Insights into Pathological Effects, Transmission, and Transcriptomic Responses
Jingyuan Lin, Meng Yang, Huijuan Yang, Guangdong Ji, Zhenhui Liu

TL;DR
This study shows that Grass Carp Reovirus can infect and damage amphioxus gills, spread through water, and trigger specific immune responses.
Contribution
First investigation of Grass Carp Reovirus infection in amphioxus, revealing its pathogenicity and waterborne transmission.
Findings
GCRV causes severe gill tissue damage in amphioxus when exposed to the virus.
GCRV can be transmitted via water, as healthy amphioxus showed infection after co-culturing with infected individuals.
Transcriptomic analysis revealed activation of the MAPK, calcium signaling, and chitin metabolic pathways in response to GCRV.
Abstract
Amphioxus belongs to the subphylum Cephalochordata and occupies a transitional position in evolution between invertebrates and vertebrates. Due to the lack of viruses suitable for immunostimulation in amphioxus, this study for the first time explored the pathogenicity and waterborne transmission of Grass Carp Reovirus (GCRV), a double-stranded RNA virus, during its infection of amphioxus. Soaking amphioxus in GCRV suspension can cause obvious damage to gill tissues and severely disrupt the structure of gill filaments. The virus survived in seawater for no more than 48 h. Infection kinetics studies showed that the expression of VP5 (a viral capsid protein) mRNA in gill tissues peaked at 14 h. After co-culturing GCRV-infected amphioxus with healthy amphioxus for 72 h, the gills of healthy amphioxus showed obvious pathological damage. Additionally, the presence of the virus was verified by…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
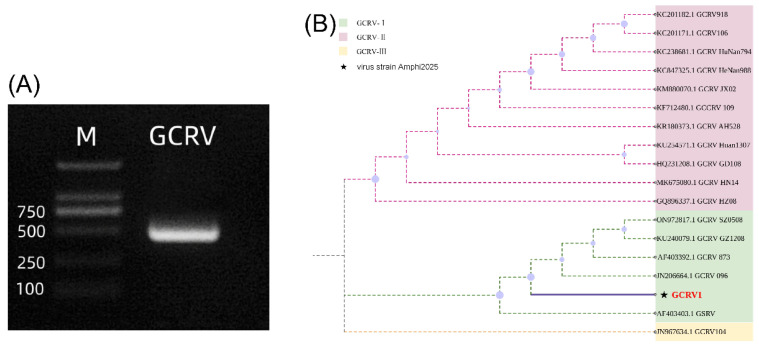 Figure 1
Figure 1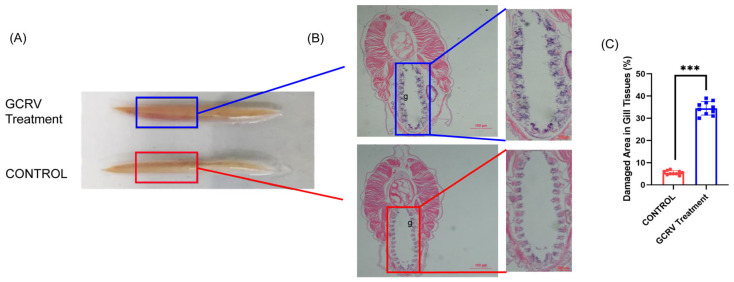 Figure 2
Figure 2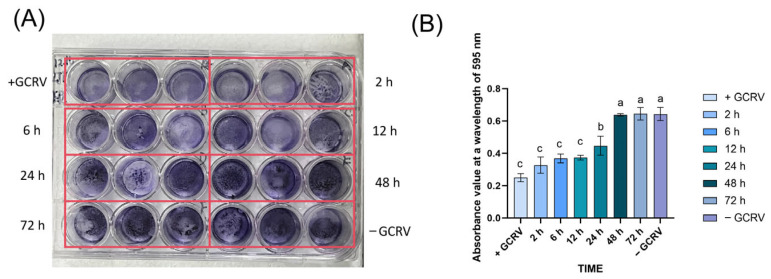 Figure 3
Figure 3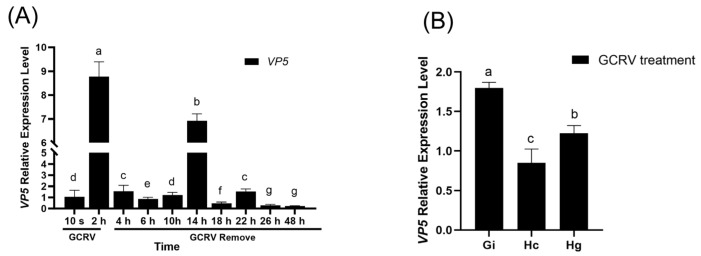 Figure 4
Figure 4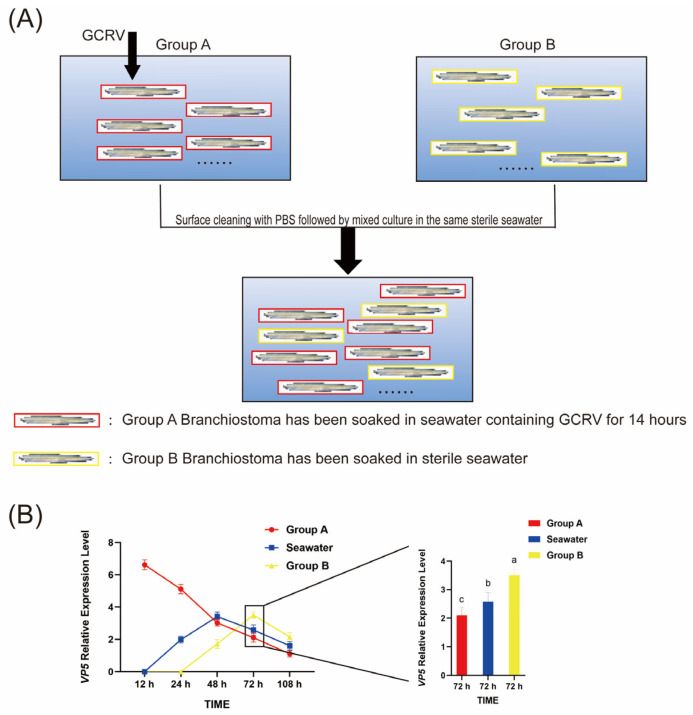 Figure 5
Figure 5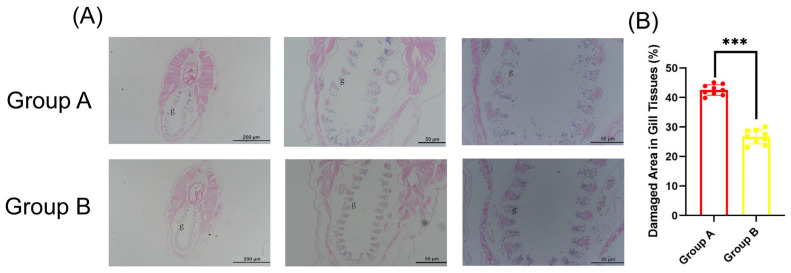 Figure 6
Figure 6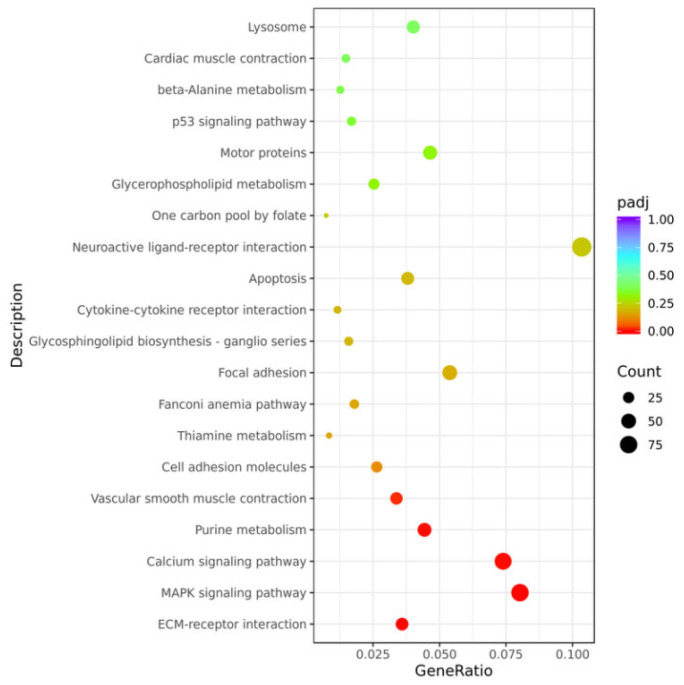 Figure 7
Figure 7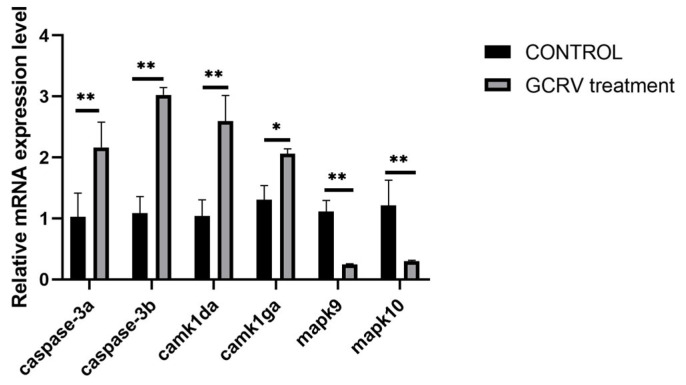 Figure 8
Figure 8- —National Natural Science Foundation of China
- —National Key Research and Development Project of the Ministry of Science and Technology
- —Science & Technology Innovation Project of Laoshan Laboratory
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAquaculture disease management and microbiota · Aquaculture Nutrition and Growth · Physiological and biochemical adaptations
1. Introduction
Grass Carp Reovirus (GCRV) is a double-stranded RNA aquatic reovirus. According to the differences in its VP4 gene sequences, it can be divided into types I, II, and III. Among these genotypes, Genotype II (GCRV-II) has emerged as the predominant pathogenic genotype in current grass carp farming systems. It is widely distributed across major aquaculture regions and exhibits high virulence to both grass carp fry and adults—frequently triggering outbreaks of hemorrhagic disease, with mortality rates in fry often exceeding 80%. By contrast, Genotype I (GCRV-I), typified by the classic strain GCRV-873, was once the dominant pathogen responsible for early outbreaks of grass carp hemorrhagic disease [1]. As for Genotype III (GCRV-III), it has been rarely detected in recent epidemiological surveys and is regarded as having relatively limited pathogenicity in commercial grass carp aquaculture [2]. Currently, research on GCRV mainly focuses on fish hosts, covering areas such as genotype identification [3], functions of pathogenic proteins (e.g., VP4-mediated cellular adsorption) [4], and vaccine development [5]. GCRV can be transmitted via water bodies, and upon infection, it targets key host tissues including the gills, liver, and kidneys [6]. However, whether the host range of GCRV as an aquatic virus is limited to cyprinid fish, especially whether it can break through species barriers to infect non-fish aquatic organisms, still requires further investigation [7].
Amphioxus is a cephalochordate animal that bridges invertebrates and vertebrates. Due to its retention of primitive characteristics from vertebrate ancestors, it is regarded as a “living fossil” for studying vertebrate immune evolution [8]. Amphioxus lacks specific immune cells and antibodies, relying on innate immune mechanisms to defend against pathogens [9]. However, no viruses capable of infecting amphioxus have been identified thus far. The aim of this study are to investigate whether GCRV (used in this paper is Genotype I, with the strain designated as GCRV-Amphi2025) can infect amphioxus; and to confirm whether GCRV can be transmitted between amphioxus individuals through water.
2. Materials and Methods
2.1. Experimental Animals and Virus Strains
Amphioxus (Branchiostoma japonicum) was bred by our laboratory. Body mass was 1.0 ± 0.2 g. They were kept in a seawater system and water temperature of 22 ± 1 °C.
Grass Carp Reovirus (GCRV) used in this study is a double-stranded RNA (dsRNA) virus belonging to genotype I. This strain, designated as GCRV-Amphi2025 in the present study, was kindly provided by Professor Yibing Zhang’s research group at the Institute of Hydrobiology, Chinese Academy of Sciences (IHB, CAS).
2.2. Infection Modeling and Sample Collection
Amphioxus were fasted for 24 h in advance. The acclimated amphioxus were divided into 2 groups (experimental group and control group), with n = 30 per group (3 biological replicates, n = 10 per replicate). Each replicate was placed in an incubator containing 200 mL of sterile seawater. For the experimental group, 40 mL of GCRV (10^5^ TCID_50_) was added; the control group received an equal volume of cell culture medium.
Samples were collected at 10 s, 2, 4, 6, 10, 14, 18, 22, 26, and 48 h after viral stimulation, respectively. At each time point, 3 biological replicate samples were collected per group (1 replicate per incubator), and each replicate included 3 amphioxus. Gill, hepatic cecum, and hindgut tissues were dissected from each amphioxus: 1 part was fixed with 4% paraformaldehyde (for histopathology), and the other was stored at −80 °C (for total RNA extraction, with 3 amphioxus’ tissues per replicate pooled as 1 RNA sample to ensure sufficient material).
2.3. qPCR Analysis
Total tissue RNA was extracted using the TRIzol method, and its purity was detected by NanoDrop 2000 (Thermo Fisher Scientific, Waltham, MA, USA). The RNA was reverse-transcribed into cDNA using the Total RNA Kit I (OMEGA, #R6834-01, Norcross, GA, USA). The expression of the VP5 gene was detected by the SYBR Green method (primers: VP5-F: 5′-CTCCCCGTGAGCGTGTATTT-3′, VP5-R: 5′-GTTAGCAGCGGTAGTGACTTG-3′). The reaction conditions were as follows: pre-denaturation at 95 °C for 30 s; 40 cycles of denaturation at 95 °C for 5 s and annealing-extension at 60 °C for 30 s. Each cDNA sample was analyzed with 3 biological replicates (corresponding to 3 tissue pools in Section 2.2) and 3 technical replicates per biological replicate. The relative expression levels were calculated using the ΔΔCt method.
2.4. Histopathology
Fixed tissues were dehydrated with gradient ethanol, embedded in paraffin, prepared as 5 μm sections, stained with HE staining and observed under the microscope. ImageJ software (version 1.54q) was used to measure the percentage of gill filament epithelial cell detachment area: 5 random fields were selected per section, and the average value of 3 sections per biological replicate was used for statistical analysis.
2.5. Transcriptome Sequencing
Gill tissues at 12 h post-infection and corresponding control samples (n = 3 biological replicates) were selected and entrusted to Novozymes for Illumina NovaSeq 6000 sequencing (PE150, single-sample data volume ≥ 6 Gb). After quality control by FastQC (version 0.11.9), raw data were aligned to the amphioxus reference genome using Hisat2 (version 2.2.1). Differential genes were screened by DESeq2 (version 1.30.1, FC ≥ 2, padj < 0.05), and GO/KEGG enrichment analysis was performed using the clusterProfiler package (version 4.16.0, Benjamini-Hochberg correction).
2.6. Virus Survival Assay
The virus suspension at 10^5^ TCID_50_/mL was added to sterile seawater at room temperature (25 °C). Samples were collected at 0, 2, 6, 12, 24, and 48 h. TCID_50_ was determined by EPC cytopathic effect assay, with 3 replicates set for each time point.
The Epithelioma Papulosum Cyprini (EPC) cell line was used for GCRV infectivity detection. EPC cells were cultured in Minimum Essential Medium (MEM) supplemented with 10% (v/v) heat-inactivated fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μg/mL streptomycin. The cell culture environment was maintained at 28 °C with 5% CO_2_, and cells were passaged every 48–72 h when reaching 80% confluency to ensure logarithmic growth phase during the experiment.
Virus-seawater mixture preparation: The GCRV-Amphi2025 stock (10^7^ TCID_50_/mL, prepared by propagating in EPC cells as described above) was diluted with sterile filtered seawater (pre-sterilized by autoclaving) to a final concentration of 10^5^ TCID_50_/mL. The mixture was incubated at room temperature (25 ± 0.5 °C) in a sealed sterile centrifuge tube, with 3 biological replicates per time point.
Sample collection and pretreatment: At 0, 2, 6, 12, 24, and 48 h post-incubation, 1 mL of the virus-seawater mixture was collected from each replicate. Samples were centrifuged at 4 °C, 5000× g for 10 min to remove potential impurities, and the supernatant was collected and stored at −80 °C temporarily (tested within 24 h to avoid viral inactivation).
TCID_50_ determination via cytopathic effect (CPE) assay: EPC cells in logarithmic growth phase were seeded into 96-well cell culture plates at a density of 2 × 10^4^ cells/well, and cultured at 28 °C with 5% CO_2_ for 24 h until cells formed a uniform monolayer. The pretreated virus supernatants were serially diluted 10-fold with MEM containing 2% FBS (from 10^−1^ to 10^−8^), and the virus-free sterile seawater was used as the negative control. The original medium in the 96-well plate was discarded, and 100 μL of the diluted virus solution (or control solution) was added to each well, with 8 technical replicates per dilution. After incubating at 28 °C with 5% CO_2_ for 1 h (to allow viral adsorption), 100 μL of MEM containing 2% FBS was added to each well, and incubation was continued for 72 h. CPE was observed daily under an inverted microscope, and the wells showing typical CPE (cell rounding, detachment, and aggregation) were recorded. The TCID_50_ value was calculated using the Reed-Muench method to quantify the viral infectivity titer at each time point [10].
Data validation: For each time point, the TCID_50_ value was considered valid only if the negative control wells showed no CPE, and the coefficient of variation (CV) among the 3 biological replicates was less than 15%.
2.7. Waterborne Transmission Experiment
Infected amphioxus (Group A: soaked in GCRV for 14 h) and healthy amphioxus (Group B: soaked in sterile seawater) were co-cultured in sterile seawater. Each group had n = 15 amphioxus, divided into 3 biological replicates (n = 5 per replicate). Samples were collected at 12, 24, 48, 72, and 108 h post-co-culture: 3 biological replicate samples were collected per group at each time point, with 1 amphioxus per replicate. Total RNA was extracted for qPCR detection of VP5 gene expression (with 3 technical replicates per RNA sample). At 72 h, histopathological analysis was performed (3 biological replicates per group, 3 sections per replicate, consistent with 2.4).
2.8. Statistical Analysis
Student’s t-test (two-group comparison) or One-way ANOVA (multi-group comparison, Tukey’s post-hoc test) was performed using SPSS 26.0, GraphPad Prism 9 plotting, and data were presented as “Mean ± Standard Deviation (SD)” based on at least 3 biological replicates and 3 technical replicates (for qPCR, TCID_50_, and histopathology). p < 0.05 was considered a significant difference.
3. Results
3.1. The GCRV Strain Was Identified as Type I
GCRV can be classified into types I, II, and III, which can be identified by the triple PCR method [11]. In this study, this method was used to identify the type of GCRV (GCRV-Amphi2025) used in the experiment. The results showed that only a specific 532 bp band was amplified with the type I primer (P01-F/R) (Figure 1A), while no amplification products were obtained with the type II and type III primers. Full-length sequencing of the VP4 gene revealed that the viral strain used in this experiment had 99.78% nucleotide homology with GCRV-873 (GenBank accession number JN206664.1). In the phylogenetic tree constructed based on the VP4 sequence, GCRV-Amphi2025 clustered with GCRV-873 (bootstrap value = 98%, Figure 1B), further confirming it as a typeIGCRV.
3.2. GCRV Infection Caused Structural Damage of Amphioxus Gill
Currently, no virus suitable for amphioxus immunostimulation has been identified. Here, we tested two fish-targeted viruses—double-stranded RNA virus GCRV (Grass Carp Reovirus) and single-stranded RNA virus SVCV (Spring Viraemia of Carp Virus)—to stimulate amphioxus. After preliminary experiments, we found that only GCRV appeared to cause damage to amphioxus. Therefore, we conducted a detailed observation of the immune response of amphioxus to GCRV. Amphioxus were soaked in viral suspension, and gross morphological observations revealed distinct phenotypic differences: the body region (corresponding to the gill area, marked by the blue rectangle in Figure 2A) of GCRV-treated amphioxus showed obvious redness and swelling, while no such abnormality was observed in the control group (marked by the red rectangle in Figure 2A). Further histological examination (Figure 2B) demonstrated that diffuse detachment of gill filament epithelial cells, significant mesenchymal congestion, and disorganized gill tissue structure were observed in amphioxus following GCRV treatment, while gill filament epithelial cells in the control group were neatly arranged without mesenchymal congestion. Statistical analysis of damaged areas in gill tissues (Figure 2C) showed that the proportion of damaged gill tissue in the virus-stimulated group was significantly higher than that in the control group.
3.3. The GCRV Survived in Seawater for No More than 48 h
To determine the survival duration of GCRV in seawater, we added a 10^5^ TCID_50_/mL virus suspension to sterile seawater at room temperature (25 °C) and collected samples at 0, 2, 6, 12, 24, 48 and 72 h to detect infectivity on EPC cells. Crystal violet assay showed that the infectivity of GCRV on EPC cells after 48 h in seawater was indistinguishable from the virus-free control group (Figure 3), indicating that the virus survival time in seawater was ≤48 h.
3.4. Infection Kinetics of GCRV on Amphioxus
To detect the dynamic changes of GCRV in amphioxus, amphioxus were soaked in seawater containing GCRV for 2 h, then transferred to normal seawater without GCRV. During this period, gill tissues were sampled at regular intervals to detect the expression of GCRV capsid protein VP5 mRNA. Real-time fluorescent quantitative PCR results showed that the expression level of VP5 mRNA increased rapidly after amphioxus were soaked in seawater containing GCRV for 2 h, and then decreased rapidly after transfer to normal seawater without GCRV. However, it significantly increased again at 14 h, followed by a gradual decline. Although the expression was low at 48 h, it was still detectable (Figure 4A). Tissue distribution analysis showed that the expression of VP5 mRNA in gill tissues at 14 h was significantly higher than that in intestinal and hepatic caecum tissues (Figure 4B), indicating that the gills might be the main target organ for GCRV infection.
3.5. GCRV Can Be Transmitted Among Amphioxus Through Co-Culture
We investigated the possibility of inter-individual transmission of GCRV when infected amphioxus (Group A) were co-cultured with healthy ones (Group B). After 12 h of GCRV infection of amphioxus in Group A, they were mixed with those in Group B (Figure 5). Subsequently, gill tissues from both groups were sampled at 12, 24, 48, 72 and 108 h post-mixing to detect the expression of VP5 mRNA. After 24 h of co-culture, VP5 mRNA expression was detectable in water, but not in group B amphioxus; by 48 h of co-culture, VP5 mRNA expression in water increased, while VP5 mRNA expression in group A amphioxus decreased to a level comparable to that in water, and VP5 mRNA expression became detectable in group B amphioxus at this point. By 72 h of co-culture, VP5 mRNA expression in group B amphioxus continued to rise and was higher than that in group A amphioxus (Figure 5). By 108 h of co-culture, VP5 mRNA expression had decreased in both groups of amphioxus and in water. These data indicate that GCRV can be transmitted among amphioxus through co-culture.
Additionally, tissue sections showed obvious gill damage in both group A and group B amphioxus after 72 h of co-culture (Figure 6A); HE staining (Figure 6A) showed both groups had gill lesions, with Group A exhibiting more severe epithelial detachment and interstitial hemorrhage than Group B. Quantitative analysis (Figure 6B) revealed the damaged area was 42.6 ± 1.9% in Group A, significantly higher than 28.5 ± 0.9% in Group B (Student’s t–test, *** p < 0.001).These results confirm that GCRV can be indirectly transmitted from one amphioxus to another through water.
3.6. Transcriptome Analysis Reveals Significant Activation of Immune-Related Pathways
We further analyzed the effect of GCRV stimulation on gene expression in amphioxus through transcriptome sequencing. After treating amphioxus with GCRV for 14 h, gill tissues were collected for RNA-seq analysis. According to the screening criteria of |log_2_FC| ≥ 2 and padj < 0.05, a total of 523 differentially expressed genes (DEGs) were identified, of which 317 genes were significantly upregulated.
GO functional enrichment analysis showed that the DEGs were significantly enriched in biological processes such as aminoglycan metabolic process, chitin metabolic process and chitin binding, as well as molecular functions such as peptidase activity (Figure S1). KEGG pathway analysis indicated that after GCRV stimulation, the DEGs were significantly enriched in the MAPK signaling pathway, calcium signaling pathway and ECM-receptor interaction pathways (Figure 7). We validated some of the differentially expressed immunity-related genes by qPCR, and the expression of caspase-3a, caspase-3b, camk1da and camk1ga genes were all significantly upregulated, mapk9 and mapk10 genes were significantly down-regulated (Figure 8), consistent with the RNA-seq data.
4. Discussion
This study is the first to confirm that grass carp reovirus (GCRV)—a virus originally identified in fish—can infect the cephalochordate amphioxus. Triple PCR detection and VP4 gene sequencing results indicated that the viral strain infecting amphioxus belongs to GCRV genotype I, with its VP4 gene sharing 99.78% homology with that of the reference strain GCRV-873.
The high conservation of the VP4 protein is hypothesized to serve as a key molecular basis for GCRV’s cross-species infection capability [3]. In fish hosts, the VP4 protein of GCRV-I mediates viral adsorption by recognizing host cell surface receptors, specifically integrin αVβ3 [12] or mucopolysaccharides [13]. Our transcriptome sequencing analysis revealed significant enrichment of chitin-binding protein-encoding genes and amino sugar metabolic pathways in the gill tissues of amphioxus following GCRV stimulation. This observation suggests that GCRV may utilize chitin derivatives on the surface of amphioxus gill epithelial cells as receptors, thereby breaking through species barriers via conserved glycoprotein-mediated interactions [14]. Notably, this cross-species infection mechanism resembles that of the newly isolated HGCRV (healthy grass carp reovirus), which exhibits cross-fish-host transmission ability. This similarity highlights the need for further research to explore the universal significance of viral surface protein conservation in facilitating cross-species infection [15].
The virus survival curve in a seawater environment showed that the infectious titer dropped below the detection limit after 48 h, which differs from the survival characteristics of genotype I GCRV in freshwater environments [16]. This may be attributed to the effect of salt ions on the stability of the viral capsid [17]. Waterborne exposure experiments confirmed that the virus can be transmitted between amphioxus individuals through water [18]. Waterborne transmission may lead to the rapid spread of diseases in farmed populations [14]. However, daily water changes (≥50%) can effectively reduce the viral load in water and disrupt the transmission chain within 48 h post-infection [17].
Infection kinetics showed that VP5 mRNA in gill tissues peaked at 14 h, 3.2-fold higher than in intestinal tissues, which is consistent with the gill tropism observed in grass carp infected with genotype I and II GCRV [19]. The peak of VP5 mRNA at 14 h post-infection (hpi) results from the dynamic balance between GCRV replication and the host innate immune response: GCRV-I strains have a 12–16 h replication cycle, with capsid protein synthesis (including VP5) peaking in the late phase (8–16 hpi) to support virion assembly, which provides the viral replication basis for the 14 hpi peak [20]; meanwhile, amphioxus initiates innate immune responses (e.g., ERK2-mediated MAPK pathway activation) around 12 hpi upon viral stimulation, but the translation and functional activation of antiviral effectors require a 2–4 h delay, so the immune inhibitory effect has not yet been fully exerted at 14 hpi, allowing sustained high-level VP5 transcription—a time window shaped by both viral replication kinetics and the primitive characteristics of the amphioxus innate immune response (slightly delayed activation compared to vertebrates) [21]. HE staining of histopathological sections revealed severe damage to gill filament tissues in the infected group, accompanied by activation of the MAPK signaling pathway and the calcium signaling pathway. Since amphioxus lack T/B cells, their immune responses rely on innate pathways activated by pattern recognition receptors (such as chitin receptors) [22]. The enrichment of the ECM-receptor interaction pathway in this study may reflect attempts by gill epithelial cells to resist viral infection through extracellular matrix remodeling. However, excessive activation of the MAPK pathway may mediate apoptosis via caspase-3, leading to an imbalance between “immune defense” and “tissue damage” [6]. This response pattern is highly similar to the early inflammatory response in vertebrates [8].
This study also has limitations: first, it only uses a laboratory infection model and does not verify the viral carriage rate in wild amphioxus populations; second, it does not clarify the specific interaction mechanisms between viral and amphioxus. Future research could use yeast two-hybrid technology to screen VP4- or VP5-binding proteins, and carry out epidemiological investigations on wild amphioxus populations. In addition, single-cell transcriptome analysis can identify specific target cell types (such as ciliated cells or basal cells) in gill tissues infected by the virus, further clarifying the cell-specific mechanisms of interactions between cephalochordates and viruses.
In conclusion, this study is the first to confirm that grass carp reovirus (GCRV) can infect amphioxus and possesses the ability for waterborne transmission. These findings not only expand the known host range of GCRV but also establish a novel model for investigating the cross-species transmission mechanisms of aquatic viruses and the evolutionary trajectory of vertebrate innate immunity.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Li M. Xue M. Zhou Y. Liu W. Meng Y. Xu C. Li Y. Jiang N. Fan Y. The Antiviral Efficacy of Andrographolide against Grass Carp Reovirus in Vitro and in Vivo Fish. Shellfish. Immunol.202516311039010.1016/j.fsi.2025.11039040354886 · doi ↗ · pubmed ↗
- 2Rao Y. Su J. Insights into the Antiviral Immunity against Grass Carp (Ctenopharyngodon Idella) Reovirus (GCRV) in Grass Carp J. Immunol. Res.2015201567043710.1155/2015/67043725759845 PMC 4337036 · doi ↗ · pubmed ↗
- 3Yan X. Wang Y. Xiong L. Jian J. Wu Z. Phylogenetic Analysis of Newly Isolated Grass Carp Reovirus Springer Plus 2014319010.1186/2193-1801-3-19024834372 PMC 4021032 · doi ↗ · pubmed ↗
- 4Jiang L. Liu A.-Q. Zhang C. Zhang Y.-A. Tu J. Hsp 90 Regulates GCRV-II Proliferation by Interacting with VP 35 as Its Receptor and Chaperone J. Virol.202296 e 011752210.1128/jvi.01175-2236102647 PMC 9555151 · doi ↗ · pubmed ↗
- 5Gao Y. Huo X. Wang Z. Yuan G. Liu X. Ai T. Su J. Oral Administration of Bacillus Subtilis Subunit Vaccine Significantly Enhances the Immune Protection of Grass Carp against GCRV-II Infection Viruses 2021143010.3390/v 1401003035062234 PMC 8779733 · doi ↗ · pubmed ↗
- 6Yang Z. Zhang H. Yuan Z. Chen J. Zheng G. Zou S. The Effects of GCRV on Various Tissues of Grass Carp (Ctenopharyngodon Idella) and Identification of Differential Interferon-Stimulating Genes (IS Gs) through Muscle Transcriptome Analysis Ecotoxicol. Environ. Saf.202428411695610.1016/j.ecoenv.2024.11695639208574 · doi ↗ · pubmed ↗
- 7Feng Y. Shi C. Ouyang P. Huang X. Chen D. Wang Q. Yin J. Zhang D. Geng Y. The National Surveillance Study of Grass Carp Reovirus in China Reveals the Spatial-Temporal Characteristics and Potential Risks Aquaculture 202254773744910.1016/j.aquaculture.2021.737449 · doi ↗
- 8D’Aniello S. Bertrand S. Escriva H. Amphioxus as a Model to Study the Evolution of Development in Chordates Elife 202312 e 8702810.7554/e Life.8702837721204 PMC 10506793 · doi ↗ · pubmed ↗