Duck Plague Virus Full-Length UL15 Protein Is a Multifunctional Enzyme Which Not Only Possesses Nuclease Activity but Also Exerts ATPase and DNA-Binding Activity
Qiao Yang, Guoying Zhou, Jing Yang, Mingshu Wang, Ying Wu, Bin Tian, Anchun Cheng

TL;DR
This study shows that the DPV pUL15 protein has three key functions: DNA binding, ATP hydrolysis, and DNA cleavage, making it essential for viral DNA packaging.
Contribution
The study identifies the full-length DPV pUL15 as a multifunctional enzyme with independent ATPase, DNA-binding, and nuclease activities.
Findings
Full-length pUL15 exhibits non-sequence-specific nuclease activity.
The N-terminal region of pUL15 is responsible for DNA-binding and ATPase functions.
The C-terminal region of pUL15 is responsible for nuclease activity, independent of the other functions.
Abstract
Duck plague virus (DPV), an alphaherpesvirus, utilizes a terminase complex for genome packaging. This study focuses on the large terminase subunit, pUL15, and characterizes its functional domains. The results showed that the full-length pUL15 protein exhibits three key enzymatic activities: it binds and cleaves DNA non-specifically, and hydrolyzes ATP. Further analysis using truncated mutants mapped the DNA-binding and ATPase functions to the N-terminal region, whereas the nuclease activity was localized to the C-terminus. Notably, these functions are independent, as the loss of nuclease activity did not impair the other two. These findings confirm DPV pUL15 as a multifunctional enzyme essential for genome processing and provide a fundamental basis for understanding the mechanism of viral DNA packaging in herpesviruses. The genome of the herpesvirus is a linear double-stranded DNA. The…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
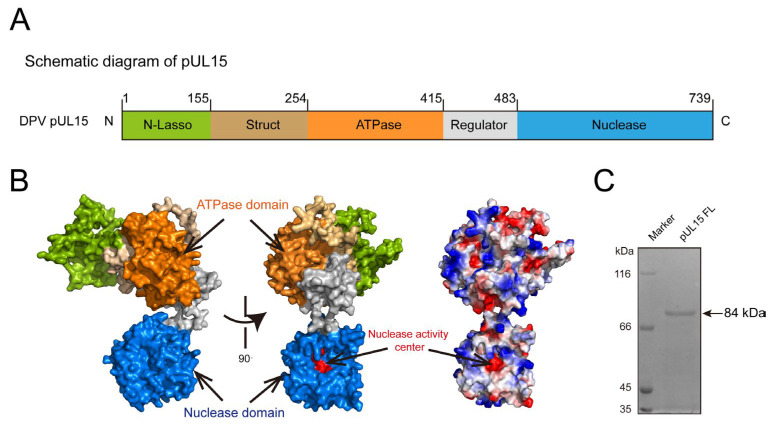 Figure 1
Figure 1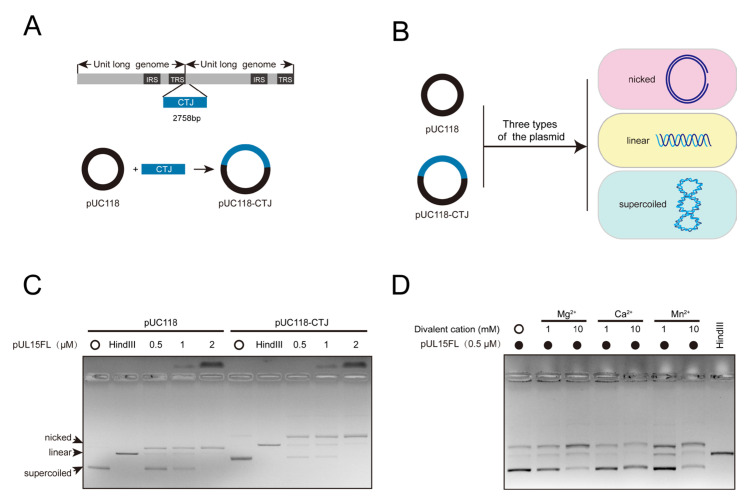 Figure 2
Figure 2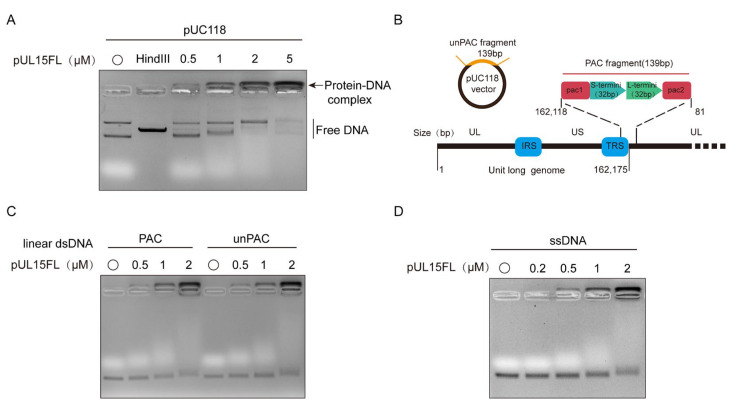 Figure 3
Figure 3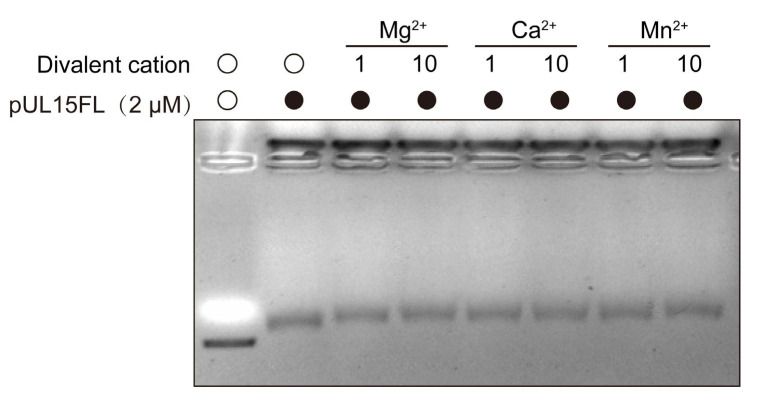 Figure 4
Figure 4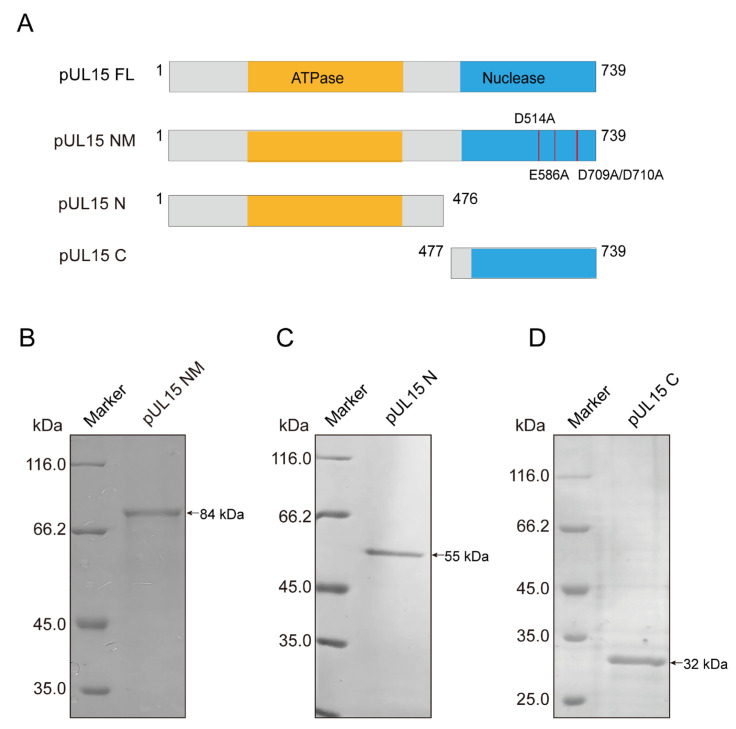 Figure 5
Figure 5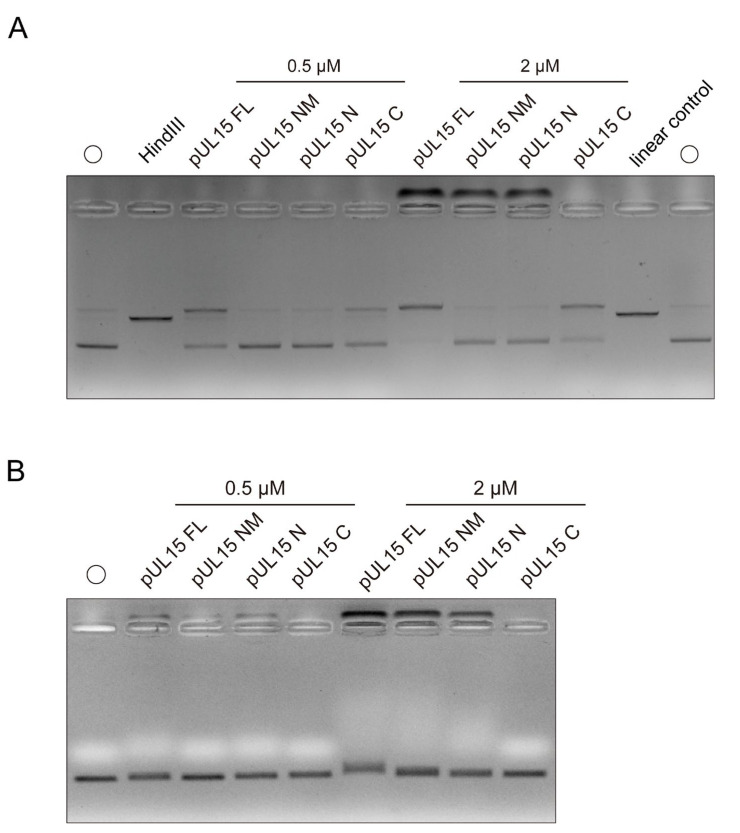 Figure 6
Figure 6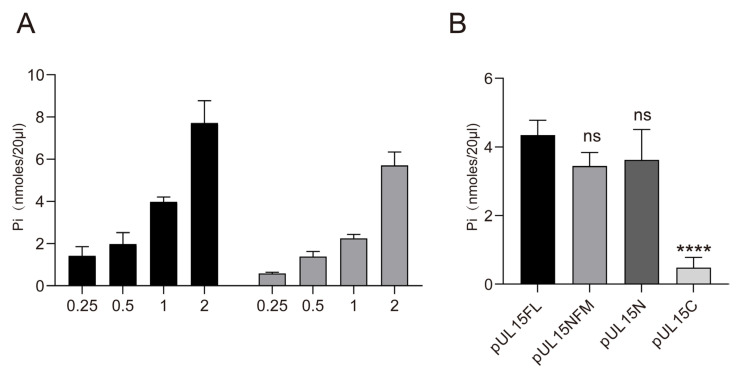 Figure 7
Figure 7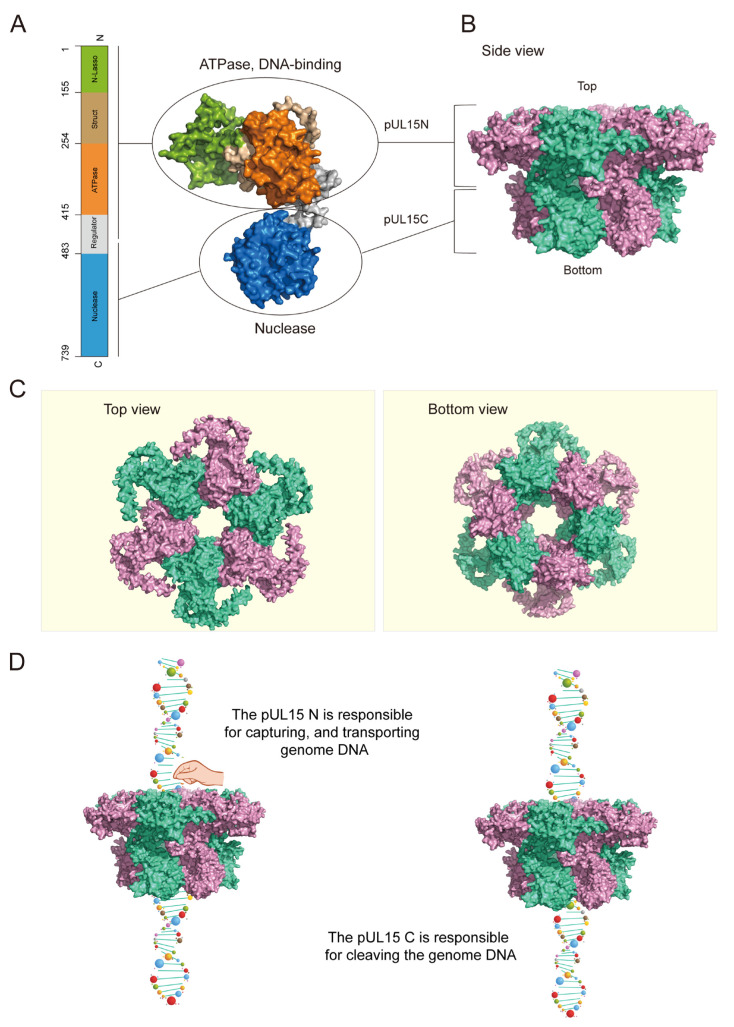 Figure 8
Figure 8- —China Agriculture Research System
- —Sichuan Veterinary Medicine, Drug Innovation Group of the China Agricultural Research System
- —Research Interest Training Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHerpesvirus Infections and Treatments · Mosquito-borne diseases and control · Toxoplasma gondii Research Studies
1. Introduction
Most large double-stranded DNA (dsDNA) viruses package their genomes into preformed capsids, as observed in herpesviruses and bacteriophages [1,2,3,4,5,6]. During replication, newly synthesized viral DNA forms head-to-tail concatemeric genomes, which must be cleaved into unit-length genomes and packaged into preassembled procapsids to generate mature virions [7,8]. The viral terminase plays a crucial role in this process, facilitating both the cleavage of the concatemeric genome and its translocation into the procapsid [9,10]. In herpesviruses, the terminase complex typically consists of three subunits: the large subunit pUL15, the small subunit pUL28, and the third subunit pUL33, or their respective homologs [11,12,13,14]. Following assembly in the host cytoplasm, the terminase complex translocates into the host nucleus to perform its function [15]. The large subunit of the terminase complex, pUL15, and its homologs contain two enzymatically active domains: an N-terminus ATPase domain and a C-terminal nuclease domain [16,17]. When pUL15 is null, the cleavage of the viral genome is inhibited and the mature capsid containing DNA cannot be produced; thus, the production of progeny virions is affected [18].
During herpesvirus packaging, the terminase complex docks onto the unique portal vertex, which consists of dodecameric pUL6 in HSV-1. Dodecameric pUL6 forms a circular channel, facilitating the entry of DNA into the preformed capsid in an ATP-dependent manner [19,20,21]. Once the capsid is filled with a unit-length genome, the large terminase subunit executes a second cleavage event, completing the genome packaging process [22]. In addition to the terminase complex and portal protein, several other essential proteins, including UL17, UL25, and UL32, are required for the concatemeric genome cleavage and packaging. Loss of function in any of these proteins results in packaging failure and the accumulation of immature capsids within the nucleus [23,24,25,26].
Despite the low sequence similarity, pUL15 and its homologs are structurally conserved across herpesviruses, underscoring their critical role in genome packaging. Sequence alignment and structure analyses indicate that pUL15 and its homologs belong to the Additional Strand Catalytic E (ASCE) superfamily, with highly conserved ATPase regions containing canonical Walker A and Walker B motifs [27,28,29]. Moreover, the confirmed nuclease-active sites of HSV-1 pUL15, amino acids D509, E581, D706, and D707, are highly conserved among herpesviruses [30]. To date, most studies on herpesvirus pUL15 and its homologs have focused on their nuclease activity. In vitro assays have demonstrated that HSV-1 pUL15 cleaves DNA nonspecifically, as observed in human cytomegalovirus (HCMV) and duck plague virus (DPV) [30,31,32,33]. While some reports have confirmed that pUL28 or its homologs can bind to DNA [34,35,36,37,38], the DNA-binding capability of pUL15 or its homologs remains largely unexplored, leaving uncertainty about their specificity in DNA binding.
The replication of DPV, also named Anatid herpesvirus-1, necessitates the terminase-mediated cleavage of the concatemeric genome to produce mature virions [39]. Sequence alignment reveals that DPV pUL15 is homologous to the large terminase subunits in HSV-1, HCMV, Epstein–Barr virus (EBV), and other herpesviruses, sharing highly conserved nuclease activity sites and Walker motifs. Previous research has demonstrated that the C-terminus domain of DPV pUL15 only possesses non-sequence-specific nuclease activity but lacks DNA-binding ability [33]. This study further explored the function of DPV full-length pUL15, revealing that it is a multifunctional enzyme possessing nuclease, ATPase, and DNA-binding activities. It was demonstrated that the N-terminus domain of pUL15 is responsible for ATPase and DNA-binding activities, while the C-terminus domain plays a key role in DNA cleavage.
2. Materials and Methods
2.1. Cloning, Mutation, and Protein Purification
The DPV full-length UL15 gene sequence (Gene ID: 8223370) was codon-optimized and inserted into the pET28a vector, and then transformed into E. coli BL21 for expression. UL15 N-terminus sequence (1 bp to 1428 bp) was amplified using the primers listed in Table A1, and the purified PCR product was cloned into the pET28a vector. A previously constructed plasmid, pET28a-UL15C NM, harboring mutations in the nuclease functional sites (D514A/E586A/D709A/D710A) of the pUL15C-terminal domain, was used as a template to generate a full-length nuclease functional site mutant plasmid (pET28a-UL15NM) via fusion PCR, and then transformed into E. coli BL21. Protein expression was performed as described previously [33]. Induced cultures were lysed by sonication, followed by centrifugation to remove debris. The supernatant was incubated with Ni-NTA agarose beads (Smart-Lifesciences, Changzhou, China) at room temperature for 30 min. The beads were washed with wash buffer (50 mM Tris–HCl, 0.5% Triton, 1 M NaCl, and 40–100 mM imidazole, adjusted for different mutant proteins) to remove impurities, and the target proteins were eluted using elution buffer (50 mM Tris–HCl, 250 mM NaCl, and 300 mM imidazole). Protein concentration was measured using a BCA assay kit (Solarbio, Beijing, China) and stored at −80 °C.
2.2. Nuclease Activity Assay
The nuclease activity assay was carried out according to the method previously described with some modifications [33]. Purified full-length or mutant pUL15 was incubated with 500 ng of circular pUC118 plasmid or pUC118-CTJ plasmid in nuclease buffer (10 mM Tris–HCl [pH 7.5], 10 mM MgCl_2_·6H_2_O, 1 mM DTT, 50 mM NaCl) in a 20 µL reaction system at 37 °C for 20 min. Hind III (Takara, Beijing, China) digestion of plasmid DNA under identical conditions served as a control to obtain linear plasmid bands. To evaluate the effect of metal ions on the nuclease activity of pUL15FL, 0.5 µM protein was incubated with pUC118 plasmid in nuclease buffer containing 1 mM or 10 mM MgCl_2_, MnCl_2_, or CaCl_2_. Reactions were terminated by adding 6× DNA loading buffer (Beyotime, Shanghai, China) and analyzed using 0.7% agarose gel electrophoresis, followed by ethidium bromide staining.
2.3. DNA Binding Assay
The key cis-acting signal for herpesvirus genome packaging is the “a” sequence, typically containing the conserved motifs Pac1 and Pac2 necessary for DNA cleavage and packaging. A 139 bp dsDNA fragment containing the DPV genome Pac motifs (PAC) or a 139 bp nonspecific sequence (unPAC) was used as DNA-binding substrate. The two fragments were amplified by PCR using the primers shown in Table A1, and the sequences of the two fragments are shown in Table A2. The DNA-binding reaction buffer contained 100 mM Tris–HCl, 0.5 mM Na_2_EDTA, 1 M KCl, 10 mM MgCl_2_, 10 mM DTT, and 25% glycerol. The purified pUL15 or its mutants were incubated with dsDNA Pac and unPac fragments in 20 µL DNA-binding reaction systems at 37 °C for 20 min. Single-stranded DNA (ssDNA) substrates (Invitrogen, Carlsbad, CA, USA) were substituted for dsDNA under similar conditions to evaluate the ssDNA-binding of pUL15FL. The reactions were terminated by adding 6× DNA loading buffer and analyzed on 1% agarose gels with ethidium bromide staining.
2.4. ATPase Activity Assay
ATPase activity was measured at 37 °C in reaction buffer containing 1 mM ATP, 40 mM Tris–HCl (pH 7.5), 50 mM NaCl, 2 mM MgCl_2_·6H_2_O, and indicated concentrations of full-length or mutant pUL15. ATP hydrolysis was detected using the Malachite Green Phosphate Detection Kit (Beyotime, Shanghai, China). Pi produced by ATP hydrolysis reacted with malachite green and molybdate to form a green phosphomolybdate complex, which could be detected at 630 nm. To evaluate the hydrolysis of different adenosine nucleotides, 1 mM ATP was replaced with 1 mM ADP as the substrate.
2.5. Structure Prediction and Analysis of DPV pUL15
The AlphaFold3 server was utilized (https://alphafoldserver.com/ (accessed on 11 October 2025)) to predict the structure of the monomer and hexamer of DPV pUL15. The sequences submitted to the server corresponded to the Genebank (UL15: YP_003084405.1). The predicted structural models (in PDB format) generated by AlphaFold3 were downloaded for further analysis. All structural visualization, analysis, and figure generation were performed using PyMOL 3.0.3.
3. Results
3.1. DPV Full-Length pUL15 Exhibits Non-Specific Nuclease Activity, Enhanced by Metal Ions
A previous study demonstrated that the DPV pUL15C-terminus domain possesses a negatively charged nuclease activity center and exhibits non-specific nuclease activity similar to its homologs in other herpesviruses [33]. Sequence alignment between DPV and HSV-1 pUL15 revealed that DPV full-length pUL15 (pUL15FL) not only contains the nuclease domain, but also harbors the ATPase domain and other conserved domains (Figure 1A,B). To determine whether pUL15FL exhibits specific nuclease activity, the UL15 gene was cloned into the pET28a vector, and then the recombinant His-tagged pUL15 (84 kDa) was expressed and purified (Figure 1C).
The plasmids pUC118 and pUC118-CTJ, with the latter containing a 2758 bp of concatemeric terminus junction (CTJ), were used as the substrates to assess nuclease activity (Figure 2A). Plasmids commonly contain three types: nicked (open circle), linear, and supercoiled (closed circle) (Figure 2B). Their migration rates in gel electrophoresis from slow to fast are open circle, linear, and supercoiled. The results indicated that the amount of both supercoiled plasmids decreased with increasing protein concentration, but the nicked and linear plasmids increased compared with the control, confirming that pUL15FL still non-specifically cuts DNA even in its full-length state. Interestingly, as the protein concentration increased, the DNA retention near the loading well also increased, suggesting that pUL15FL possesses a DNA-binding ability (Figure 2C).
To investigate the influence of metal ions on the nuclease activity of DPV pUL15FL, Mg^2+^, Ca^2+^, and Mn^2+^ were introduced into the reaction system. To minimize potential interference from DNA binding, a low concentration of pUL15FL was used. As expected, divalent cations enhanced the nuclease activity of pUL15FL, with Mg^2+^ and Mn^2+^ exhibiting stronger effects. Notably, no DNA retention was observed near the loading well under these conditions, indicating that metal ions may not affect the DNA binding ability of pUL15FL (Figure 2D).
3.2. DPV pUL15FL Exhibits Non-Specific DNA-Binding Ability
The above results demonstrated that DPV pUL15FL not only exerted nuclease activity, but also possessed non-specific DNA-binding ability in the nuclease reaction buffer. To determine whether this persisted in a DNA-binding reaction buffer, the pUC118 plasmid was used as substrate. As shown in Figure 3A, as the pUL15FL concentration increased, the free DNA decreased and the DNA remaining near the loading well increased, suggesting that pUL15FL could bind DNA, and the buffer conditions did not affect DNA-binding specificity.
To assess whether pUL15FL can bind to different forms of DNA, linear dsDNA and ssDNA were tested as substrates. The results showed that pUL15FL could bind non-specifically to both dsDNA and ssDNA, indicating that its DNA interaction is independent of a specific form and sequence (Figure 3C,D). Since divalent metal ions could promote the nuclease activity of pUL15, we also wanted to explore whether metal ions could also enhance the binding ability of pUL15FL to DNA. Here, 1 and 10 mM Mg^2+^, Ca^2+^, and Mn^2+^ were used for detection. It was found that, unlike nuclease activity, even in high concentrations, metal ions still could not enhance the DNA-binding ability of pUL15FL. The result showed the DNA-binding capacity of pUL15FL did not depend on metal ions (Figure 4). The above results indicated that pUL15FL could bind to non-specific DNA independently of metal ions.
3.3. The N-Terminus of pUL15 Is Responsible for Its Interaction with DNA
A previous study has shown that the C-terminal domain of DPV pUL15 exhibits nuclease activity but lacks DNA-binding ability. Combined with the findings of this study, we hypothesized that DNA-binding activity resides in the N-terminal domain. To test this, a plasmid expressing the N-terminus domain of pUL15 was constructed in this study, and the pUL15C, which expressed the C-terminus domain, was constructed in the previous study. Additionally, a nuclease-deficient mutant pUL15NM was generated to examine whether DNA binding depends on its nuclease activity (Figure 5A). Finally, the soluble pUL15NM (84 kDa), pUL15N (55 kDa), and pUL15C (32 kDa) were purified and detected through SDS-PAGE, respectively (Figure 5B–D).
As shown in Figure 6A, plasmid pUC118 was used as the substrate to detect the nuclease activity in different mutants. Whether under conditions of low concentrations or high concentrations of pUL15FL and its mutants, it was obvious that the nicked plasmid increased but the supercoiled plasmid decreased when only pUL15FL or pUL15C was present in the reaction, compared to the control. This indicated the plasmid was cleaved by pUL15FL and pUL15C, but not pUL15NM and pUL15N (Figure 6A). When the concentration of protein was increased to 2 µM, DNA binding was easier to detect. It was obvious that plasmid DNA accumulated near the loading wells when pUL15FL, pUL15NM, and pUL15N were present in the reaction (Figure 6A). The formation of protein–DNA complex indicated that, in addition to the pUL15 C, the pUL15FL, pUL15NM, and pUL15N all possessed the ability to bind DNA. These results suggested that the N-terminus domain of pUL15 exerted DNA-binding activity, and the loss of nuclease activity did not affect the binding of pUL15 to DNA.
To further rule out the influence of nuclease activity on the observation of pUL15’s ability to bind to DNA, the substrate was replaced by the linear dsDNA, PAC. In Figure 6B, pUL15FL and its mutants, except pUL15C, exhibited obvious DNA-binding activity at high protein concentrations. These findings indicated that the N-terminus region of DPV pUL15 was only capable of DNA binding capacity, while the C-terminus region could only exert nuclease activity. In addition, the loss of nuclease activity of the full-length pUL15 did not affect its DNA-binding capacity.
3.4. The N-Terminus of DPV pUL15 Not Only Exerted the Ability to Bind DNA, but Also Possessed ATPase Activity
The ATPase activity of the terminase large subunit has been well characterized in bacteriophages, including SPP1, Sf6, and Phi29 [5,40,41]. In HSV-1, a previous study demonstrated that the pUL15 contains the ATPase domains and hydrolyzes ATP. However, many aspects of pUL15 ATPase function remain unclear. In this study, we found that DPV pUL15FL hydrolyzes both ATP and ADP, with ATP yielding a higher release of phosphate groups than ADP (Figure 7A). Based on the analysis of sequence and structure, it was known that the ATPase region is located at the N-terminus. However, we still did not know whether ATPase activity could be exerted completely without the presence of the C-terminus. Therefore, we detected the ATPase activity of different mutants of DPV pUL15. As shown in Figure 7B, the pUL15N exhibited ATPase activity like pUL15FL, whereas pUL15C did not. Notably, similar to its DNA-binding ability, pUL15NM retained ATPase activity even in the absence of its nuclease function. These findings suggest that the DNA-binding and ATP-hydrolysis activities of pUL15 are independent of its nuclease domain, with the N-terminus playing a crucial role in both functions.
3.5. Structure Prediction and Functional Model of DPV pUL15
The above results showed that DPV pUL15 was a multifunctional protein, which simultaneously possessed ATPase, DNA-binding, and nuclease activity. In this study, it was demonstrated that the N-terminus of DPV pUL15 could bind to DNA and could hydrolyze ATP, and the C-terminus of DPV pUL15 possessed the nuclease activity. The structure of the monomer and hexamer of DPV pUL15 was predicted by AlphaFold 3. The prediction results showed that the pUL15 was in an inverted L shape (Figure 8A). The N-terminus domain with DNA binding ability and ATPase activity was located in the upper part of the inverted L shape, while the C-terminus domain with nuclease activity was located in the lower part. The N-terminus domain and the C-terminus domain were connected by a region defined as a regulator (Figure 8A). Based on the composition of the terminase in HSV-1, we predicted the structure of pUL15 hexamer in DPV. The results showed that the DPV pUL15 hexamer presented an inverted cap structure with a channel in the middle (Figure 8B,C). The channel in the middle of the hexamer could be used to bind and transport DNA. According to the confirmed functions and structures of DPV pUL15, we proposed a hypothetical functional model of DPV pUL15 (Figure 8D). During the process of genome package and cleavage in DPV, pUL15 forms a hexamer, and the genome DNA binds to the channel in the hexamer. The pUL15 provides energy for the translocation of DNA in the channel by hydrolyzing ATP, thus allowing the genome to enter the capsid. When the complete genome enters the capsid, the N-terminus of DPV pUL15 releases the genomic DNA, and the C-terminus cleaves the genomic DNA through its nuclease activity, causing the remaining tandem genome to be detached from the capsid.
4. Discussion
Herpesviruses possess a linear double-stranded DNA (dsDNA) genome enclosed within an icosahedral capsid [42]. The newly synthesized viral genome exists in host cells as a head-to-tail concatemeric DNA, which must be cleaved into unit-length genomes before packaging into the capsid. This process is mediated by a virus-encoded terminase complex [43,44]. In HSV-1, pUL15, pUL28, and pUL33 form a complex in host cells, which is transported into the nucleus via the nuclear localization signal in pUL15 [11]. The large subunit pUL15 comprises two structural domains: a C-terminal nuclease domain, structurally similar to RNase H, and an N-terminal ATPase domain belonging to the ASCE superfamily [45,46,47]. pUL15 plays a crucial role in cleaving concatemeric genomes and hydrolyzing ATP to generate the energy required for genome translocation [41]. DPV, a member of the herpesvirus family, encodes homologs of HSV-1 terminase subunits, including pUL15, pUL28, and pUL33. It has been shown that DPV pUL15 exhibits DNA cleavage activity, but the nuclease activity is non-sequence-specific in vitro [33]. Similar non-sequence-specific nuclease activity has been reported for the large subunits of terminases in other herpesviruses and bacteriophages [5,30,32,48]. However, herpesvirus concatemeric DNA cleavage in vivo is highly specific. Furthermore, previous studies on the nuclease activity of pUL15 have primarily focused on its C-terminal region. Therefore, the present study aimed to investigate whether full-length DPV pUL15 specifically cleaves DNA.
In this study, the sequence of DPV UL15 was optimized and ligated into the prokaryotic expression vector pET28a, and the soluble full-length UL15 protein was successfully purified. It was found that full-length DPV pUL15 functions as a metal-ion-dependent nuclease; its nuclease activity was significantly enhanced in the presence of Mn^2+^ or Mg^2+^. It was also found that the full-length DPV pUL15 can still cleave different DNA substrates, indicating that DPV pUL15 exhibits non-sequence-specific nuclease activity, which is similar to that of HSV-1 pUL15 and HCMV pUL89 [16,49]. Notably, unlike the truncated pUL15C, full-length pUL15 exhibited significantly enhanced DNA-binding ability at higher protein concentrations, a characteristic commonly observed in bacteriophage terminase large subunits [50,51]. This finding was consistent with structural analyses of HSV-1 pUL15, but represented the first experimental confirmation of this phenomenon. These results suggested that the N-terminal domain of pUL15 was primarily responsible for DNA binding, while the C-terminal domain exerted nuclease activity. The loss of nuclease function did not significantly affect DNA binding.
As one of the most powerful molecular motors identified, the terminase large subunit hydrolyzes ATP to drive the cleavage and packaging of concatemeric viral DNA. The ATPase activity of pUL15 in herpesviruses was first detected in HSV-1 in recent years, yet many aspects of its function remain unclear. In this study, we demonstrated that the N-terminus of DPV pUL15 could significantly hydrolyze ATP just like the full-length pUL15. The above results suggest that the N-terminal ATPase domain had the potential to bind to DNA, whereas the nuclease domain is dispensable for DNA binding. This study demonstrated that DPV pUL15 is a multifunctional enzyme, which not only possesses nuclease and ATPase activity, but also has DNA-binding ability.
The structural analysis of the HSV-1 terminase complex showed that the complex can be assembled into monomers, hexamers, and decadimers, and the hexamer is the predominant assembly form. In this ring-like structure, six copies of the ATPase domain are arranged inside the ring, forming a “funnel” channel structure. Six basic residues are observed inside this channel. These basic residues are likely to be responsible for binding to DNA during translocation [16,45]. A mechanism has also been reported in bacteriophage P74-26, in which the N-terminal domain of its large subunit TerL is responsible for gripping DNA to enable cleavage by the C-terminal domain [52]. DPV pUL15 hexamer, which was predicted by AlphaFold 3, displayed a similar structure with a channel that might be responsible for DNA binding and translocation. It was hypothesized that different enzymatic activities of DPV pUL15 played a synergistic role in the process of viral genome cleavage and packaging, such as performing actions like “DNA binding”–“hydrolyzing ATP”–“DNA translocation”, etc. and repeating these cycles until the complete genomic DNA was packaged into the capsid, and then it exerted its nuclease activity to complete the genome packaging.
DPV pUL15, as a multifunctional enzyme with independent domains, has direct implications for understanding viral pathogenesis in waterfowl. As the terminase complex is essential for the production of infectious virions, our findings suggest that the nuclease and ATPase activities of pUL15 are attractive targets for antiviral intervention. From a practical veterinary perspective, the results provide a foundation for several future research avenues: (i) high-throughput screening for compounds that disrupt UL15’s nuclease or ATPase activities; and (ii) engineering live-attenuated vaccines with mutations in the functional domain that impair viral replication. Furthermore, given the high conservation of the terminase complex across herpesviruses, the insights gained from this study not only contribute to the control of duck plague, but could also inform the development of broad-spectrum anti-herpesviral drugs targeting the terminase complex. Herpesviruses (such as HCMV, KSHV, and HSV) can cause a variety of serious diseases. Antiviral drugs currently used in clinical practice, such as acyclovir and ganciclovir, mainly play a role by inhibiting viral DNA polymerase [53,54]. However, the application of these drugs is often limited by adverse reactions and drug resistance mutations [55]. The concatemeric DNA packaging process is highly specific to herpesviruses and has no counterpart in host cellular processes, making it an ideal target for developing novel antiviral agents. Small molecule inhibitors targeting the nuclease or ATPase activities of the UL15 protein exhibit unique mechanisms of action and potentially improved safety profiles, offering new strategies for the prevention and treatment of herpesvirus infections. Therefore, targeting the terminase complex represents a promising direction for future antiviral drug development [56]. In this context, elucidating the structure and mechanism of the terminase complex and its role in DNA packaging will be of critical importance.
5. Conclusions
In summary, this study demonstrated that DPV pUL15 was a multifunctional enzyme with ATPase, nuclease, and DNA-binding activities. In vitro, DPV pUL15 did not specifically recognize or cleave DNA, its DNA-binding and ATP hydrolysis abilities were dominated by the N-terminus, and nuclease activity was carried out by the C-terminus. During viral replication, it was speculated that the different functions of DPV pUL15 worked in synergy to complete the cleavage and packaging of the viral genome. Our study provided clues for the future exploration of DPV genome cleavage and packaging. Meanwhile, it provided a basis for comparative analysis of the terminase complex of different hosts.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baines J.D. Herpes simplex virus capsid assembly and DNA packaging: A present and future antiviral drug target Trends Microbiol.20111960661310.1016/j.tim.2011.09.00122000206 · doi ↗ · pubmed ↗
- 2Neuber S. Wagner K. Messerle M. Borst E.M. The C-terminal part of the human cytomegalovirus terminase subunit p UL 51 is central for terminase complex assembly J. Gen. Virol.20189911913410.1099/jgv.0.00098429205134 · doi ↗ · pubmed ↗
- 3Chiu S.H. Wu M.C. Wu C.C. Chen Y.C. Lin S.F. Hsu J.T. Yang C.S. Tsai C.H. Takada K. Chen M.R. Epstein-Barr virus BALF 3 has nuclease activity and mediates mature virion production during the lytic cycle J. Virol.2014884962497510.1128/JVI.00063-1424554665 PMC 3993834 · doi ↗ · pubmed ↗
- 4Smits C. Chechik M. Kovalevskiy O.V. Shevtsov M.B. Foster A.W. Alonso J.C. Antson A.A. Structural basis for the nuclease activity of a bacteriophage large terminase EMBO Rep.20091059259810.1038/embor.2009.5319444313 PMC 2685612 · doi ↗ · pubmed ↗
- 5Zhao H. Christensen T.E. Kamau Y.N. Tang L. Structures of the phage Sf 6 large terminase provide new insights into DNA translocation and cleavage Proc. Natl. Acad. Sci. USA 20131108075808010.1073/pnas.130113311023630261 PMC 3657791 · doi ↗ · pubmed ↗
- 6Mc Voy M.A. Adler S.P. Human cytomegalovirus DNA replicates after early circularization by concatemer formation, and inversion occurs within the concatemer J. Virol.1994681040105110.1128/jvi.68.2.1040-1051.19948289333 PMC 236542 · doi ↗ · pubmed ↗
- 7Heming J.D. Huffman J.B. Jones L.M. Homa F.L. Isolation and characterization of the herpes simplex virus 1 terminase complex J. Virol.20148822523610.1128/JVI.02632-1324155374 PMC 3911699 · doi ↗ · pubmed ↗
- 8Beard P.M. Duffy C. Baines J.D. Quantification of the DNA cleavage and packaging proteins U(L)15 and U(L)28 in A and B capsids of herpes simplex virus type 1J. Virol.2004781367137410.1128/JVI.78.3.1367-1374.200414722291 PMC 321391 · doi ↗ · pubmed ↗