Phytotoxic Effects and Microbial Responses to Ciprofloxacin and Its Removal by Hydrilla verticillata
Linzhi Lu, Rong Huang, Liang Wan, Guijia Li, Zhenhao Xu, Jiahao Guo

TL;DR
This study shows how the aquatic plant Hydrilla verticillata can remove the antibiotic ciprofloxacin from water while also suffering toxic effects and altering microbial communities.
Contribution
The study identifies specific biodegradation pathways and microbial shifts in response to ciprofloxacin using Hydrilla verticillata.
Findings
Hydrilla verticillata removed up to 37% of ciprofloxacin at low concentrations under high biomass conditions.
Ciprofloxacin exposure reduced plant growth, photosynthesis, and chlorophyll content while increasing oxidative stress.
Ciprofloxacin exposure altered microbial communities, reducing diversity and promoting antibiotic-resistant bacteria.
Abstract
Ciprofloxacin (CIP), a widely used fluoroquinolone antibiotic, is frequently detected in aquatic environments, raising concerns over its ecological risks. In this study, the submerged macrophyte Hydrilla verticillata was employed to investigate its capacity for CIP removal and the associated ecotoxicological effects. A series of batch experiments were conducted to evaluate plant growth, photosynthetic efficiency, oxidative stress responses, CIP biodegradation pathways, and shifts in epiphytic microbial communities. Results showed that CIP significantly inhibited the growth of H. verticillata, with inhibition rates of plant length and fresh weight reaching 15.8% and 29.7%, respectively, at 5 mg/L. Photosynthetic parameters were severely suppressed. Fv/Fm represented the maximum quantum efficiency of PSII, significantly decreased by 94.3% at 10 mg/L, while chlorophyll a and b contents…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
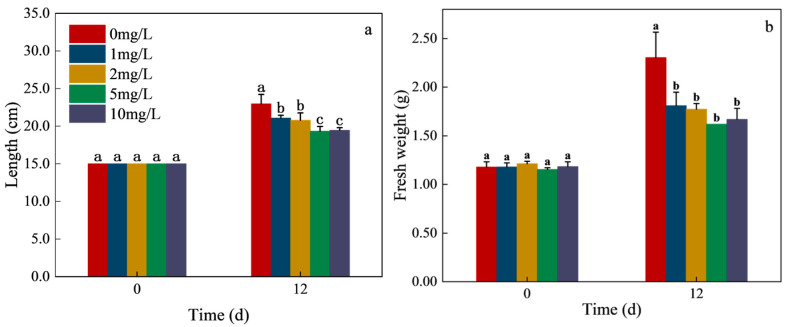 Figure 1
Figure 1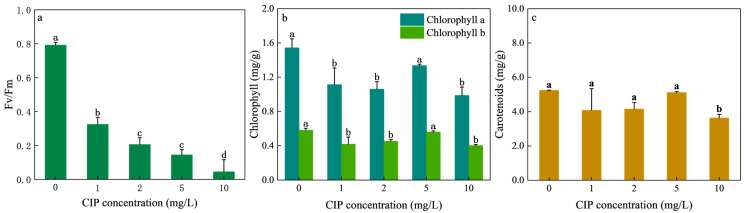 Figure 2
Figure 2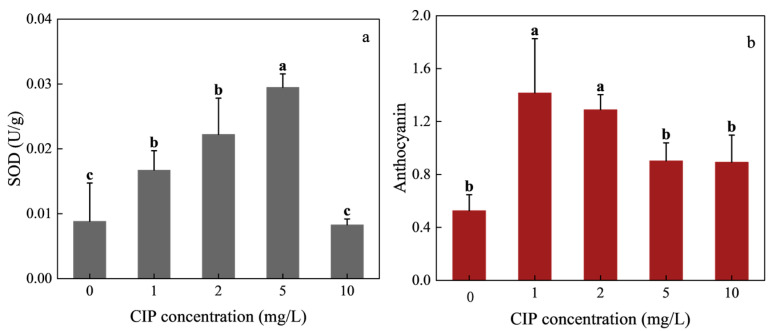 Figure 3
Figure 3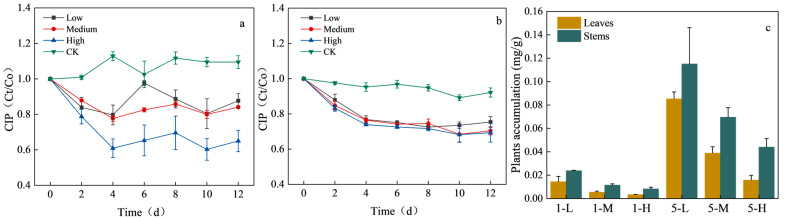 Figure 4
Figure 4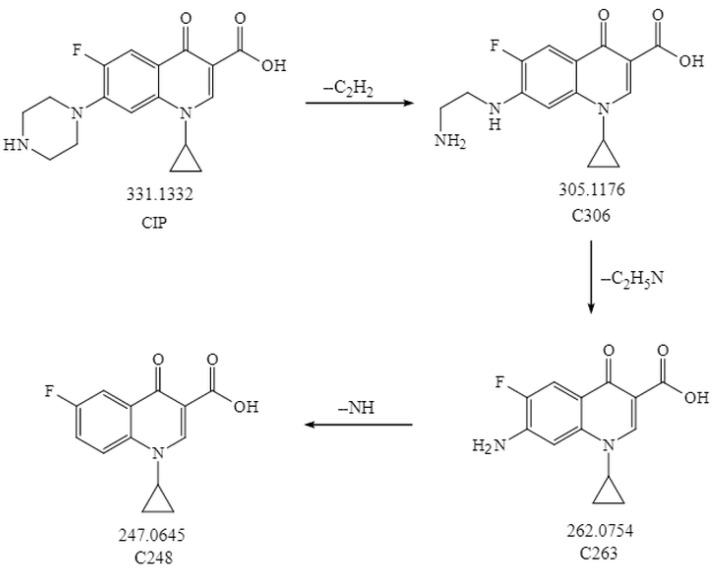 Figure 5
Figure 5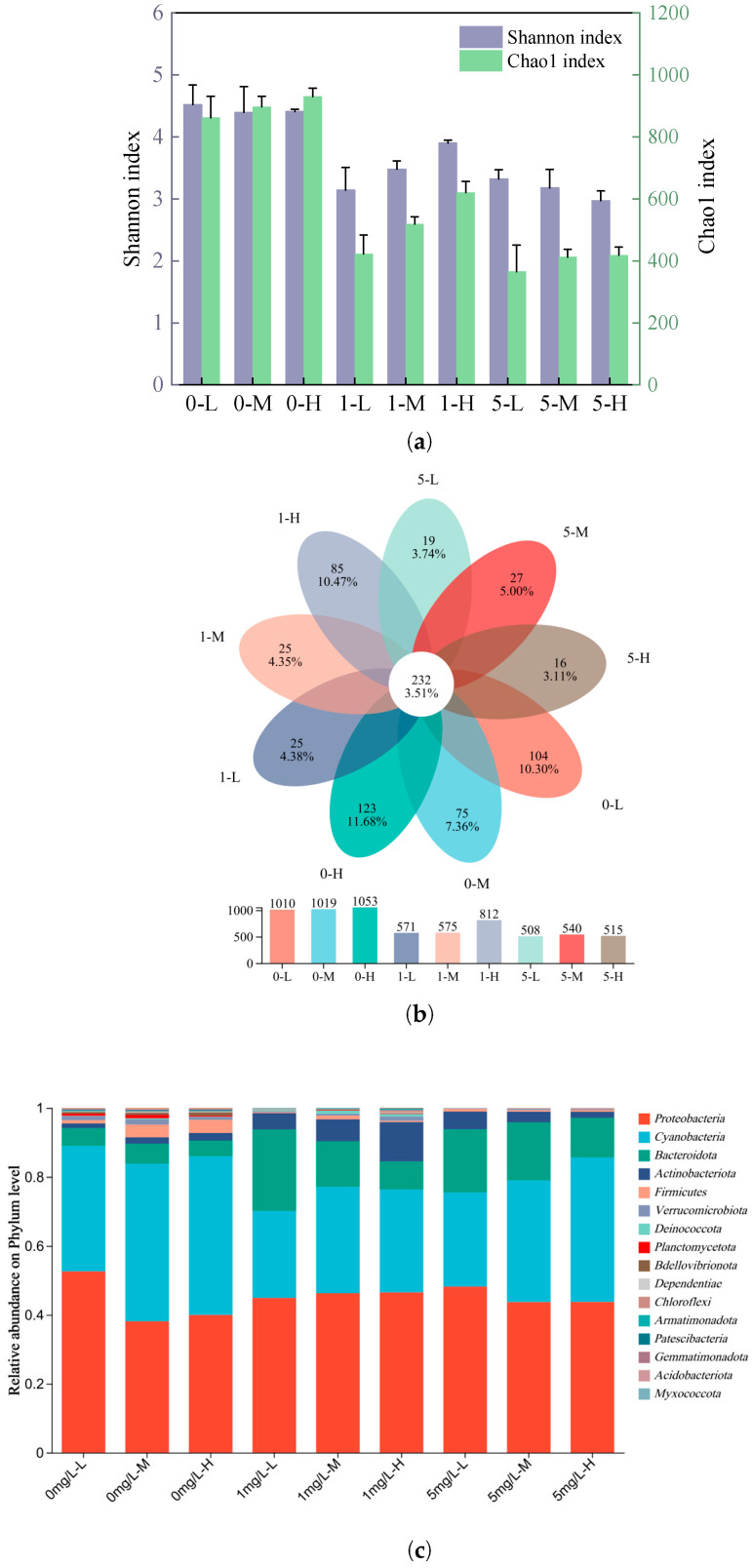 Figure 6
Figure 6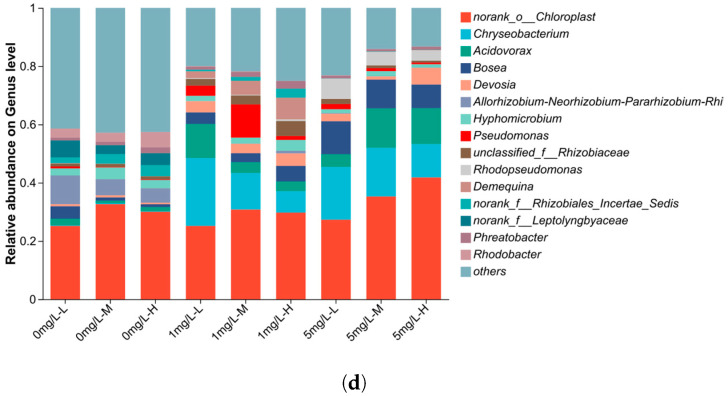 Figure 7
Figure 7 Figure 8
Figure 8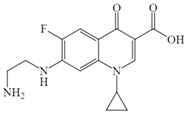 Figure 9
Figure 9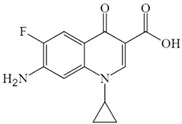 Figure 10
Figure 10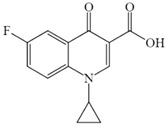 Figure 11
Figure 11- —National Natural Science Foundation of China
- —Open Project Funding of Key Laboratory of Intelligent Health Perception and Ecological Restoration of Rivers and Lakes, Ministry of Education, Hubei University of Technology
- —Open Project Funding of Hubei Key Laboratory of Environmental Geotechnology and Ecological Remediation for Lake & River, Hubei University of Technology
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPharmaceutical and Antibiotic Environmental Impacts · Pesticide and Herbicide Environmental Studies · Advanced oxidation water treatment
1. Introduction
Global antibiotic production and consumption have risen sharply in recent years. Human antibiotic use nearly doubled from 2018 to 2023, escalating from 40.1 billion defined daily doses (DDD) to 75.1 billion DDD [1,2]. Similarly, agricultural and aquaculture antibiotics are projected to increase from 99,502 tons in 2020 to over 107,000 tons by 2030 [3]. However, conventional wastewater treatment plants remove only limited amounts of antibiotics, leading to their continuous release and pseudo-persistence in aquatic environments [4,5]. Consequently, chronic low-dose exposure poses substantial ecotoxicological risks to aquatic organisms and threatens ecosystem structure and function [6].
As primary producers in shallow lakes, submerged macrophytes play a central role in organic matter production through light-driven photosynthesis. They also provide essential surfaces and substrates for epiphytic biofilms composed of algae, bacteria, and other microorganisms [7]. Importantly, submerged macrophytes contribute to nutrient cycling, pollutant removal, water quality improvement, and overall ecosystem stability [8]. However, direct exposure to antibiotics may disrupt plant physiology, leading to inhibited growth, reduced photosynthetic activity, and altered metabolic profiles. Such phytotoxic effects not only impair the ecological balance but also may weaken the pollutant-removal capacity of aquatic macrophytes [9]. Although several studies have explored antibiotic toxicity in submerged macrophytes, research remains fragmented and species coverage is limited. For example, enrofloxacin inhibited the growth of Vallisneria spiralis while simultaneously enhancing phosphorus uptake from water [10]. Exposure of Myriophyllum aquaticum to tetracycline suppressed plant growth and elevated MDA and antioxidant enzyme activity [11]. Likewise, sulfonamides disrupted the growth and photosynthesis of Vallisneria natans [12], and oxytetracycline and sulfadiazine altered physiological responses in V. natans and associated biofilms [13]. These studies indicate that antibiotics can trigger overproduction of reactive oxygen species (ROS) and disrupt photosynthetic efficiency, leading to cellular damage. However, interspecific variation in tolerance has also been observed, suggesting differences in antioxidant defense capacity among species. Despite its global prevalence and ecological importance, the response of H. verticillata to ciprofloxacin has not been systematically investigated.
Submerged macrophytes such as H. verticillata and V. natans are widely applied in phytoremediation due to their ability to improve water quality in eutrophic lakes by removing nitrogen and phosphorus from both water columns and sediments [14]. Phytoremediation is cost-effective, environmentally sustainable, and scalable. Nevertheless, its efficacy may be limited by plant sensitivity to contaminants such as antibiotics, which can reduce growth and pollutant uptake efficiency [15]. The ability of macrophytes to accumulate and metabolize antibiotics is a key determinant of their tolerance and remediation potential. Despite their proven ability to reduce antibiotic concentrations in water, research on the underlying degradation and transformation mechanisms in plants remains rare. For instance, V. natans removed 99.7% of sulfonamides within 13 days [12], M. aquaticum eliminated >85% of tetracyclines within 12 weeks [16], and Ceratophyllum demersum removed up to 44% of ciprofloxacin from aquaculture effluents [17]. Egeria densa achieved 58% ciprofloxacin removal within 96 hours [18]. While these results demonstrate the potential of submerged macrophytes to remove antibiotics, most studies have focused on pollutant reduction efficiency rather than elucidating metabolic pathways or identifying plant-derived antibiotic metabolites, which may exhibit distinct chemical structures and ecotoxicological properties compared to the parent compounds [19]. H. verticillata is a widely distributed submerged macrophyte known for its strong biological purification capacity and adaptability to diverse environmental conditions [20]. It can tolerate low light intensity, limited nutrient availability, and a wide range of physical and chemical water conditions, enabling it to outcompete other aquatic plants across broad habitats. These attributes make H. verticillata a particularly suitable candidate for phytoremediation of antibiotic-contaminated waters.
Therefore, a comprehensive understanding of antibiotic phytotoxicity, biodegradation, and metabolic transformations in submerged macrophytes is essential to optimize phytoremediation strategies and assess ecological risks of antibiotics. Ciprofloxacin (CIP), a widely used fluoroquinolone, is frequently detected in surface waters with median concentration of 0.164 mg/L and maximum concentration of 6.5 mg/L in freshwater ecosystem, posing considerable ecological risks [21,22]. This study systematically investigated (i) the phytotoxic effects of CIP on the growth and photosynthesis of H. verticillata, (ii) antioxidant defense responses including SOD and anthocyanin regulation, (iii) the biodegradation efficiency and metabolic pathways of CIP within plant tissues, and (iv) the associated changes in epiphytic bacterial communities of H. verticillata. This study provides new insights into the role of H. verticillata in CIP removal and the potential application of submerged macrophytes in phytoremediation of antibiotic-contaminated waters.
2. Materials and Methods
2.1. Plant Materials and Chemicals
H. verticillata used in this experiment was collected in July 2024 from a shallow lake near Hubei University of Technology. H. verticillata with green coloration, intact apical tips, and no visible signs of damage or algae infestation were chosen. Plants were carefully washed with deionized water to remove sediment and epiphytes. Prior to experiment, plants were pre-cultured in 10% Hoagland nutrient solution for 2 weeks under controlled conditions (25 ± 1 °C, light intensity 3000 lx, 12 h light/12 h dark cycle) to ensure uniform physiological status. CIP (CIP hydrochloride, ≥97% purity) was purchased from Aladdin (Shanghai, China). The CIP used in this study was in the form of CIP hydrochloride which is highly soluble in water. Stock solutions (1000 mg/L) were prepared in ultrapure water, filtered through 0.22 μm PTFE membranes (Tianjin Jinteng Experiment Equipment Co., Ltd., Tianjin, China), and stored at 4 °C in the dark until use. The filtration process did not cause any significant loss of CIP [23,24,25].
2.2. Batch Experiments
2.2.1. Ecotoxicological Effects of CIP on H. verticillata
The H. verticillata (15 cm in length, fresh weight 1.1 ± 0.1 g) were cultured in a 2 L container with 10% Hoagland solution and anchored with 50 mm glass beads. CIP was added at 0, 1, 2, 5, and 10 mg/L. These concentrations were chosen based on concentrations reported in aqueous environments and other toxicological studies [26]. A CIP-free control was included. All treatments were performed with three independent biological replicates. The plants were incubated in a growth chamber (25 °C, 3000 lx, 12 h light/12 h dark cycle) for 12 days. Evaporation losses were replenished daily with deionized water. At the end of the experiment, plants were harvested for biomass, growth, and biochemical analyses, and culture solutions were collected for antibiotic quantification. Relative growth rate was calculated according to reference [10].
2.2.2. Biodegradation of CIP by H. verticillata
To evaluate plant-mediated biodegradation of CIP by H. verticillata, pre-cultured plants were trimmed. Two CIP concentrations (1 and 5 mg/L) were tested under three biomass densities in container (fresh weight): low (1.4 g), medium (2.8 g), and high (4.2 g), simulating natural densities of submerged macrophytes in lakes [13]. Blank controls (CIP solution without plants) were included. All treatments were conducted in triplicate. Plants were cultured in a 2 L container with 10% Hoagland solution for 12 days under the same incubation conditions described above.
2.3. Assessment of Biochemical Component Changes in H. verticillata
Chlorophyll fluorescence was measured using a portable plant efficiency analyzer (PEA, Hansatech, UK). Leaves were dark-adapted for 15 min before determining the maximum photochemical efficiency of PSII (Fv/Fm) [27]. Chlorophyll and carotenoids were extracted in 95% methanol, and absorbance was measured at 665, 652, and 470 nm. Pigment concentrations were calculated according to reference [11]. SOD activity was determined by nitroblue tetrazolium (NBT) photoreduction [28]. Anthocyanins were extracted in acidified ethanol (1% HCl, v/v) and quantified at 530 nm using a UV–vis spectrophotometer, the relative concentration of astaxanthin was calculated by the change in absorbance [29].
2.4. Determination of CIP Concentrations and CIP Metabolites
2.4.1. CIP Quantification
CIP in culture media was quantified using HPLC (Ultimate 3000, Thermo Fisher Scientific, Waltham, MA, USA) with a C18 column (250 × 4.6 mm, 5 μm) at 40 °C [30]. The mobile phase was methanol/0.04% phosphoric acid buffer with 0.05% triethylamine (70:30, v/v). The flow rate was 1.0 mL/min, injection volume 10 μL, and UV detection wavelength 278 nm. The retention time of CIP was 5.97 min.
CIP in H. verticillata tissues was performed according to reference [31]. At the end of the exposure experiment, leaves and stems of H. verticillata were separately collected and freeze-dried for 24 h, followed by grinding into fine powder. Tissue powder was extracted with 5 mL acetonitrile by vortexing for 15 min. The extracts were centrifuged at 8000 r/min for 10 min, and the supernatants were filtered through 0.22 μm organic membranes. The filtrates were analysis using Q Exactive UHPLC-Q Exactive Orbitrap MS (Thermo Fisher Scientific, USA).
2.4.2. Metabolite Identification
CIP metabolites were analyzed using UHPLC-Q Exactive Orbitrap MS (Thermo Fisher Scientific, USA) with a ZORBAX SB-C18 column (2.1 × 50 mm) [6]. Culture samples were centrifuged and extracted three times with ethyl acetate. The extracts were dried over anhydrous Na_2_SO_4_, evaporated under N_2_ and redissolved in methanol. Chromatographic separation was performed with a binary gradient: (A) water with 0.1% formic acid and (B) methanol with 0.1% formic acid, injection volume 1 μL. Mass spectrometry was operated in positive electrospray ionization (ESI^+^) mode at 4.0 kV. Data were acquired in full MS/dd-MS² mode (resolution 70,000), and metabolites were annotated by comparison with the mzCloud and ChemSpider databases using Xcalibur 3.0 software.
2.5. DNA Extraction and High Throughout Sequencing
Apical 10 cm segments of H. verticillata were transferred into 50 mL centrifuge tubes with 45 mL PBS buffer, shaken for 30 min, ultrasonicated at 40 kHz for 10 min, and shaken again for 10 min. This was repeated three times until leaves regained their natural green color. The washing solution was centrifuged at 8000 rpm for 10 min, and the pellet was stored at −80 °C. Samples were subsequently sent to Shanghai Meiji Biomedical Technology Co., Ltd. (Shanghai, China) for microbial analysis [32].
Total microbial DNA was extracted using the E.Z.N.A.^®^ Soil DNA Kit (Omega Bio-tek, Norcross, GA, USA) according to the manufacturer’s instructions. DNA quality was checked by 1% agarose gel electrophoresis, and concentrations were quantified using a NanoDrop 2000 spectrophotometer. The V3–V4 hypervariable regions of the bacterial 16S rRNA gene were amplified using primers 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′) with sample-specific barcodes. PCR amplification was carried out in 20 μL reactions containing 4 μL 5× FastPfu buffer, 2 μL dNTPs (2.5 mM), 0.8 μL each primer (5 μM), 0.4 μL FastPfu polymerase, and 10 ng template DNA. Cycling conditions were: 95 °C for 3 min, followed by 27 cycles of 95 °C for 30 s, 55 °C for 30 s, 72 °C for 30 s, and a final extension at 72 °C for 10 min.
PCR products were purified using a PCR Clean-Up Kit (Yuhua, China) and quantified by Qubit 4.0. Libraries were constructed using the NEXTFLEX Rapid DNA-Seq Kit, and sequencing was performed on the Illumina NovaSeq 6000 platform (Illumina, San Diego, CA, USA) with 2 × 250 bp paired-end reads.
2.6. Statistical Analyses
All data were tested for normality and homogeneity of variance prior to analysis. One-way ANOVA was used to assess treatment effects, followed by least significant difference (LSD) and Duncan’s multiple range tests for pairwise comparisons (p < 0.05). Statistical analyses were conducted using IBM SPSS Statistics 25.0 (IBM, Armonk, NY, USA). Graphical visualizations were produced using Origin 9.0 (OriginLab, Northampton, MA, USA). Microbial data analyses were carried out on the Majorbio Cloud Platform (https://cloud.majorbio.com, accessed on 10 August 2025).
3. Results and Discussion
3.1. Effects of CIP the Growth of H. verticillate
Plant growth is a key endpoint in evaluating the phytotoxicity of CIP, as it reflects the capacity of submerged macrophytes to tolerate and potentially remediate xenobiotics. As shown in Figure 1a,b, the length and fresh weight of H. verticillata in the control group increased significantly after 12 days, indicating healthy growth under laboratory conditions. However, CIP exposure markedly suppressed growth. The inhibition of H. verticillata length was concentration-dependent, with reductions of 8.22%, 9.48%, 15.82%, and 15.28% at 1, 2, 5, and 10 mg/L CIP, respectively (Figure 1a). Fresh weight exhibited even greater sensitivity, with inhibition rates of 21.42%, 23.01%, 29.66%, and 27.50% at the corresponding concentrations (Figure 1b).
These results demonstrate that CIP exerted stronger inhibitory effects on biomass accumulation than on elongation. This pattern is consistent with previous studies on submerged macrophytes. For example, tetracycline significantly reduced the fresh weight of Myriophyllum aquaticum at 10 mg/L, with roots and leaves being more sensitive than stems [11]. Similarly, sulfamethoxazole exposure suppressed leaf and root elongation of Vallisneria natans, with biomass showing greater inhibition than morphological indices [12]. The higher sensitivity of biomass to xenobiotics suggests that fresh weight is a more reliable indicator of phytotoxic stress than length growth, likely due to the cumulative impacts of metabolic disruption and photosynthetic inhibition.
3.2. Effects of CIP the Photosynthesis of H. verticillata
CIP exposure significantly impaired the photosynthesis of H. verticillata. As shown in Figure 2a, the maximum photochemical efficiency (Fv/Fm) decreased sharply with increasing CIP concentrations. Compared with the control (0.792), Fv/Fm values dropped to 0.325, 0.206, 0.145, and 0.045 at 1, 2, 5, and 10 mg/L CIP, representing decreases of 59.01%, 73.99%, 81.73%, and 94.32%, respectively. The effects of CIP on the chlorophyll a, chlorophyll b content of H. verticillata are shown in Figure 2b. Relative to the control, chlorophyll a of H. verticillata decreased by 27.85%, 31.33%, 13.41%, and 36.10% at 1, 2, 5, and 10 mg/L, while chlorophyll b decreased by 28.17%, 22.33%, 3.84%, and 31.20%. Carotenoid levels were comparatively less affected (Figure 2c), showing only slight reductions under CIP exposure except at the highest concentration.
Chlorophyll fluorescence has been widely used to assess plant responses to stress [27]. Fv/Fm, an indicator of PSII efficiency to ROS accumulation [33], revealed severe photoinhibition in H. verticillata. Generally speaking, Fv/Fm values lower than 0.2 indicate severe PSII damage, highlighting strong, concentration-dependent photoinhibition [34]. The observed declines demonstrate that CIP disrupted PSII reaction centers and impaired electron transport, thereby reducing light energy conversion. Similar inhibition of PSII was observed in V. natans exposed to oxytetracycline and sulfadiazine [13], supporting that fluoroquinolones can significantly impair the photosynthetic machinery of submerged plants.
Chlorophylls and carotenoids are fundamental pigments for light harvesting, energy absorption, and photosynthetic metabolism. The strong reduction in chlorophyll a and b content observed here suggests that CIP interfered with chloroplast structure and chlorophyll biosynthesis, while also accelerating degradation via chlorophyllase activity [35,36]. Comparable chlorophyll reductions were reported in V. natans under sulfonamide stress [12], where photosynthetic inhibition translated into growth suppression. Notably, leaf yellowing observed in H. verticillata at 10 mg/L CIP corresponds with severe chlorophyll loss, serving as a visible indicator of phytotoxicity.
Carotenoids, in contrast, are key non-enzymatic antioxidants that scavenge ROS and protect chlorophyll under stress [37]. The relatively stable carotenoid levels in H. verticillata suggest a protective defense mechanism, consistent with previous findings where carotenoid stability was maintained under phenolic stress [38,39]. However, under 10 mg/L CIP, the marked decline in carotenoids indicates that antioxidant defenses were overwhelmed, leading to irreversible metabolic impairment.
3.3. Effects of Oxidative Stress by CIP on H. verticillate
Superoxide dismutase (SOD) activity exhibited a clear biphasic response to CIP exposure (Figure 3a). At 1, 2, and 5 mg/L CIP, SOD activity increased by 89.19%, 151.56%, and 233.69% relative to the control, peaking at 3.3-fold the control value under 5 mg/L exposure. However, at 10 mg/L, SOD activity dropped by 6.4% compared with the control, showing no significant difference (p > 0.05). Anthocyanin content followed a similar trend (Figure 3b). After 12 days of cultivation, all CIP-treated groups contained significantly more anthocyanins than the control. Increases were 168.99%, 144.94%, 71.52%, and 69.62% at 1, 2, 5, and 10 mg/L, respectively. The strongest accumulation occurred under 1–2 mg/L CIP exposure, while higher concentrations dampened this stimulatory effect.
SOD is the first line antioxidant enzyme in plants, catalyzing the dismutation of superoxide radicals into hydrogen peroxide and oxygen, thereby protecting membranes and organelles from oxidative damage. The sharp increase in SOD activity at 1–5 mg/L CIP suggests that H. verticillata initiated an active defense against CIP-induced reactive oxygen species (ROS). Such activation of antioxidant defenses has also been reported for Vallisneria natans, where low-level CIP exposure stimulated SOD activity but higher concentrations suppressed it [26]. The decline at 10 mg/L in H. verticillata indicates that ROS generation exceeded the enzymatic detoxification capacity, leading to disruption of antioxidant regulation and subsequent oxidative damage [40].
Anthocyanins, non-enzymatic antioxidants belonging to the flavonoid family, also responded in a biphasic pattern. At low CIP concentrations, enhanced ROS signaling likely activated transcriptional pathways for anthocyanin biosynthesis [41,42], providing additional protection against oxidative stress. However, at higher concentrations, anthocyanin accumulation was suppressed. This may reflect accelerated degradation of anthocyanins under severe oxidative conditions or inhibition of biosynthetic pathways caused by widespread metabolic impairment. Similar trends have been observed in Chlorella vulgaris exposed to cefazolin sodium, where anthocyanins first increased then declined at higher doses [43].
In general, antioxidant activity increases under relatively low levels of environmental stress but declines under more severe conditions due to irreversible cellular damage. The simultaneous increases in SOD and anthocyanin activities observed in this study indicate that both enzymatic and non-enzymatic antioxidants act synergistically to mitigate stress-induced damage. This hormesis-like response suggests that low concentrations of CIP stimulate antioxidant defenses, which holds significant implications for understanding plant tolerance mechanisms. Collectively, these results reveal a hormesis response of H. verticillata to CIP-induced oxidative stress: low-to-moderate CIP concentrations enhance antioxidant defenses, whereas excessive stress overwhelms the system, leading to antioxidant suppression. This different response underscores the importance of oxidative stress modulation in determining plant tolerance thresholds to antibiotics in aquatic environments.
3.4. Biodegradation Efficiency and Pathway of CIP by H. verticillata
CIP concentrations in the aqueous phase during the 12-day experiment are shown in Figure 4. In the control groups (without plants), CIP concentrations remained nearly constant at both 1 mg/L and 5 mg/L, confirming negligible abiotic loss. By contrast, treatments containing H. verticillata exhibited significant concentration declines. At 1 mg/L, removal efficiencies reached 12.33%, 12.31%, and 36.96% under low (L), medium (M), and high (H) planting densities, respectively. At 5 mg/L, corresponding removal efficiencies were 25.62%, 29.60%, and 30.69%. These results indicate that removal was biomass-dependent, with higher plant densities achieving greater removal, particularly under the 1 mg/L condition (Figure 4a). At 5 mg/L (Figure 4b), removal increased with biomass but tended to plateau between medium and high densities, suggesting a potential saturation of uptake or metabolic capacity.
CIP accumulation in plant tissues at the end of the experiment is presented in Figure 4c. In the 1 mg/L treatment, leaf accumulation reached 0.014, 0.006, and 0.003 mg/g in the L, M, and H groups, respectively, while stem accumulation was 0.024, 0.012, and 0.008 mg/g. In the 5 mg/L treatment, leaf accumulation increased to 0.085, 0.039, and 0.016 mg/g, while stem accumulation reached 0.115, 0.069, and 0.044 mg/g. Across treatments, stems consistently accumulated higher levels than leaves, and accumulation was substantially greater under 5 mg/L than under 1 mg/L. These findings suggest that H. verticillata contributes to CIP removal through a combination of uptake, adsorption, and potential metabolic degradation. The biomass dependence indicates that greater plant surface area and metabolic activity enhance removal efficiency. Notably, stems accumulated more CIP than leaves, likely due to differences in tissue composition and transport capacity, which may favor storage in structural tissues. Similar preferential accumulation in stems has been reported for sulfonamides in Vallisneria natans [12]. Moreover, many antibiotics can be absorbed and accumulated in various plant tissues. For instance, Myriophyllum aquaticum removed tetracycline from water, accumulated in roots, stems, and leaves [11]. Likewise, CIP has been reported to accumulate in the roots of Eichhornia crassipes. Sufficient lipophilicity allows CIP to cross the lipid bilayer of plant cell membranes, while adequate hydrophilicity facilitates its translocation through cell sap [44]. These results indicate that antibiotic accumulation in aquatic plants is not limited to passive adsorption but also involves active transportation and partition. Compared with advanced oxidation processes (AOPs), the removal capacity of H. verticillata remains limited. However, since antibiotics typically occur in aquatic environments at concentrations ranging from ng/L to μg/L, the performance of H. verticillata in removing CIP under such environmentally relevant concentrations and at larger scales remains promising and needs further investigation.
To elucidate the mechanism of CIP biodegradation in the H. verticillata cultivation system, degradation intermediates were identified using UHPLC-MS/MS. A total of three metabolites C306, C263, and C248 were detected, with their molecular formulas, molecular weights, and proposed structures summarized in Table 1. Details on their MS spectra are summarized in the Supplementary Material (Figures S1–S4). Based on these results, a possible biodegradation pathway of CIP is proposed in Figure 5. Specifically, CIP undergoes partial ring-opening to yield C306. Subsequent removal of the entire piperazine ring produces C263, which is further deaminated to form C248.
In this study, analysis of intermediate products supports the hypothesis that H. verticillata contributes to active biodegradation of CIP. The proposed degradation pathway involves sequential N-deethylation to form C306, piperazine-ring cleavage to produce C263, and deamination yielding C248. Such reactions are consistent with the chemical reactivity of fluoroquinolones, as the piperazine moiety is particularly prone to substitution or decomposition via deethylation and deamination [45]. Indeed, C306 has been identified as a CIP metabolite in nitrifying biofilm systems [45], and also detected during CIP degradation by Gloeophyllum striatum [46,47]. Similarly, C263, lacking the piperazine moiety at the C7 position, was observed in the roots of Vetiveria zizanioides exposed to CIP [31]. These findings confirm that the transformation products observed in H. verticillata align with well-documented biodegradation routes of fluoroquinolones in both microbial and plant systems.
3.5. Effects of CIP on the Epiphytic Bacterial Communities of H. verticillata
To investigate the impact of CIP on bacterial communities associated with biodegradation, 16S rRNA sequencing was conducted. Alpha diversity indices (Shannon and Chao1) showed that CIP exposure significantly reduced both bacterial richness and diversity compared with controls (Figure 6a), indicating that CIP stress simplified the structural complexity of epiphytic microbiomes. A Venn diagram further revealed that CIP markedly reduced the number of unique OTUs (Figure 6b), suggesting a decline in microbial richness and overlap. Such losses are consistent with the selective pressure of antibiotics, which favor resistant populations while suppressing sensitive taxa [48,49].
At the phylum level (Figure 6c), Proteobacteria and Cyanobacteria were the dominant groups, accounting for more than 70% of the communities, followed by Bacteroidota, Actinobacteria, and Firmicutes. CIP exposure increased Actinobacteria and Bacteroidota while reducing Firmicutes, reflecting differential resistance of microorganism. The mutations in the DNA gyrase (gyrA/B) of Actinobacteria can reduce CIP binding affinity [50], while Bacteroidota utilize efflux pumps AcrAB-TolC to expel antibiotics [51]. Consistent with our results, CIP exposure has been shown to enhance the abundance of Actinobacteria in biofilms associated with Vallisneria natans [52]. Conversely, the decline of Firmicutes may be explained by their selective inhibition under fluoroquinolone stress [53].
At the genus level (Figure 6d), norank_o__Chloroplast, Chryseobacterium, Acidovorax, Bosea, and Devosia were particularly abundant. Norank_o__Chloroplast sequences, derived from chloroplasts, represent photosynthetic organisms capable of oxygen release during daytime, which may interfere with denitrification processes [54]. Their abundance increased with plant biomass, likely reflecting enhanced epiphytic colonization. Chryseobacterium are Gram-negative rods with intrinsic antibiotic resistance [55]. Acidovorax, also Gram-negative, have capacity to degrade polycyclic aromatic hydrocarbons (PAH) phenanthrene [56]. Bosea can produce β-lactamases and L,D-transpeptidases or homologous enzymes that enable the degradation of antibiotics such as amoxicillin [57]. Devosia, belonging to the Proteobacteria, are root-colonizing bacteria that can directly promote plant growth [58], and Proteobacteria were the main microbes to remediate piperazine contained wastewater which similar to the degradation pathway of CIP [59]. The increased abundance of these genera under CIP exposure indicates both antibiotic resistance and potential roles in antibiotic biodegradation. Collectively, these results demonstrate that CIP exposure alters the microbial ecology of H. verticillata associated communities, reducing the diversity while selectively enriching resistant genera. Such community shifts may synergistically contribute to the observed biodegradation of CIP through plant microbe interactions.
4. Conclusions
This study demonstrated that CIP exerts phytotoxicity on H. verticillata through inhibited growth, reduced photosynthetic efficiency, and oxidative stress imbalance. Nevertheless, H. verticillata contributes to CIP removal in aquatic systems. Three degradation intermediates (C306, C263, and C248) were identified, and the possible biodegradation pathway was proposed, involving de-ethylation, piperazine ring cleavage, and deamination. Furthermore, CIP exposure alters the epiphytic microbial community, favoring antibiotic-resistant genera while reducing overall bacterial diversity. These findings indicate that H. verticillata is a potential agent for CIP removal from aquatic environments in phytoremediation. Future research should focus on long-term exposure experiments at environmentally relevant concentrations to fully evaluate the practical applicability of H. verticillata in phytoremediation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Browne A.J. Chipeta M.G. Haines-Woodhouse G. Kumaran E.P.A. Hamadani B.H.K. Zaraa S. Henry N.J. Deshpande A. Reiner R.C. Day N.P.J. Global Antibiotic Consumption and Usage in Humans, 2000–18: A Spatial Modelling Study Lancet Planet Health 20215 e 893e 90410.1016/S 2542-5196(21)00280-134774223 PMC 8654683 · doi ↗ · pubmed ↗
- 2Klein E.Y. Impalli I. Poleon S. Denoel P. Cipriano M. Van Boeckel T.P. Pecetta S. Bloom D.E. Nandi A. Global Trends in Antibiotic Consumption during 2016–2023 and Future Projections through 2030 Proc. Natl. Acad. Sci. USA 2024121 e 241191912110.1073/pnas.241191912139556760 PMC 11626136 · doi ↗ · pubmed ↗
- 3Mulchandani R. Wang Y. Gilbert M. Van Boeckel T.P. Global Trends in Antimicrobial Use in Food-Producing Animals: 2020 to 2030 PLOS Glob. Public Health 20233 e 000130510.1371/journal.pgph.000130536963007 PMC 10021213 · doi ↗ · pubmed ↗
- 4Kovalakova P. Cizmas L. Mc Donald T.J. Marsalek B. Feng M. Sharma V.K. Occurrence and Toxicity of Antibiotics in the Aquatic Environment: A Review Chemosphere 202025112635110.1016/j.chemosphere.2020.12635132443222 · doi ↗ · pubmed ↗
- 5Ren Z. Xu H. Wang Y. Li Y. Han S. Ren J. Combined Toxicity Characteristics and Regulation of Residual Quinolone Antibiotics in Water Environment Chemosphere 202126312830110.1016/j.chemosphere.2020.12830133297242 · doi ↗ · pubmed ↗
- 6Wan L. Wu Y. Zhang Y. Zhang W. Toxicity, Biodegradation of Moxifloxacin and Gatifloxacin on Chlamydomonas Reinhardtii and Their Metabolic Fate Ecotoxicol. Environ. Saf.202224011371110.1016/j.ecoenv.2022.11371135653971 · doi ↗ · pubmed ↗
- 7Zhao Z. Qin Z. Xia L. Zhang D. Hussain J. Dissipation Characteristics of Pyrene and Ecological Contribution of Submerged Macrophytes and Their Biofilms-Leaves in Constructed Wetland Bioresour. Technol.201826715816610.1016/j.biortech.2018.06.06030014994 · doi ↗ · pubmed ↗
- 8Zhang H. Luo X. Li Q. Huang S. Wang N. Zhang D. Zhang J. Zheng Z. Response of the Submerged Macrophytes Vallisneria natans to Snails at Different Densities Ecotoxicol. Environ. Saf.202019411037310.1016/j.ecoenv.2020.11037332151866 · doi ↗ · pubmed ↗