Genotype-Specific Vector Competence of Aedes albopictus for Japanese Encephalitis Virus Genotypes I, III, and V
Bo-Ram Yun, Ji-Young Kwon, Dongmi Kwak, Hee Il Lee

TL;DR
This study shows that Aedes albopictus mosquitoes can transmit certain types of Japanese encephalitis virus more effectively than others.
Contribution
The study reveals genotype-specific vector competence of Ae. albopictus for JEV genotypes I, III, and V.
Findings
Ae. albopictus showed high competence for JEV genotype V with 100% dissemination rate and 71.7% head–thorax positivity.
Genotype III had moderate competence with 76.9% dissemination rate but low transmission rate at 6.6%.
Genotype I showed minimal infection and negligible transmission potential in Ae. albopictus.
Abstract
Japanese encephalitis virus (JEV), a mosquito-borne flavivirus, poses a significant public health threat in Asia. Although Culex species are primary vectors, the role of Aedes albopictus in JEV transmission has gained attention under changing ecological conditions. This study evaluated the vector competence of Ae. albopictus for three JEV genotypes: I (GI), III (GIII), and V (GV). Laboratory-reared Ae. albopictus were orally challenged with each genotype, and infection rate (IR), dissemination rate (DR), head–thorax positivity rate (HTR, proxy for potential transmission), and transmission rate (defined as saliva positivity) were assessed at 7 and 14 days post-infection (dpi). Ae. albopictus showed marked genotype-specific differences. By 14 dpi, GV had the highest DR (100.0%) and HTR (71.7%), with viral RNA detected in 36.7% of TR. GIII showed moderate competence (76.9% DR, 39.3% HTR),…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
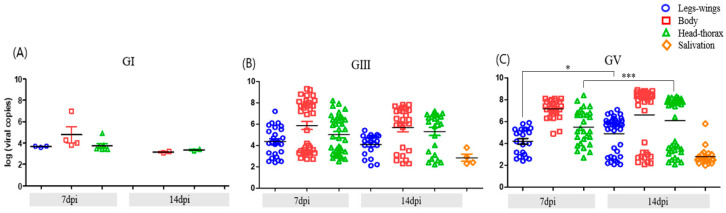 Figure 1
Figure 1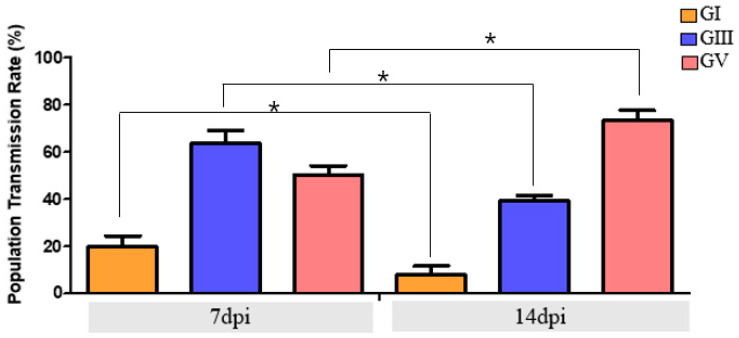 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMosquito-borne diseases and control · Dengue and Mosquito Control Research · Malaria Research and Control
1. Introduction
Japanese encephalitis virus (JEV), a mosquito-borne flavivirus, is the leading cause of viral encephalitis in Asia, responsible for an estimated 68,000 cases and over 10,000 deaths annually, predominantly among children in endemic regions [1]. The virus is maintained in a zoonotic transmission cycle involving Culex mosquitoes, particularly Culex tritaeniorhynchus, and vertebrate amplifying hosts, such as pigs and ardeid birds [2]. Human infections are incidental but can lead to outbreaks under favorable ecological and climatic conditions [3,4,5,6].
Among the five recognized JEV genotypes (GI–GV), genotype III (GIII) was the first to be identified, initially isolated in Japan in 1935 [7,8]. Genotype II (GII) and genotype V (GV) were subsequently reported in the 1950s from India and Malaysia, respectively [9,10], while genotype IV (GIV) was detected in Indonesia in 1979. Genotype I (GI), now the dominant strain across much of Asia, emerged in East Asia in the late 1970s [2,11,12,13].
Historically, GIII was the predominant genotype circulating in East and Southeast Asia. However, over the past two decades, GI has largely displaced GIII as the dominant strain in many regions [14]. Notably, GV, once considered extinct following its initial isolation in Malaysia, re-emerged in 2009 when it was detected in Cx. tritaeniorhynchus in China [15]. In the Republic of Korea (ROK), GV was first identified in 2010 in Cx. bitaeniorhynchus, with subsequent detections in other Culex species, including Cx. orientalis in 2020, indicating its sustained circulation [14,15,16]. GIV has also been reported, mainly in Indonesia and parts of Oceania, but its limited geographic distribution and relatively low public health impact have resulted in sparse investigation [17].
Although Culex species remain the principal vectors of JEV, accumulating evidence suggests that Aedes albopictus may also contribute to its transmission, particularly in ecologically altered or urban environments [18,19,20]. Commonly known as the Asian tiger mosquito, Ae. albopictus is a highly adaptable species with a global distribution facilitated by international trade and climate change. In ROK, Ae. albopictus is already widely distributed nationwide [21]. JEV RNA has been detected in field-collected Ae. albopictus in ROK [22]. While repeated isolations of JEV from Ae. albopictus have been reported in Asia, the primary vectors remain Culex species, and the epidemiological significance of Ae. albopictus in natural transmission cycles is still unclear [23,24,25]. The established competence of Ae. albopictus for transmitting other arboviruses—including dengue, chikungunya, and Zika—warrants further investigation into its potential role in JEV transmission [19,20,23,26,27].
Vector competence, the intrinsic ability of a mosquito to acquire, sustain, and transmit a pathogen, varies with both mosquito species and viral genotype. Prior studies have demonstrated that susceptibility to JEV can vary among mosquito species, populations, and viral genotypes, highlighting the importance of investigating local vector competence for both primary and potential supplementary vectors [18,20,21,28]. However, comparative data on Ae. albopictus responses to different JEV genotypes remain limited. In particular, the ability of Ae. albopictus to support replication and transmission of the re-emerging GV genotype is poorly characterized. This gap is especially critical given the recent predominance of GV in ROK, underscoring the need for localized assessments of vector competence.
Given the ongoing genotype shifts in JEV and the expanding ecological range of Ae. albopictus, it is essential to evaluate its potential role in transmitting diverse JEV genotypes. In this study, we assessed the vector competence of laboratory-reared Ae. albopictus for JEV GI, GIII, and GV. Using oral infection assays, we evaluated genotype-specific differences in infection, dissemination, and transmission at multiple time points post-infection. These findings aim to enhance understanding of genotype-dependent transmission dynamics and inform future risk assessments and vector control strategies in the context of evolving JEV ecology.
2. Materials and Methods
2.1. Mosquitoes
Ae. albopictus (F43 generation), originally collected from Incheon, ROK, were used for experimental infections. Following field collection, the colony was continuously maintained under laboratory conditions. Mosquitoes were reared at 27 ± 1 °C, 70 ± 5% relative humidity, and a 12:12 h light–dark photoperiod. Adults were provided with 10% sucrose ad libitum, and females were blood-fed weekly using an artificial membrane feeding system (Hemotek Ltd., Accrington, UK). Each genotype was tested in three independent biological replicates, using newly prepared infectious blood meals and separate cohorts of mosquitoes.
2.2. Production and Titration of JEV Strains for Vector Competence Assays
JEV GI was isolated from Cx. tritaeniorhynchus in 2005 [29], GIII from the same species in 1994 [29], and GV from Cx. orientalis in 2020 [16] (Table 1). Viral stocks were propagated in Vero cells as previously described [19,30]. Briefly, each strain was cultured in Eagle’s Minimum Essential Medium (MEM; Welgene, Gyeongsan, Republic of Korea) supplemented with 10% fetal bovine serum (FBS; Thermo Fisher Scientific, Waltham, MA, USA), 100 U/mL penicillin, and 100 μg/mL streptomycin at 37 °C in a humidified incubator (Vision Scientific Co.,Ltd., Daejeon, Republic of Korea) with 5% CO_2_ for 3 days in T75 flasks. Viral replication was confirmed by the presence of cytopathic effects (CPE) observed under a light microscope (ZEISS, Oberkochen, Germany). For virus titration, BHK-21 cells were seeded in six-well plates(SPL, Pocheon, Republic of Korea) 1 day prior to infection. Serial 10-fold dilutions of each viral stock were prepared in MEM containing 2% FBS and 1% penicillin–streptomycin (P/S), and 300 μL of each dilution was inoculated into individual wells. Following 1 h of incubation at 37 °C with 5% CO_2_, the inoculum was replaced with 4 mL of overlay medium containing 0.5% agarose, 2% FBS, and 1% P/S in MEM. After 5 days of incubation at 37 °C, the cells were fixed with 4% paraformaldehyde for 30 min and stained with 1% crystal violet. Plaques were counted, and viral titers are expressed as plaque-forming units per milliliter (PFU/mL).
2.3. Oral Infection of JEV
All infection assays were conducted in a Biosafety Level 3 (BSL-3) facility (Permit No. KCDC-18-3-04) at the Division of Vectors and Parasitic Diseases, Korea Disease Control and Prevention Agency (KDCA). Female Ae. albopictus mosquitoes (5–7 days old) were deprived of sucrose for 24 h prior to exposure. Infectious blood meals were prepared by mixing defibrinated sheep blood (Kisan Bio Co., Ltd., Seoul, Republic of Korea) with virus suspension at a 2:1 ratio, supplemented with ATP to a final concentration of 10 nM to stimulate feeding. Viral titers in the blood meals were adjusted to 6.5 × 10^6^–1.0 × 10^7^ PFU/mL for GI, 1.2 × 10^6^–1.0 × 10^7^ PFU/mL for GIII, and 1.0 × 10^6^–1.0 × 10^7^ PFU/mL for GV. Mosquitoes were allowed to feed overnight in a dark incubator at 27 ± 1 °C using a Hemotek membrane feeding system (Hemotek Ltd., Blackburn, UK). Fully engorged females were selected and maintained under standard rearing conditions (27 ± 1 °C, 70 ± 5% relative humidity, 12:12 h light–dark cycle) for the remainder of the experiment.
2.4. Mosquito Dissection and Saliva Collection
To evaluate infection dynamics, mosquitoes were dissected at 7 and 14 days post-infection (dpi) in the BSL-3 insectary under controlled conditions (27 ± 1 °C, 70 ± 5% relative humidity). For each individual, the midgut, legs and wings, and head–thorax were carefully separated and transferred into individual microcentrifuge tubes (SPL, Pocheon, Republic of Korea) containing 300 μL of MEM supplemented with 2% FBS and a ceramic homogenization bead. These tissues were used to assess infection (midgut), dissemination (legs and wings), and head–thorax positivity as a proxy for potential salivary gland invasion. At 14 dpi, saliva was also collected to assess transmissibility. Cold-anesthetized mosquitoes had their legs and wings removed, and their proboscis was inserted into a 10 μL droplet of phosphate-buffered saline (PBS) held within a cut pipette tip under the same environmental conditions (27 ± 1 °C, 70 ± 5% RH). After 30 min of salivation, saliva droplets were collected. All dissected tissues and saliva samples were stored at −80 °C until analysis.
Infection rate (IR), dissemination rate (DR), and transmission rate (TR) were determined based on the detection of viral RNA in specific tissues. IR was defined as the proportion of mosquitoes with viral RNA detected in the midgut, indicating successful initial infection. DR was calculated as the proportion of infected mosquitoes with viral RNA in the legs and wings, representing viral dissemination beyond the midgut barrier. Head–thorax positivity rate (HTR) was defined as the proportion of mosquitoes with viral RNA detected in the head–thorax, serving as a proxy for potential transmission.
In accordance with the accepted definition, TR is defined strictly as the proportion of mosquitoes with viral RNA detected in saliva, reflecting actual transmission potential. In this study, HTR is presented alongside TR as a complementary measure, since saliva-based assays may underestimate transmission due to methodological limitations.
2.5. RNA Extraction and qRT-PCR Detection of JEV
All tissue samples, excluding saliva, were homogenized using the Precellys^®^ Evolution homogenizer (Bertin Technologies, Montigny-le-Bretonneux, France) with two cycles of bead beating at 7500 rpm for 30 s each. Homogenates were centrifuged at 13,000 rpm for 1 min, and 30 μL of the supernatant was transferred to a new tube for RNA extraction. Total RNA was extracted using the Clear-S™ Total RNA Extraction Kit (Invirustech, Gwangju, Republic of Korea) according to the manufacturer’s instructions.
JEV RNA targeting the non-structural protein 5 (NS5) gene was detected using the Clear-MD^®^ Flavivirus Real-Time RT-PCR Detection Kit (Invirustech, Gwangju, Republic of Korea). The qRT-PCR conditions were as follows: reverse transcription at 45 °C for 10 min; enzyme inactivation at 95 °C for 10 min; followed by 40 cycles of amplification consisting of denaturation at 95 °C for 10 s, annealing at 60 °C for 15 s, extension at 72 °C for 10 s, and signal acquisition at 80 °C for 15 s. A melting curve analysis was subsequently performed to confirm product specificity: denaturation at 95 °C for 30 s, annealing at 70 °C for 30 s, followed by gradual heating to 95 °C in 0.5 °C increments every 30 s. Samples with a cycle threshold (Ct) value ≤ 40 and a melting peak between 82 °C and 88 °C were considered positive for JEV RNA. Quantification was calibrated using a standard curve generated from a Vircell JEV RNA quantified control (MBC134-R; AMPLIRUN Japanese Encephalitis RNA control, Granada, Spain, 2024) using 10-fold serial dilutions (10^5^–10^3^ copies per reaction); the resulting standard-curve data are provided in Supplementary Figure S1.
2.6. Viral Titration by TCID50 Assay
To quantify infectious virus, samples from the legs–wings, body, and head–thorax were collected at 7 and 14 dpi. Each tissue homogenate (30 μL) was used for titration via the 50% tissue culture infectious dose (TCID_50_/mL) assay using BHK-21 cells. BHK-21 cells were seeded into 96-well plates at a density of 1.0 × 10^4^ cells/well in 200 μL of MEM supplemented with 5% FBS and 1% penicillin–streptomycin (P/S), and incubated overnight at 37 °C. Serial 10-fold dilutions of each homogenate were prepared in MEM containing 2% FBS, and 100 μL of each dilution was added to five replicate wells. Plates were incubated at 37 °C for 5 days, after which CPE were visually assessed. Cells were subsequently fixed and stained with crystal violet to confirm CPE. Viral titers were calculated using the Reed–Muench method based on the number of CPE-positive wells at each dilution [3].
2.7. Statistical Analysis
All statistical analyses were performed using GraphPad Prism version 5.0 (GraphPad Software, San Diego, CA, USA). Comparisons of viral RNA loads between time points within each tissue were assessed using the nonparametric Mann–Whitney U test. For comparisons across multiple groups (e.g., among JEV genotypes), the Kruskal–Wallis test, followed by Dunn’s multiple comparisons test, was applied. Differences in HTR (proxy for potential transmission) and TR (defined as saliva positivity) between 7 and 14 dpi were analyzed for each genotype using Fisher’s exact test. PTR (population transmission rate) was calculated as the proportion of saliva-positive mosquitoes among blood-fed survivors. A p-value < 0.05 was considered statistically significant.
3. Results
3.1. Genotype-Dependent Differences in the Vector Competence of Ae. albopictus
The vector competence of Ae. albopictus for three JEV genotypes (GI, GIII, GV) was assessed based on IR, DR, HTR, and TR (saliva positivity) at 7 and 14 dpi (Table 2). The data revealed pronounced genotype-specific differences. GV-infected mosquitoes displayed the highest competence at 14 dpi, with a DR of 100.0% and HTR of 71.7%. However, the TR, defined as saliva positivity, was 36.7%, indicating that although many mosquitoes showed evidence of potential salivary gland invasion, only about one-third were capable of viral excretion. Mosquitoes infected with GIII showed intermediate competence. At 14 dpi, DR reached 76.9% and HTR was 39.3%, but the corresponding TR was only 6.6%, suggesting inefficient release of virus into saliva despite moderate dissemination and potential salivary gland infection.
In contrast, GI-infected mosquitoes exhibited minimal competence. At 14 dpi, dissemination was not observed beyond the midgut in most individuals, and the TR was only 4.0%, confirming negligible transmission potential for this genotype in Ae. albopictus. Together, these results demonstrate that while HTR provides an indicator of potential transmission, TR based on saliva positivity represents the conservative and accepted measure of actual transmission capacity, which varied markedly among JEV genotypes.
3.2. Temporal Dynamics of Viral Dissemination and Transmission
Temporal analysis revealed efficient progression of GV infection within mosquito tissues between 7 and 14 dpi. A marked increase in viral RNA detection in both the head–thorax region and saliva was observed at 14 dpi, indicating successful viral escape from tissue barriers and a high probability of transmission. In GIII-infected mosquitoes, dissemination to secondary tissues was already apparent by 7 dpi and remained relatively stable thereafter, with only a modest increase over time. In contrast, GI infection showed no substantial progression during the same period, remaining largely restricted to the midgut. These patterns underscore distinct temporal dynamics in viral replication and dissemination across JEV genotypes (Figure 1).
3.3. Infectious Viral Titers in Ae. albopictus
To determine the presence of infectious virus, TCID_50_ assays were performed on mosquito body, legs–wings, and head–thorax samples at 7 and 14 dpi (Table 3). Infectious JEV was consistently detected in GIII-infected mosquitoes. Mean viral titers in the body were 2.39 ± 0.10 log_10_ TCID_50_/mL (n = 9) at 7 dpi and 2.30 ± 0.08 log_10_ TCID_50_/mL (n = 5) at 14 dpi. Although viral dissemination to the head–thorax was observed in a limited number of mosquitoes, the corresponding titers remained low, 2.33 and 2.35 log_10_ TCID_50_/mL at 7 and 14 dpi, respectively.
3.4. Transmission Potential of Ae. albopictus at the Population Level
To evaluate the population-level transmission potential of Ae. albopictus, viral RNA was quantified in both the head–thorax region (head–thorax positivity rate, indicating potential transmission) and saliva (reflecting actual transmission) (Figure 2). At 14 dpi, GV-infected mosquitoes exhibited the highest HTR (71.7%) based on head–thorax positivity; however, the TR was 36.7%, providing the most accurate indicator of transmission. GIII-infected mosquitoes showed an HTR of 39.3%, but only 6.6% TR, reflecting a strong barrier to viral excretion. GI-infected mosquitoes exhibited minimal competence, with just 4.0% TR, confirming negligible transmission potential. These comparisons highlight that while HTP provides a proxy measure of potential salivary gland invasion, TR based on saliva positivity represents the conservative and accepted indicator of true transmission capacity. These findings indicate a markedly reduced ability of GI to escape midgut and salivary barriers, further highlighting the limited transmission potential of this genotype in Ae. albopictus.
4. Discussion
This study provides novel insights into the genotype-specific vector competence of Ae. albopictus for JEV, with particular emphasis on the recently re-emerged GV. Under controlled laboratory conditions, Ae. albopictus supported the infection, dissemination, and transmission of JEV GV at levels comparable to, or exceeding, those of GIII, a historically dominant strain. In contrast, GI, currently the predominant circulating strain in East Asia, exhibited markedly reduced vector competence, with limited midgut infection and negligible transmission potential [30,31].
In line with the accepted definition, TR was defined strictly as the proportion of mosquitoes with detectable JEV RNA in saliva [32,33]. To provide additional context on viral dissemination within the vector, we also evaluated population transmission rates based on HTR as a proxy for potential transmission. HTR reflects possible salivary gland invasion but may overestimate actual transmission capacity, since the presence of viral RNA in the head–thorax does not guarantee viral excretion. Conversely, TR offers the most conservative and reliable indicator of true transmission, although it may underestimate potential because forced salivation methods can fail to detect low-level expectoration [31,34,35,36,37,38,39]. Accordingly, HTR and TR should be interpreted as complementary rather than contradictory indicators of vector competence [36].
Our results demonstrated that GV-infected mosquitoes had both high HTR (71.7%) and substantial TR (36.7%), indicating efficient dissemination and partial but effective escape into saliva. For GIII, HTR was 39.3%, but TR was only 6.6%, suggesting a pronounced bottleneck at the level of viral release into saliva. For GI, the TR was just 4.0%, confirming negligible transmission potential. These genotype-dependent differences highlight that viral replication and dissemination kinetics within the mosquito strongly influence ultimate transmission potential [18,19].
A notable finding was the poor vector competence of Ae. albopictus for GI. Despite its predominance in current human and animal infections, GI rarely disseminated beyond the midgut in infected mosquitoes, and transmission was undetectable. This suggests that GI exhibited low replication efficiency in Ae. albopictus and instead relied primarily on other vectors, such as Cx. tritaeniorhynchus, for efficient transmission. Alternatively, genotype-specific immune responses, such as enhanced activation of RNA interference or other antiviral pathways in Ae. albopictus, may suppress GI replication. These findings highlight the complexity of genotype–vector compatibility, which is shaped by the interplay between viral genetics and vector physiology. This interplay is also strongly influenced by ecological and climatic factors [27]. Consistent with this observation, the number of GI-positive mosquitoes was markedly lower at 14 dpi (n = 7), despite a total of 106 individuals being engorged with GI-infected blood meals across three independent replicates. In contrast, 38 and 29 mosquitoes remained positive for GIII and GV, respectively, at 7 dpi. This uneven sample size reflects the inherently reduced replication and persistence of GI in Ae. albopictus rather than insufficient experimental design, further supporting genotype-specific differences in viral replication dynamics.
Temporal infection dynamics further emphasized the differences among the genotypes. GV demonstrated rapid progression from infection to head–thorax positivity (TR) and saliva positivity between 7 and 14 dpi, consistent with high replication efficiency and systemic dissemination. GIII exhibited early dissemination by 7 dpi but showed limited progression thereafter, while GI remained largely restricted to the midgut throughout. These trends were corroborated by our temporal analysis (Section 3.2), which showed a marked increase in viral RNA in the head–thorax and saliva of GV-infected mosquitoes at 14 dpi, indicating effective escape from tissue barriers. In contrast, GIII showed minimal change after initial dissemination, and GI failed to progress meaningfully over time. Together, these findings demonstrate that intra-vector replication kinetics are a critical determinant of genotype-specific transmission potential [2,19]. Such genotype dynamics are also shaped by repeated introductions across regions, as shown by molecular studies indicating frequent incursions of JEV from Southeast and East Asia into Japan [37]. Consistent with saliva assay results, only 4.0% of GI-infected mosquitoes were saliva-positive, further confirming the negligible transmission potential of this genotype in Ae. albopictus.
Taken together, our findings indicate that Ae. albopictus exhibits only low to moderate transmission competence for JEV when evaluated using saliva positivity as the benchmark. This conservative interpretation aligns with entomological surveillance in the ROK where JEV detections in Ae. albopictus have been rare (two positive pools out of >40,000 tested) [16,22]. Although experimental infections have demonstrated that Ae. albopictus is a competent laboratory vector for more than 20 arboviruses [37,38], field evidence supports its major role only for chikungunya virus and, to a lesser extent, dengue and Zika viruses. For JEV, while repeated isolations from Ae. albopictus and other mosquitoes have been reported in Asia [24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39], these should be interpreted with caution. Virus isolation alone is insufficient to incriminate a species as a natural vector; this requires supporting evidence of host feeding patterns, natural infection dynamics, and efficient transmission to vertebrate hosts [38]. Thus, while Culex species remain the principal vectors of JEV, the supplementary role of Ae. albopictus—particularly for GV—in urban and peri-urban environments cannot be dismissed [18,37,39,40].
This study has several limitations. First, all experiments were conducted under controlled laboratory conditions using a laboratory-reared mosquito colony; therefore, environmental temperature fluctuations, host availability, microbiota composition, and vector age—factors known to influence vector competence—were not captured. Future investigations should validate these findings under semi-field or field conditions and elucidate the molecular mechanisms that underlie genotype-specific susceptibility in Ae. albopictus.
Second, although inoculated viral titers differed slightly among JEV genotypes, all infectious blood meals were maintained within the biologically relevant range of 10^6^–10^7^ PFU/mL commonly used in vector-competence studies; thus, minor variation within this window is unlikely to have substantially influenced infection outcomes [34]. Third, although saliva positivity (TR) is the most direct measure of transmission, it may underestimate potential due to technical limitations of forced salivation, whereas head–thorax (HTR) may overestimate it; thus, both indicators should be interpreted as complementary rather than contradictory [31,33,34]. Finally, to further assess external transmissibility, future studies will include mammalian infection/challenge experiments to validate transmission potential in vertebrate hosts.
In conclusion, our results demonstrate that Ae. albopictus is a competent but limited vector for JEV GV and GIII, while showing negligible competence for GI, under laboratory conditions. Importantly, this represents the first evidence of GV transmission by Ae. albopictus, underscoring that this globally distributed and ecologically adaptable mosquito could act as a supplementary vector under certain ecological contexts. These findings highlight the need for continued monitoring of JEV genotype–vector interactions, while carefully distinguishing laboratory competence from field incrimination, to better assess their potential impact on future transmission dynamics in the context of ecological change and climate-driven expansion [18,39].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Campbell G.L. Hills S.L. Fischer M. Jacobson J.A. Hoke C.H. Hombach J.M. Marfin A.A. Solomon T. Tsai T. Tsu V.D. Estimated global incidence of Japanese encephalitis: A systematic review Bull. World Health Organ.20118976677410.2471/BLT.10.08523322084515 PMC 3209971 · doi ↗ · pubmed ↗
- 2Ricklin M.E. Garcìa-Nicolàs O. Brechbühl D. Python S. Zumkehr B. Posthaus H. Oevermann A. Summerfield A. Japanese encephalitis virus tropism in experimentally infected pigs Vet. Res.2016473410.1186/s 13567-016-0319-z 26911997 PMC 4765024 · doi ↗ · pubmed ↗
- 3Pham T.T. Meng S. Sun Y. Lv W. Bahl J. Inference of Japanese encephalitis virus ecological and evolutionary dynamics from passive and active virus surveillance Virus Evol.20162 vew 00910.1093/ve/vew 00927774302 PMC 4989885 · doi ↗ · pubmed ↗
- 4Furlong M. Adamu A.M. Hoskins A. Russell T.L. Gummow B. Golchin M. Hickson R.I. Horwood P.F. Japanese encephalitis enzootic and epidemic risks across Australia Viruses 20231545010.3390/v 1502045036851664 PMC 9962251 · doi ↗ · pubmed ↗
- 5Zhao S. Li Y. Fu S. Liu M. Li F. Liu C. Yu J. Rui L. Wang D. Wang H. Environmental factors and spatiotemporal distribution of Japanese encephalitis after vaccination campaign in Guizhou Province, China (2004–2016)BMC Infect. Dis.202121117210.1186/s 12879-021-06857-334809606 PMC 8607706 · doi ↗ · pubmed ↗
- 6Zhao X. Cao M. Feng H.H. Fan H. Chen F. Feng Z. Zhou X.H. Japanese encephalitis risk and contextual risk factors in southwest China: A Bayesian hierarchical spatial and spatiotemporal analysis Int. J. Environ. Res. Public Health 2014114201421710.3390/ijerph 11040420124739769 PMC 4024990 · doi ↗ · pubmed ↗
- 7Solomon T. Dung N.M. Kneen R. Gainsborough M. Vaughn D.W. Khanh V.T. Japanese encephalitis J. Neurol. Neurosurg. Psychiatry 20006840541510.1136/jnnp.68.4.40510727474 PMC 1736874 · doi ↗ · pubmed ↗
- 8Erlanger T.E. Weiss S. Keiser J. Utzinger J. Wiedenmayer K. Past, present, and future of Japanese encephalitis Emerg. Infect. Dis.200915110.3201/eid 1501.08031119116041 PMC 2660690 · doi ↗ · pubmed ↗