Chilean Aloysia Essential Oils: A Medicinal Plant Resource for Postharvest Disease Control
Valentina Silva, Catalina Ferreira, Susana Flores, Evelyn Muñoz, Constanza Reyes, Carmen Trujillo, Esperanza Gálvez, Katy Díaz, Alejandro Madrid

TL;DR
This paper explores the use of Chilean Aloysia essential oils as a natural alternative to synthetic fungicides for controlling postharvest fungal diseases in crops.
Contribution
The study identifies Aloysia citriodora and its compound farnesol as effective natural antifungal agents against multiple phytopathogens.
Findings
A. citriodora essential oil and farnesol inhibited three postharvest rot fungi with low EC50 values.
GC/MS analysis revealed distinct chemical compositions between A. citriodora and A. polystachya essential oils.
Molecular docking suggests farnesol interacts with succinate dehydrogenase, indicating a possible mechanism of action.
Abstract
Postharvest fungal rot causes significant economic losses in the agroindustry. Current control methods involving the use of synthetic fungicides are becoming increasingly ineffective and pose environmental risks. This necessitates exploring sustainable alternatives, such as essential oils derived from medicinal plants, to achieve safer and effective disease control. This research examined the chemical composition and efficacy of essential oils from Aloysia citriodora, Aloysia polystachya and their compounds against the postharvest rot fungi Monilinia fructicola, Monilinia laxa, and Botrytis cinerea. The main compounds of essential oils were analyzed by GC/MS and revealed differences in their composition. A. citriodora is characterized by the presence of spathulenol and caryophyllene oxide. In contrast, A. polystachya is characterized by the predominance of carvone. The results show that…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
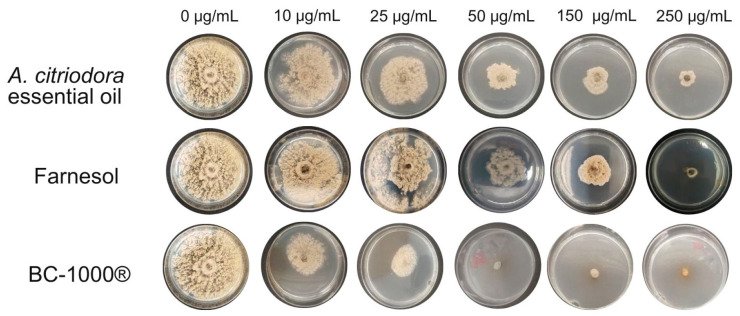 Figure 1
Figure 1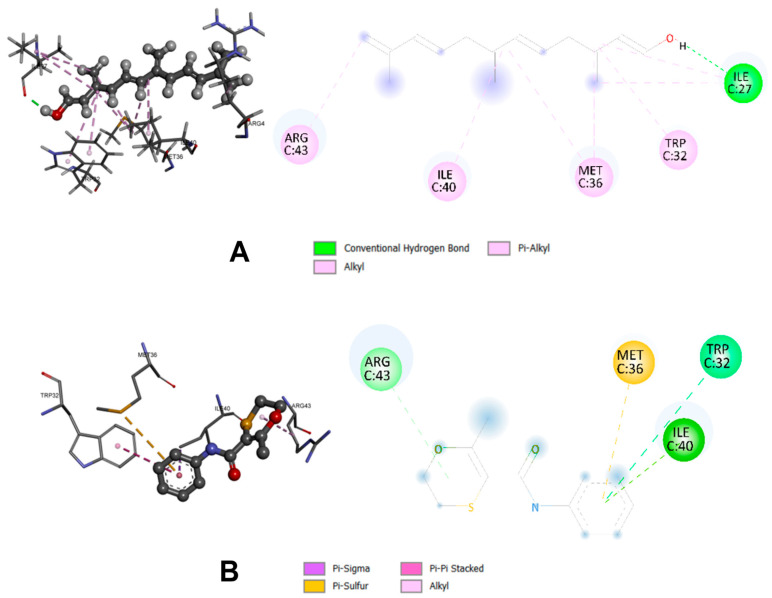 Figure 2
Figure 2- —Agencia Nacional de Investigación y Desarrollo (ANID)
- —Programa de Contratación de Ayudantes de Investigación de la Universidad de Playa Ancha en el Marco del Proyecto de Fortalecimiento
- —Programa de Apoyo para el pago de Procesamiento de Artículos InES Género 2025, Dirección General de Investigación, Universidad de Playa Ancha, Chile
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEssential Oils and Antimicrobial Activity · Insect Pest Control Strategies · Allelopathy and phytotoxic interactions
1. Introduction
Phytopathogenic fungi are a major contributor to plant diseases, causing significant economic losses and posing food safety risks due to production and accumulation of mycotoxins. Some disease-causing agents, such as Monilinia fructicola, Monilinia laxa and Botrytis cinerea are classified as necrotrophs, as they feed on dead host tissues. These pathogens are responsible for gray mold and brown rot, correspondingly, culminating in severe fungal ailments [1,2]. The pathogenicity mechanism exhibited by these fungal includes the exudation of toxins, enzymes responsible for the degradation of cellular walls, and other substances unique to the pathogenic agents [3], these fungal species exhibit a global distribution and are acknowledged for their capacity to induce wilting in floral structures, cause canker formation in ligneous tissues, and trigger decay in fruit bodies. Its life cycle depends on conidia, which are spread by wind, insects, or rain. The pathogen’s growth depends on favorable conditions like high humidity, frequent rainfall, and warm temperatures [4,5].
The interest in studying B. cinerea arises from the significant losses it causes in crop yield, resulting in economic damage ranging from USD 100 billion globally [6]. It also affects a wide variety of economically important crops, including strawberries, stone fruits, kiwifruits, tomatoes and grapes, especially in their postharvest [7]. This pathogen is hard to control due to its ability to infect many different plants and its diverse ways of causing disease. As a result, it leads to various symptoms, but the most recognizable one is the development of soft rot accompanied by gray masses of conidia, commonly known as gray mold [4]. Furthermore, approximately 10% of post-harvest productivity worldwide is lost due to brown rot disease caused by M. fructicola and M. laxa, it is an important constraint, difficult to control, which spreads rapidly in commercial crops in wet seasons especially in its main hosts Prunus sp. such as P. avium (Cherry), P. domestica (Plum), P. persica (Peach), P. nucipersica (Nectarine), P. salicina (Japanese Plum) among others, with emphasis during postharvest when ideal conditions for infection are achieved [8,9].
The European Union authorizes the use of fungicides such as quinone-outside inhibitors (QoI), succinate dehydrogenase inhibitors (SDHI) and methyl benzimidazole carbamates (MBCs) for the control of these phytopathogens [10]; the latter interact with the β-tubulin receptor, altering the formation of microtubules and consequently preventing mitosis in the fungus. The resistance mechanisms reported for these fungicides are related to binding site mutations in β-tubulin proteins for the case of studies conducted on Monilinia sp. [11]. On the other hand, monitoring of B. cinerea strains in different crops have shown a sustained increase in fungal resistance to these fungicides, including multi-resistance in crops that have been subjected to their use for years [10]. Among other treatments used for brown rot, copper-based products also are an alternative for its control [12]; however, copper is persistent in soil, not biodegradable and is toxic to non-target organisms and aquatic ecosystems, with long-lasting effects [13,14,15]. The economic impact of these phytopathogens in the agricultural industry, the resistance generated to their traditional control, their potential adverse effects on the environment, have led to the search for options for the control of phytopathogenic organisms that affect crops of agricultural interest.
The literature reports the use of essential oils derived from medicinal and aromatic plants as an alternative to control phytopathogens [16]. Essential oils, in general, contain monoterpenes and sesquiterpenes that show different mechanisms of action depending on their origin and composition; they may also have synergistic effects that reduce the risk of resistance development and are therefore considered as ingredients for the development of new biopesticides [17].
Aloysia citriodora and Aloysia polystachya commonly known as “Cedrón” and “Burrito”, respectively, are aromatic plant species known for their use in traditional medicinal and indigenous practice in South America, where they are native. A. citriodora is widely used in infusions for digestive disorders, in commercial beverages, insomnia management [18], and for its antioxidant activity [19], the study of its essential oil has demonstrated antibacterial and antifungal activity against phytopathogens such as Alternaria linariae, Sclerotinia sclerotiorum, and Fusarium sp. [20,21,22,23,24]. A. polystachya is one of the most frequently prescribed herbs in folk medicine, as an anxiolytic, in cardiovascular conditions [25,26,27,28] and for the treatment of the cultural-bound disease “empacho”, a digestive disorder [29,30]. Likewise, A. polystachya essential oil reports antibacterial activity [31], and regarding its antifungal activity, it shown to reduce the growth of Fusarium verticillioides (sin. Fusarium moniliforme), a species of fungus that infects corn [32]. The viability of Aloysia species as agricultural solutions is also reinforced by evidence of their safety profile. The traditional use of A. citriodora and A. polystachya in folk medicine, as well as toxicity studies conducted on Artemia salina, have shown that their essential oils are non-toxic [18]. At a regulatory level, the Food and Drug Administration (FDA) has classified Aloysia citriodora as a safe substance (GRAS), which allows for its incorporation into various food products [33]. However, to the best of our knowledge, no studies have determined the activities of the essential oils from the leaves of A. polystachya and A. citriodora on Monilinia sp. Therefore, the aim of this study was to determine the chemical composition and the antifungal efficacy of these essential oils and their components against the causal agents of postharvest rot: M. fructicola, M. laxa, and B. cinerea. Additionally, molecular docking of the most active compound (farnesol) was performed on the enzyme succinate dehydrogenase.
2. Results and Discussion
2.1. Chemical Composition of Essential Oils
Essential oil yield (weight/dry weight of plant) is 1.25% for A. citriodora leaves and 1.22% for A. polystachya leaves. GC-MS analysis of hydrodistillated A. citriodora essential oil allowed identifying 16 different components representing 99.93% of the relative proportion of these compounds in the oil. The global chromatographic analysis of A. citriodora oil showed a complex mixture consisting mainly of oxygenated mono- and sesquiterpenes and small amounts of sesquiterpene hydrocarbons. It was dominated by oxygenated sesquiterpenes (68.53%) and, to a lesser extent, by oxygenated monoterpenes (17.58%), while sesquiterpene hydrocarbons were present only in low percentages (13.82%). The major components detected in the oil were spathulenol (38.84%) and caryophyllene oxide (17.80%) followed by α-curcumene (8.57%), piperitone (7.22%), farnesol (5.13%), and citral (3.16%). In A. polystachya oil, GC/MS analysis identified 8 compounds, which represented 99.45% of the oil (Table 1). The major oil components were oxygenated monoterpenes, which usually occurs in the Aloysia genus; carvone was dominant among the major components (88.41%) followed by dihydrocarvone (4.57%) and R-limonene (3.90%).
Several studies have reported that the essential oil of A. citriodora typically contains high levels of monoterpenes [34,35,36]. In contrast, sesquiterpenes are usually present in moderate to low concentrations [18]. However, significant variations in the chemical composition of A. citriodora essential oils have been observed depending on geographic origin. Elechosa et al., studying the essential oils obtained from Northwestern Argentina, reported the existence of several chemotypes dominated by different components such as thujones (31.4–90.4%), citronellal (39.0–66.7%), carvone (48.2–70.9%), citral (51.0–54.3%), and linalool (78.2–85.1%) [37]. Similarly, in plants from the same region, Olmedo et al. found neral (27.3%), spathulenol (25.6%) and geranial (24.4%) as major constituents [38], getting closer to our results due to the higher proportion of spathulenol. In agreement with our findings, Oukerrou et al. reported a qualitatively similar sesquitepene-rich profile in A. citriodora cultivated in Morocco, with β-spathulenol (9.42–15.61%), α-curcumene (11.28–15.15%), and caryophyllene oxide (13.25–14.14%) as major components [20]. Along the same line, Tammal et al. identified spathulenol (13.07%) and caryophyllene oxide (11.78%) as predominant in oils extracted from plants collected in sub-humid regions of Tunisia [39]. In contrast, oils from arid and semi-arid zones in the same country were dominated by monoterpens particularly citral. Interestingly, Al-Maharik et al. reported that essential oils from A. citriodora grown in Jericho, an arid region in Palestine, exhibited a predominantly sesquiterpene profile (77.4%), with α-curcumene (26.94%), spathulenol (13.69%), geranial (10.79%), caryophyllene oxide (8.66%), and neral (7.59%) as the main constituents [40]. These findings suggest that the chemical composition of A. citriodora essential oils is highly variable and influenced by multiple factors such as regional climate, harvesting time, plant organ used, and the extraction method.
The chemical composition of the essential oil of A. polystachya was qualitatively consistent with that previously reported by our research group, in which the major constituents were R-carvone (91.03%), R-limonene (4.10%), and dihydrocarvone (1.07%) [41]. The predominance of R-carvone observed in this study aligns with reports from other A. polystachya oils obtained from Latin American regions, where R-carvone typically accounts for over 70% of the total composition [42,43].
2.2. Antifungal Activity of Essential Oils and Compounds
The essential oil of A. citriodora and some of its main constituents were active against B. cinerea, M. fructicola, and M. laxa, varying in levels of effectiveness. The A. citriodora oil showed the strongest inhibitory effect on the mycelial growth of M. fructicola (61.89 ± 1.16 µg/mL) and M. laxa (73.05 ± 3.1 µg/mL), being moderate for B. cinerea (183.26 ± 1.7 µg/mL), these results are shown in Table 2.
These findings are consistent with previous reports documenting the inhibitory activity of A. citriodora essential oil against phytopathogenic fungi such as Alternaria linariae, Sclerotinia sclerotiorum, and Fusarium sp. [22,24,39]. Similarly, other studies have demonstrated its moderate antifungal effect against B. cinerea, achieving 69% radial inhibition at a concentration of 250 ppm [44]. Fontana et al. also confirmed the oil’s efficacy against both B. cinerea and M. fructicola, reporting EC_50_ values of 0.63 mL/L and 0.21 mL/L, respectively [21]. This antifungal activity has often been attributed to its high content of monoterpenes, particularly citral and limonene, which are frequently described as the most abundant components in this species.
Among the analyzed components of the essential oil of A. citriodora, farnesol stood out for its potent antifungal activity. This compound registered the lowest EC_50_ values against M. laxa (45.32 ± 3.3 μg/mL) and M. fructicola (72.18 ± 0.07 μg/mL). It is important to note that there are no previous reports in the literature on the activity of farnesol specifically against M. fructicola. When comparing the results for M. laxa with those reported by Balsells-Llauradó et al. (2023), a superiority in potency is evident. Specifically, the maximum inhibition obtained against M. laxa was 84.2% at a concentration of 0.89 mg/mL [45]. This notable efficacy is achieved at a concentration up to 100 times lower than that used in the bibliographic study, establishing a considerably higher level of activity for pure farnesol against the tested strain. For the phytopathogen B. cinerea, an EC_50_ of 230.0 ± 3.1 μg/mL was determined. When contrasting this finding with the bibliography, it is evident that this concentration is significantly higher than the farnesol doses tested by Cotoras et al. (2012), whose maximum reported concentration was 166 μg/mL [46]. Given that our EC_50_ value exceeds the maximum concentration evaluated in that study by 1.38 times, this suggests a lower antifungal potency of farnesol under our assay conditions. In the present study, geraniol (EC_50_ = 163.4 ± 0.79 µg/mL) and nerolidol (EC_50_ = 173.5 ± 2.0 µg/mL) showed moderate activity against M. fructicola and M. laxa, respectively; however, their antifungal activity was notably lower against B. cinerea, with EC_50_ values exceeding 250 µg/mL for both. This low performance against B. cinerea partially contrasts with other reports in the literature, as one study calculated the EC_50_ of pure geraniol against this pathogen at 235 µg/mL, a value slightly better than the threshold observed here [47]. Additionally, it has been reported that geraniol can inhibit the mycelial growth of B. cinerea by 78% at a concentration of only 100 µg/mL and conidial germination by 96% at 250 µg/mL, suggesting that significant inhibition is possible at concentrations near this level [48]. On the other hand, the low activity of pure nerolidol against B. cinerea is well-supported, as despite being a main component (12.8%) of the essential oil of Baccharis dracunculifolia, one study showed that an extremely high concentration of 25 mg/mL was required to achieve 85.2% inhibition of mycelial growth. Therefore, the low performance observed in this work (EC_50_ > 250 µg/mL) is consistent with the characterization of nerolidol as a compound with moderate to low activity against this pathogen [49].
The inhibitory effects of most effective samples against M. fructicola are shown in Figure 1.
Nonetheless, several essential oils with sesquiterpene-rich profiles have also demonstrated strong antifungal properties. For example, the leaf essential oils of the medicinal plants Lannea egregia and Emilia sonchifolia, characterized by high concentrations of sesquiterpenes, were highly effective against Aspergillus niger [50]. Likewise, the fruit peel essential oil of Hymenaea stigonocarpa, in which spathulenol and caryophyllene oxide were the predominant compounds, showed antifungal activity against Botrytis cinerea, Sclerotinia sclerotiorum, and Colletotrichum truncatum [51]. Previous studies have shown that geraniol, nerolidol, and farnesol induce a cell death mechanism in filamentous phytopathogenic fungi by causing mitochondrial dysfunction and an increase in reactive oxygen species (ROS), leading to death by apoptosis [52,53,54]. In this study, the differences in the fungicidal activity of these compounds can be attributed to their respective chemical structures. Geraniol, being a monoterpene, is less lipophilic than farnesol, a sesquiterpene. This lower lipophilicity could limit its ability to penetrate fungal cell membranes. This hypothesis is supported by their logP values, with geraniol at 3.28, significantly lower than farnesol at 5.31. On the other hand, nerolidol and farnesol are both sesquiterpene alcohol isomers, which means they share the same molecular formula but differ in the spatial arrangement of their atoms. This characteristic is key, as farnesol is a primary allylic alcohol, making it more susceptible to oxidation than nerolidol, which is a tertiary alcohol. Additionally, farnesol longer chain and greater number of double bonds make it generally more prone to oxidation than geraniol [55]. The activity shown by farnesol is confirmed by a series of studies on phytopathogenic fungi such as Fusarium graminearum, Penicillium expansum, Rhizoctonia solani and Ustilaginoidea virens [46,56,57,58,59]. To the best of our knowledge, this is the first report of farnesol exhibiting inhibitory activity against species of the genus Monilinia. Despite belonging to the same genus, the essential oil from A. polystachya did not inhibit fungal growth, as none of its primary components—R-limonene, linalool, α-terpineol, dihydrocarvone, and R-carvone—showed significant activity. This can be attributed to its compositional profile, which differs from that of A. citriodora and lacks the specific bioactive metabolites effective against these filamentous fungi. This intraspecific variation is well-documented, as the efficacy of essential oils can differ based on their chemotype. For instance, studies on A. polystachya have shown that its carvone-rich chemotype is more effective against B. cinerea via fumigation than its α-thujone-rich chemotype [60]. Likewise, the main compound in A. polystachya oil, carvone, has been shown to inhibit the growth of filamentous fungi and even reduce the development of post-harvest lesions caused by B. cinerea in cherry tomatoes [61], also differing from our results. This discrepancy may be due to differences in the carvone isomer, the fungal strains, or the specific technique employed, as the aforementioned study applied carvone by fumigation it onto the PDA medium [61]. Regarding the existing evidence in Monilinia sp., our results are consistent with those obtained in other studies on dihydrocarvone, which followed the same methodology as our study, where this molecule has not been shown to inhibit mycelial growth and conidia germination of Monilinia sp. [62]. Consequently, A. polystachya does not appear to be a viable alternative for controlling these phytopathogens. Beyond efficacy, a critical consideration for any potential agricultural treatment is its impact on fruit quality. Previous research has shown that carvone, for instance, does not significantly affect the organoleptic quality of fruit upon application [61]. Similarly, other components found in essential oils, such as geraniol, nerolidol, and farnesol (present in the essential oil of A. citriodora), have been approved for foliar application in acaricide formulations like BioMite^®^, supporting their safety profile for agricultural use.
2.3. Molecular Docking
As a strategy to approach a possible mechanism of action of farnesol, a compound that has featured remarkably in antifungal assays, we have applied in silico assays on a target usually used by current chemical controls; the enzyme succinate dehydrogenase (SDH), also known as mitochondrial complex II, is an enzyme anchored to the inner mitochondrial membrane of aerobic organisms, which plays a key role in both the Krebs cycle and the electron transport chain. It is composed of four subunits: A and B, which are hydrophilic, and C and D, which are hydrophobic and embedded in the membrane [63].
Due to its metabolic relevance, SDH has been identified as a preferred target for the development of new fungicides [64,65,66]. Inhibitors of this enzyme constitute an important class of agricultural control agents, recognized for their high efficacy and, in some cases, broad-spectrum bactericidal activity [62]. The scientific literature describes a great structural diversity among potential SDH inhibitors; for example, Soto et al. [67] documented that certain hydrated geranylated phenol hydrates exert antifungal activity against B. cinerea through inhibition of this enzyme. In the present study, molecular docking of farnesol was performed using the crystal structure of SDH (PDB ID: 2FBW). The reference ligand was 2-methyl-N-phenyl-5,6-dihydro-1,4-oxathiine-3-carboxamide (CBE), present in the crystallized structure. The results (Table 3) show that farnesol exhibited a binding energy of −7.5 kcal/mol, slightly higher in affinity than CBE (−7.2 kcal/mol).
Both ligands shared interactions with key active-site residues, including Trp32, Met36, Ile40, and Arg43 (see Figure 2), suggesting that farnesol may bind to the same catalytic pocket and potentially interfere with enzymatic function in a comparable manner. Notably, farnesol also established an additional hydrogen bond with Ile27, absent in the CBE complex, which may contribute to increased complex stability and explain the modest improvement in calculated affinity. These findings are consistent with previous reports describing the antifungal activity of farnesol against Candida albicans, where it acts as a morphogenesis modulator, reduces biofilm formation, and enhances the effect of conventional antifungals [68,69]. In B. cinerea, it has been shown to inhibit mycelial growth and trigger an-apoptosis-like phenotype characterized by reactive oxygen species accumulation and structural damage [70].
Taken together, the observed affinity of farnesol for SDH, combined with a similar and partly superior interaction profile to that of a known inhibitor, supports the hypothesis that this sesquiterpene may act as a direct modulator of the enzyme. This highlights its potential as a structural scaffold for the design of new fungicides or as an adjuvant in the control of agriculturally important phytopathogens.
3. Materials and Methods
3.1. General Data
All reagents and compounds present of the essential oils were purchased from Sigma-Aldrich Co., (St. Louis, MO, USA), and AK Scientific Inc. (30023 Ahern Ave, Union City, CA 94587, USA).
3.2. Plant Material
Plant samples were collected, avoiding unhealthy specimens or signs of contamination, during spring of 2024, in Olmué, Valparaíso Region, Central Chile at an altitude of 200 mts (32°59′34″ S 71°09′34″ W) authentication was performed by Mr. Patricio Novoa, and voucher specimens (AC-0922 and AP-0922, A. citriodora and A. polystachya, respectively) was deposited for future reference in the Natural Products and Organic Synthesis Laboratory of Universidad de Playa Ancha, Valparaíso, Chile.
3.3. Essential Oil Extraction
To facilitate extraction, 500 g of each fresh plant was cut and ground in a food processor (Model: BLSTMGK15051, Oster, Racine, WI, USA). Each sample was then separately subjected to hydrodistillation with 2 L of water using a Clevenger-type apparatus [71]. The temperature was maintained at 100 °C until the water boiled, at which point it was reduced and controlled at 70 °C for 4 h. Hydrolate was subjected to liquid–liquid partition in a separator funnel and the obtained essentials oils were dried over anhydrous sodium sulfate and stored at 2–8 °C in amber glass vials. The yield of fresh essential oil was determined as the quotient of the weight of oil collected and the dry weight of plant material extracted. The yield was 1.25% for A. citriodora and 1.22% for A. polystachya.
3.4. Gas Chromatography Analyses
The sample (1 μL) was analyzed by Gas Chromatography–Mass Spectrometry (GC/MS). Analysis was carried out using a Hewlett-Packard GC/MS 6890 coupled to a Hewlett-Packard 5973 mass-selective detector (electron ionization, 70 eV, Palo Alto, CA, USA). Helium was used as carrier gas at a rate of 1.3 mL/min, and the capillary column used was a HP-5ms. The temperature program was 40 °C (5 min) to 280 °C (8 min) at a rate of 5 °C/min. Compounds in the chromatograms were identified by comparison of their mass spectra with those in the NIST 2021 library database, and by comparison of their retention index with those reported in the literature, for the same type of column or those of commercial standards, when available.
3.5. Antifungal Activity
Fungal Growth Conditions
Species used in this investigation were kindly provided by the collection of the Mycology Unit of the Servicio Agrícola y Ganadero (SAG) of Chile. The cultures were grown on 90 mm diameter Petri dishes on potato dextrose agar (PDA; DIFCO™).
3.6. Antifungal Assay In Vitro
Determination of mycelial growth inhibition of the samples was performed by measuring the radius of growth. The PDA media contained compounds or essential oils at concentrations between 10 and 250 µg/mL, previously dissolved in ethanol and water. The percentage of inhibition was determined according to methods described previously [72]. PDA 1% ethanol medium was used as a negative control, while a commercial fungicide, BC-1000^®^ (grapefruit extract of Citrus x paridisi, 50% w/v, CHEMIE, Providencia, Chile), was used as positive control and measured under the same conditions as the tested samples. For each treatment, a 4 mm disc of mycelium was inoculated in the center of a plate and incubated for 3 (for B. cinerea) o 5–7 days (Monilinia spp.), at 23 °C [62,73]. The results were expressed as the average effective concentration (EC_50_), that is, the concentration at which mycelial growth was reduced by 50%. This value was determined using Origin ProV. 8 software (OriginLab Corporation, Northampton, MA, USA). All treatments were performed in triplicate.
3.7. Molecular Docking
The three-dimensional models of the ligands were constructed using UCSF Chimera 1.18 software. Polar hydrogens were added to each ligand, Gasteiger charges were assigned, and energy minimization was performed using the General AMBER Force Field (GAFF). Graphical representations were created with the free version of Discovery Studio Visualizer. The crystal structure of succinate dehydrogenase (SDH, PDB ID: 2FBW, resolution 2.06 Å) was obtained from the Protein Data Bank http://www.rcsb.org/pdb (accessed on 11 August 2025). Molecular docking of SDH with the selected ligands was carried out using AutoDock4 software, applying the Lamarckian genetic algorithm. During docking, the protein was treated as rigid while ligands were fully flexible. Search parameters included 50 runs per ligand and a maximum of 25 million energy evaluations. A clustering threshold of RMSD < 0.5 Å was set. The cocristallized ligand in the enzyme, 2-methyl-N-phenyl-5,6-dihydro-1,4-oxathiine-3-carboxamide (CBE), was used as a reference for docking. The optimal protein–ligand complex was selected based on the most populated cluster with the lowest binding free energy (ΔG). To validate the accuracy of the protocol, the cocristallized ligand was redocked under the same conditions, yielding an RMSD of 1.29 Å. All experiments were conducted at physiological pH.
4. Conclusions
This study determined the composition and evaluated the in vitro antifungal activity of the essential oils from A. citriodora and A. polystachya. The results revealed significant differences in both the types of molecules present and their respective efficacy. The oil from A. citriodora, rich in oxygenated sesquiterpenes, showed greater activity against M. fructicola and M. laxa compared to the A. polystachya oil, which is a mixture primarily composed of oxygenated monoterpenes. This difference suggests a possible synergistic effect among the constituents of the A. citriodora oil. Of the compounds evaluated from both species, farnesol, found exclusively in A. citriodora, demonstrated strong antifungal activity against the same pathogens, unlike the bioactive metabolites present in A. polystachya. Based on these findings, the essential oil of A. citriodora and farnesol, in particular, show great potential as antifungal agents. However, further studies are needed to develop stable formulations that overcome the inherent instability of essential oils in practical applications.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Martini C. Mari M. Monilinia fructicola, Monilinia laxa (Monilinia Rot, Brown Rot)Postharvest Decay Elsevier Amsterdam, The Netherlands 201423326510.1016/B 978-0-12-411552-1.00007-7 · doi ↗
- 2Rungjindamai N. Jeffries P. Xu X.-M. Epidemiology and Management of Brown Rot on Stone Fruit Caused by Monilinia laxa Eur. J. Plant Pathol.201414011710.1007/s 10658-014-0452-3 · doi ↗
- 3Kou J. Wei Y. He X. Xu J. Xu F. Shao X. Infection of Post-Harvest Peaches by Monilinia fructicola Accelerates Sucrose Decomposition and Stimulates the Embden–Meyerhof–Parnas Pathway Hortic. Res.201854610.1038/s 41438-018-0046-x 30181886 PMC 6119188 · doi ↗ · pubmed ↗
- 4Rahman F.U. Zhu Q. Wu Z. Li X. Chen W. Xiong T. Zhu X. Current Insights into the Biocontrol and Biotechnological Approaches for Postharvest Disease Management of Botrytis cinerea Postharvest Biol. Technol.202421611305510.1016/j.postharvbio.2024.113055 · doi ↗
- 5De Miccolis Angelini R.M. Landi L. Raguseo C. Pollastro S. Faretra F. Romanazzi G. Tracking of Diversity and Evolution in the Brown Rot Fungi Monilinia fructicola, Monilinia fructigena, and Monilinia laxa Front. Microbiol.20221385485210.3389/fmicb.2022.85485235356516 PMC 8959702 · doi ↗ · pubmed ↗
- 6Bi K. Liang Y. Mengiste T. Sharon A. Killing Softly: A Roadmap of Botrytis cinerea Pathogenicity Trends Plant Sci.20232821122210.1016/j.tplants.2022.08.02436184487 · doi ↗ · pubmed ↗
- 7Elad Y. Pertot I. Cotes Prado A.M. Stewart A. Fillinger S. Elad Y. Plant Hosts of Botrytis spp Botrytis—The Fungus, the Pathogen and Its Management in Agricultural Systems Springer International Publishing Champaign, IL, USA 201610.1007/978-3-319-23371-0_20 · doi ↗
- 8Mustafa M.H. Bassi D. Corre M.-N. Lino L.O. Signoret V. Quilot-Turion B. Cirilli M. Phenotyping Brown Rot Susceptibility in Stone Fruit: A Literature Review with Emphasis on Peach Horticulturae 2021711510.3390/horticulturae 7050115 · doi ↗