Isolation and Microbiological and Molecular Identification of Brucella abortus in Cattle and Pigs, Slaughtered in Cattle Sheds Located in Northern Sierra of Ecuador
Maritza Celi-Erazo, Elizabeth Minda-Aluisa, Lisbeth Olmedo-Pinchao, Lenin Ron-Garrido, Tania Ortega-Sierra, Julián López-Balladares, Marlon Carlosama-Yépez, Santiago Gonzalón-Alcarraz, Jacobus H. de Waard, Claude Saegerman, Jorge Ron-Román, Washington Benítez-Ortiz

TL;DR
This study identifies Brucella abortus in cattle and pigs in Ecuador's northern highlands using multiple diagnostic methods.
Contribution
The study combines serology, culture, and molecular diagnostics to characterize Brucella abortus strains in livestock.
Findings
Brucella abortus biovar 4 was the most common strain isolated from cattle.
Seropositive pigs did not yield Brucella isolates, showing serology limitations in swine.
Active surveillance with traceability systems can guide interventions to reduce infection risks.
Abstract
Brucellosis remains an underreported zoonotic disease in Ecuador. Its control program in cattle integrates diagnostic testing, vaccination, and eradication incentives, although participation is largely voluntary. Since 2025, vaccination has become compulsory nationwide. Human surveillance remains largely passive, and strain-level data are very limited. This study applied an integrated approach, combining serology (Rose Bengal and SAT-EDTA), microbiological culture, and molecular diagnostics, to assess the presence and diversity of Brucella spp. in cattle and pigs from six slaughterhouses in the northern Andean highlands. A total of 2054 cattle and 1050 pigs from Carchi, Imbabura, and Pichincha were sampled. Among cattle, 133 (6.5%; 95% CI: 5.5–7.6) were seropositive, and viable B. abortus strains were isolated from 17 (12.8%). Genus identification was confirmed by IS711-PCR, while…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
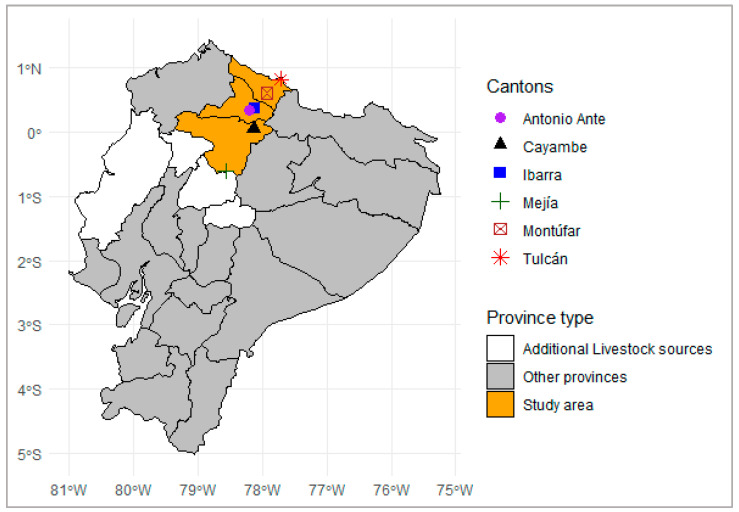 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3- —Research Directorate of the Central University of Ecuador
- —University of Liège
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBrucella: diagnosis, epidemiology, treatment · Monoclonal and Polyclonal Antibodies Research · Identification and Quantification in Food
1. Introduction
Brucellosis is a zoonotic disease caused by bacteria of the genus Brucella, which are Gram-negative, facultative intracellular coccobacilli infecting a wide range of mammals, including humans and livestock [1]. The disease remains a significant global zoonotic and public health and economic concern due to its impact on reproductive efficiency in animals and the risk of transmission to humans through direct contact or consumption of unpasteurized animal products. In Latin America, brucellosis persists despite control and vaccination programs; this endemicity is driven by factors such as complex smallholder livestock systems, unregulated animal movements, insufficient indemnities and surveillance, and under-resourced veterinary infrastructure [2].
In cattle, B. abortus is the species most frequently implicated, whereas B. suis predominates in pigs [3]. In cattle, acute infection is often associated with reproductive events, driven in part by the high concentrations of erythritol in placental and reproductive tissues, which strongly favor Brucella growth. This stage typically leads to abortion (often in the last trimester), retained placenta, orchitis, and reduced fertility. Chronic infection, by contrast, is characterized by persistent colonization of lymph nodes, mammary glands, and reproductive tissues, often without overt clinical signs, but with continued bacterial shedding and transmission risk. In pigs, acute brucellosis similarly manifests with abortion, infertility, orchitis, and lameness, while chronic cases may result in abscess formation in lymph nodes or reproductive organs, contributing to long-term persistence in herds. These clinical patterns complicate diagnosis and underscore the importance of laboratory confirmation for both surveillance and control. In this context, biovar typing is critical for distinguishing vaccine strains from circulating field isolates, tracing outbreaks, and assessing cross-species transmission. In Argentina, for instance, B. abortus biovars 1 and 4 are prevalent in cattle, and atypical B. suis strains—often resistant to dyes—have been associated with pigs [4].
In Ecuador, the epidemiological picture is complex and understudied. Although serological studies have indicated a national herd-level seroprevalence of 8% and an individual-level prevalence of 2.2% in dairy cattle [5], and 17% in dual-purpose systems [6]. Brucellosis control is addressed through the National Bovine Brucellosis Control Program, led by Agrocalidad, which integrates epidemiological surveillance, diagnostic testing, vaccination, and eradication incentives. Participation is still largely voluntary, and any herd can obtain “officially brucellosis-free” status through routine testing and culling of reactors; however, dairy herds receive stronger economic incentives (approximately USD 0.01 per liter of milk produced) once free status is achieved. This targeted benefit, absent in beef cattle operations, may partly explain the higher prevalence reported in certain coastal provinces where beef production predominates. Vaccination is a core component of the national bovine brucellosis control program in Ecuador; however, during the period of this study its application was voluntary and often limited to herds enrolled in official eradication schemes. Two vaccines were available: Brucella abortus strain S19 and strain RB51 in cattle. Coverage remains low, with estimates suggesting that fewer than 25% of animals are vaccinated nationwide [5]. Swine and in other livestock species vaccination is not practiced. However, as of 2025, vaccination with the B. abortus RB51 strain has become compulsory nationwide, specifically targeting bovine and buffalo females from 3 months of age onwards, with a two-dose scheme: the first dose administered between 3 and 9 months of age and a booster between 9 and 15 months (i.e., 6 months after the first). Moreover, strain and biovar characterization are not incorporated into the program, leaving important aspects of bovine brucellosis epidemiology and potential cross-species transmission pathways unknown. Oversight and enforcement limitations, including infrequent inspections, further constrain program effectiveness. Parra et al. (2021) reported molecular detection of B. abortus DNA in seropositive cattle from the Ecuadorian highlands, confirming infection of these animals by field strains using PCR assays targeting bcsp31 and omp2a [7]. Meanwhile, Ron-Román et al. (2019) documented regional seroprevalence and associated risk factors in Imbabura, Carchi, and Pichincha provinces—highlighting significant exposure in both cattle and farm workers and underscoring the need for intensified surveillance in these areas [8].
Regarding cattle movement, it clearly plays an important role in the spread of brucellosis. Network analyses of cattle movements (2017–2018) showed that over 90% of parishes are highly connected, with very low fragmentation, indicating frequent movement of animals between regions. Brucellosis, therefore, is likely to persist and circulate within local clusters of herds [9]. Although cattle transhumance (i.e., seasonal migration of cattle for grazing) is very limited in Ecuador, the smallholder systems common in the Andean region increase opportunities for transmission, as animals often share grazing areas and bulls are rented for natural mating. Information on the role of animal movements in other livestock species remains scarce, which limits a broader understanding of transmission dynamics.
Another relevant study in Santo Domingo de los Tsáchilas province identified B. abortus biovar 1 as the predominant field strain in cattle, combining classical biotyping with molecular assays, including IS711-PCR for genus confirmation, AMOS-PCR for species and biovar differentiation, and HOOF-Prints MLVA for high-resolution strain typing [10]. However, comprehensive biotyping studies involving other species, such as pigs (the major mammal consumed in Ecuador) and other livestock species, are lacking. Swine production systems in this Andean region range from smallholder backyard operations to semi-intensive farms, and many pigs have free-ranging access to other livestock species such as cattle, or to areas where contact with wildlife (e.g., feral pigs) can occur. This interface between domestic swine and other animal populations represents a potential pathway for cross-species transmission that remains poorly investigated in Ecuador.
Experiences from neighboring countries such as Costa Rica show the added value of incorporating strain-level diagnostics into national strategies. Hernández-Mora et al. (2017) characterized Brucella isolates from multiple species and identified B. abortus and B. suis biovars circulating in cattle and pigs [11]. Their work demonstrates the utility of combining molecular and classical techniques to strengthen public health risk assessments and to detect gaps in surveillance systems.
Molecular tools targeting conserved genes (e.g., bcsp31, omp2, IS711), coupled with classical bacteriology and biotyping, remain indispensable for identifying circulating strains and differentiating field from vaccine strains [12,13,14]. These methods are particularly important in areas where brucellosis remains endemic, and transmission dynamics are not well understood.
Therefore, the aim of this study was to isolate and characterize Brucella spp. strains from cattle and pigs sampled at slaughterhouses in the northern Andean provinces of Ecuador (Pichincha, Imbabura, and Carchi). Since isolation success is strongly associated with seropositivity, we focused on seropositive animals to recover bacterial isolates and generate strain- and biovar-specific data, thereby contributing to the understanding of local transmission pathways and supporting more targeted and effective brucellosis control efforts in Ecuador.
2. Materials and Methods
2.1. Description and Location of the Study
Between 2013 and 2017, several field visits were conducted to municipal slaughterhouses in the northern Ecuadorian provinces of Pichincha (n = 2; cantons: Mejía and Cayambe), Imbabura (n = 2; cantons: Ibarra and Antonio Ante), and Carchi (n = 2; cantons: Tulcán and Montúfar). Figure 1 shows the study area and the geographical distribution of the sampled slaughterhouses. The distribution of cattle sampled by province was as follows: Pichincha (32.13%, n = 660), Carchi (25.12%, n = 516), and Imbabura (22.54%, n = 463). Additional samples were obtained from cattle originating in other Andean provinces of Ecuador, including Cotopaxi (1.4%, n = 29), Tungurahua (1.3%, n = 27), and Cañar (0.1%, n = 2), as well as from the coastal provinces of Manabí (12.61%, n = 259) and Santo Domingo de los Tsáchilas (4.7%, n = 96). In swine, samples originated primarily from Carchi (53.90%; 566/1050), followed by Imbabura (34.76%; 365/1050) and Pichincha (11.33%; 119/1050), all of which are located within the defined study area.
During slaughter, blood samples were collected from the jugular vein using 100 mL sterile bottles. Additionally, tissue samples—including liver, spleen, and retromammary lymph nodes—were aseptically collected from the same animals. All samples were placed in sterile containers and transported under refrigeration (4 °C) to the laboratory for further analysis. A sampling protocol was applied whereby one animal was sampled for every five slaughtered. A total of 2054 cattle (including Bos taurus and Bos indicus) and 1050 pigs were sampled. Only the Mejia slaughterhouse did not process pigs during the sampling period.
For each sampled animal, a record form was completed to collect zootechnical and traceability data, including age, sex, breed, and geographic origin. This information was used to support the epidemiological analysis and spatial distribution of Brucella infection in the region.
2.2. Serological Tests for the Diagnosis of Brucellosis
Serological screening was performed using the Rose Bengal Test (RBT) and the Slow Tube Agglutination Test in the presence of EDTA (SAT-EDTA). To optimize laboratory resources and increase the likelihood of isolating Brucella, only animals that tested positive in at least one of the two serological tests were selected for bacteriological culture and strain isolation. Although tissue and lymph node samples were collected from all animals, isolation was conducted exclusively on seropositive animals. We acknowledge that this design does not allow detection of Brucella in seronegative animals, which could theoretically harbor the pathogen, but seropositive animals increase the likelihood of recovering Brucella from the tissues.
For the RBT, the Pourquier^®^ Rose Bengal antigen (IDEXX, Westbrook, ME, USA), derived from Brucella abortus Weybridge 99 strain, was used, following the World Organisation for Animal Health (WOAH, formerly OIE) standard protocol. Similarly, the SAT-EDTA test utilized antigen prepared from the same B. abortus Weybridge 99 strain, in line with WOAH/OIE recommendations. Sera were classified as positive based on visible agglutination, indicating the presence of Brucella-specific antibodies. Agglutination intensity was graded as follows: negative (−), low (+), moderate (++), high (+++), and very strong (++++) responses.
For the SAT-EDTA, the antigen was also obtained from IDEXX (Westbrook, ME, USA), and the procedure followed the protocol established by the Brucellosis Reference Laboratory of Belgium. A cut-off value of 30 International Agglutination Units (IAU)—equivalent to 25% agglutination at a 1:25 dilution—was used for both cattle and pig sera.
Samples testing positive by either or both tests were classified as seropositive under a parallel testing approach. The combined use of RBT and SAT-EDTA enables the detection of antibodies across both acute and chronic phases of infection, improving the diagnostic sensitivity of the screening protocol.
2.3. Isolation and Biotyping of the Brucella Species
Organ and lymph node samples from seropositive animals were individually macerated using a Stomacher^®^ homogenizer. Each macerate was cultured on Farrell’s selective medium, consisting of Columbia Blood Agar Base (CM0331, Oxoid, Basingstoke, United Kingdom) supplemented with 5% heat-inactivated horse serum (Ref. 16050–130, Gibco, New Zealand) and modified Brucella selective supplement (SR0209E, Oxoid, Basingstoke, United Kingdom). Inoculated plates were incubated at 37 °C with 5% CO_2_ for up to 4 weeks.
Colonies suspected to be Brucella spp. were subjected to a panel of microbiological and biochemical tests for confirmation and biotyping. Preliminary identification included colony morphology, Gram staining, and catalase activity. Species and biovar identification were carried out based on oxidase activity, hydrogen sulfide (H_2_S) production, urease activity, carbon dioxide (CO_2_) requirement, and agglutination with monospecific anti-A (Ref. R30164801 ZM01, Remel, Dartford, United Kingdom) and anti-M (Ref. R30164901 ZM02, Remel, Dartford, United Kingdom) sera specific for Brucella, which were used to confirm the antigenic expression of epitopes A and M.
Additionally, growth inhibition on media containing selective dyes (basic fuchsin, thionine, and safranin) was used for biovar differentiation, following established protocols for Brucella species classification [15]. Confirmed Brucella isolates were preserved at –80 °C in cryotubes containing glycerol-based cryopreservative.
For subsequent molecular analysis, 2–3 colonies from each isolate were suspended in 1.5 mL microtubes containing sterile distilled water. The suspensions were heat-inactivated at 65 °C, and their sterility was verified by subculturing onto Columbia Blood Agar to ensure the absence of viable organisms before DNA extraction and PCR testing.
2.4. Molecular Identification and Typing of Brucella spp.
For molecular identification, three PCR-based assays were performed to confirm genus, determine species, and differentiate vaccine strains from field isolates. Firstly, DNA extraction was performed from each Brucella-positive isolate. Bacterial suspensions were centrifuged at 2300× g for 10 min, and the resulting pellets were resuspended in TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH 8.0). A thermal lysis procedure consisting of two freeze–thaw cycles (65 °C/60 min) was applied. Supernatants were recovered by centrifugation and stored at −20 °C until further analysis.
Genus-level identification was achieved using IS711-PCR (also referred to as IS6501-PCR), targeting the IS711 insertion sequence, which is conserved among Brucella species and enables differentiation from other α-Proteobacteria. Amplification of a 261 bp product confirmed genus identification [16,17].
Species-level identification was conducted using the AMOS-PCR method [13], which allows multiplex detection of B. abortus biovars 1, 2, and 4 (498 bp); B. melitensis biovars 1–3 (731 bp); B. ovis (976 bp); and B. suis biovar 1 (285 bp). Reactions were performed with species-specific primers and known DNA controls.
To differentiate vaccine strains from field isolates, a PCR targeting the ery gene was performed. A 178-bp fragment, present in field strains but absent in the B. abortus S19 vaccine strain, was used as a marker. In addition, the absence of a 364-bp fragment specific to the RB51 vaccine strain was evaluated [18,19]. Amplifications were performed in a 20 μL reaction volume containing 1 U of Taq DNA polymerase, 1× buffer, 1.5 mM MgCl_2_, 0.2 mM of each dNTP, 0.2 μM of each primer, and approximately 10 ng of DNA template.
Primer sequences used for all PCR assays are provided in Table 1.
2.5. Data Analysis
All data were organized using Microsoft Excel^®^ and subsequently analyzed in the R environment (version 4.1.1). Seroprevalence estimates and their 95% confidence intervals were calculated under the assumption of simple random sampling for both cattle and pigs. Chi-square tests were used to assess associations between sex and seropositivity, as well as to compare seropositivity proportions among provinces. In addition, the Kappa coefficient was calculated to evaluate the level of agreement between the RB and SAT-EDTA serological tests. Statistical significance was set at p < 0.05.
3. Results
3.1. Bovine Testing Results
3.1.1. Serological Screening of Bovines
As summarized in Table 2, a total of 2054 cattle were serologically tested for brucellosis. Of these, 4.9% (101/2054) tested positive using the Rose Bengal Test (RBT), and 6.1% (126/2054) were positive using SAT-EDTA. When considering animals that tested positive in either or both tests (parallel testing), the estimated overall seroprevalence of brucellosis in cattle was 6.5% (135/2054; 95% CI: 5.5–7.6%). A high level of agreement was observed between the two serological tests, with a Kappa coefficient of 0.82 (95% CI: 0.78–0.86; p < 0.05), indicating strong concordance between RBT and SAT-EDTA in identifying seropositive animals.
Detailed data on the number of animals sampled, their provincial origin (only the origin of two animals was missing), and their serological and microbiological results are shown in Table 2. Notably, approximately 80% of the slaughtered cattle originated from the direct geographical area covered by this study. In the same way, 88.1% (119/135; 95% CI: 81.2–93.0%) of the seropositive animals originated from the study area itself, further supporting the local circulation and endemic presence of Brucella strains in the region.
3.1.2. Isolation and Biotyping of Brucella Isolates
A total of 17 isolates were obtained and identified as Brucella spp., representing an isolation rate of 12.6% (95% CI: 7.8–19.7%) from a total of 135 samples (Table 3). Among the different tissue samples analyzed, retromammary lymph nodes yielded the highest number of positive isolates, accounting for 76.47% (13/17) of all confirmed cases. Microbiological and biochemical characterization allowed for further typing of the isolates. B. abortus biovar 1 was identified in 4 out of 17 isolates (23.53%), while B. abortus biovar 4 was the predominant biovar, detected in 13 out of 17 isolates (76.47%).
Table 2 summarizes the geographical distribution of Brucella isolates and the origin of the sampled animals. Statistically significant differences were found in the proportion of isolates by sex (p < 0.05), with females representing 94.12% (16/17) of positive cases, compared to 5.88% (1/17) from males. Similarly, significant differences were observed in the distribution of isolates by slaughterhouse location (p < 0.05), with the highest number reported in Pichincha (58.82%, 10/17), followed by Carchi (29.41%, 5/17), and Imbabura (11.76%, 2/17). With respect to the province of origin of the animals, 47.06% of isolates (n = 8) came from Pichincha, 29.41% (n = 5) from Carchi, 17.65% (n = 3) from Imbabura, and 5.89% (n = 1) from Manabí.
Notably, isolate 14—originally classified as B. abortus biovar 4—exhibited atypical behavior in biochemical tests. Unlike the reference strain for this biovar, isolate 14 was not inhibited by Basic Fuchsin (20 µg) or Safranin (100 µg), suggesting a possible variant. Table 4 presents the results of microbiological and biochemical tests for identification and typing of Brucella spp. in cattle.
3.1.3. Molecular Identification
Table 5 shows the results of the molecular tests performed on the 17 Brucella spp. iso-lates from cattle. Complete concordance with microbiological typing was observed. Figure 2 shows the products of the IS711 PCR assay, which revealed a 261-bp band characteristic of the genus Brucella. Figure 3 presents the results of the AMOS-PCR assay, demonstrating a 498-bp fragment typical of B. abortus. In the same assay, PCR targeting the ery gene and RB51-specific fragments allowed the differentiation between field and vaccine strains of B. abortus. These results confirmed that the B. abortus biovar 1 isolates detected in this study were field strains, and that no live vaccine strains were circulating. Although the S19 vaccine strain was not included as a control, the expected fragment sizes for B. abortus vaccines are well documented in the literature.
3.2. Swine Testing
A total of 1050 swine blood samples were analyzed using both the Rose Bengal (RB) and Serum Agglutination Test with EDTA (SAT-EDTA) assays. Table 6 presents the distribution of sampled animals across slaughterhouses, the number of seropositive results per location, and the province of origin of the pigs.
Of the total samples, 11 animals tested positive by serology, distributed as follows: Carchi (n = 2), Imbabura (n = 4), and Pichincha (n = 5). The estimated overall seroprevalence was 1.0% (95% CI: 0.00–2.0%). The level of agreement between the RB and SAT-EDTA tests was low and not statistically significant, with a Kappa coefficient of 0.15 (95% CI: 0.09–0.20; p > 0.05).
None of the 11 lymph nodes and internal organ samples from seropositive swine yielded Brucella spp. isolates.
4. Discussion
This study aimed to identify and characterize Brucella spp. in cattle and pigs from slaughterhouses located in three provinces of the northern Ecuadorian highlands (Carchi, Imbabura, and Pichincha). It is the first study that combines serological screening, classical microbiology, biovar typing, and molecular diagnostics to assess Brucella infection dynamics across two livestock species in this region. Our findings provide important insights for the national brucellosis control program and offer baseline data for strain-level surveillance in Ecuador.
4.1. Brucellosis in Cattle
Using serology (RB and SAT-EDTA), we observed an overall seroprevalence of 6.5% in cattle. This value is consistent with previously reported estimates in Ecuador [5,6], reinforcing the endemic nature of bovine brucellosis in the region. The high agreement but not perfect between the two tests (κ = 0.82; 95% CI: 0.78–0.86) confirms their complementary diagnostic value: SAT-EDTA is more sensitive in detecting acute infections, while the RB test is better suited to chronic or post-vaccinal exposure [20].
Although the study was not designed to estimate national prevalence due to potential interference from vaccination (e.g., S19 or RB51), these findings should be interpreted in the context of Ecuador’s national brucellosis control program. As described, brucellosis control in Ecuador has historically relied on voluntary participation, with limited vaccination coverage and inconsistent enforcement. Although compulsory nationwide vaccination with B. abortus RB51 was only introduced in 2025, its effectiveness and coverage are still under evaluation. The detection of seropositive animals in slaughterhouses therefore reflects the epidemiological situation during the pre-compulsory vaccination period. Biotyping and molecular analysis revealed a predominance of B. abortus biovar 4 over biovar 1 in animals originating from the study area, and all isolates were confirmed as field strains, indicating that no live vaccine strains were circulating. Importantly, since most slaughtered animals analyzed in this work were sourced from the defined study area, these results are representative of the local epidemiological situation in this part of the Ecuadorian highlands rather than of the country as a whole. Overall, our study provides locally relevant evidence that complements official surveillance, highlights persistent gaps in herd-level control, and offers a baseline reference against which the future impact of compulsory RB51 vaccination can be assessed.
Brucella isolation was achieved in 12.6% (17/135) of seropositive cattle. This relatively low culture positivity is likely multifactorial. Our study specifically selected seropositive animals, independent of their pregnancy or parturition status. This approach could have increased the likelihood of isolation, but it might also not have had a substantial effect since many slaughtered animals were non-pregnant females or males, in which Brucella load in target tissues tends to be lower; further, Farrell’s medium (though widely used) has limited sensitivity for certain B. abortus strains contributing to reduced isolation rates as Brucella multiplies most in the reproductive tract. Additionally, acute serological responses perform better around parturition or abortion [21,22]. Notably, 76.5% of successful isolations were obtained from retromammary lymph nodes, supporting previous findings on tissue tropism related to erythritol metabolism and reproductive tract affinity [23,24].
The predominance of B. abortus biovar 4 (76.5%) over biovar 1 (23.5%) aligns with previous findings in the region [8,10]. Interestingly, biovar 1 was previously identified in cattle from the coastal province of Santo Domingo de los Tsáchilas [10]. In the current study, similar strains were detected, but now in livestock originating mainly from the same study area in highlands provinces of Pichincha, Carchi, and Imbabura, suggesting either a regional spread of these lineages or the persistence of common circulating strains across ecological zones. This underscores the need for continuous and geographically representative strain monitoring. It is a worth to mention that one isolate (Code 14) displayed atypical growth patterns in the presence of selective dyes, indicating a possible variant within biovar 4, as previously described in strains from Canada and the USA [25].
Brucellosis vaccination in Ecuador uses B. abortus strains S19 and RB51, with vaccination historically voluntary; but RB51 vaccine has become compulsory only in 2025. Our study identified circulating field strains. These findings suggest that the observed seroprevalence largely reflects natural infections rather than vaccine-induced antibodies, so that biovar-specific results highlight local transmission dynamics and underscore the value of molecular surveillance to distinguish field strains from vaccine strains.
4.2. Molecular Characterization
The molecular assays confirmed genus and biovar identification, with 100% agreement between phenotypic and genotypic typing. IS711-PCR confirmed all isolates as Brucella spp., while AMOS-PCR identified B. abortus biovars 1 and 4. To differentiate field strains from vaccine strains (S19 or RB51), we used PCR amplification of the ery gene (178 bp), which confirmed that all isolates were field strains, consistent with prior work [18,26]. While RB51 can also be identified using its rough phenotype and resistance to rifampicin, these simpler and cost-effective tests can complement molecular surveillance. In Ecuador, RB51 vaccination is now mandatory, but some farmers still use S19; therefore, integrating molecular detection with phenotypic monitoring is important for accurate surveillance and for distinguishing vaccine-related infections from field infections. These findings highlight the need for improved molecular surveillance and standardized vaccination practices as part of Ecuador’s national control strategy.
4.3. Brucellosis in Swine
This study provides the first evidence of Brucella seropositivity in pigs in Ecuador. Although the estimated seroprevalence was low (1.0%), the result is epidemiologically relevant, especially given the increasing recognition of swine as potential reservoirs of zoonotic Brucella spp. in Latin America [4,11]. The agreement between RB and SAT-EDTA tests was poor, probably due to the low number of positive samples; additionally, no Brucella spp. were isolated from tissue samples. These findings raise questions about the specificity of the serological tests in swine. Cross-reactivity of serological tests with Yersinia enterocolitica O:9, a common infection in pigs, may explain the seropositive results [27,28]. Given the shared O-polysaccharide antigen between Y. enterocolitica and Brucella spp., serological testing in pigs should be interpreted with caution. The absence of culture-positive samples in this study limits our ability to confirm true infections. Thus, we emphasize the need to evaluate alternative diagnostic approaches and improve test specificity for porcine brucellosis surveillance [29]. Additionally, the close contact between pigs and cattle, other livestock species in small-scale farming systems in Ecuador facilitates the potential transmission of Brucella spp. between species. Brucella suis biovar 1 has been reported in neighboring countries such as Colombia and Brazil [4], highlighting the potential for regional spread [29]. These findings underscore the importance of comprehensive surveillance and control measures in pigs and at the livestock-wildlife interfaces.
4.4. Value of Active Surveillance in Slaughterhouses
Our findings highlight the strategic value of slaughterhouse-based active surveil-lance. The ability to sample large numbers of animals from diverse geographical origins provides a unique opportunity to detect infected animals that may not show clinical signs. When paired with robust traceability data, such surveillance can help map transmission patterns and inform targeted interventions. Moreover, slaughterhouses represent high-risk points for occupational exposure. We observed infected animals being processed in facilities with inadequate biosafety implementation, reinforcing the need for improved sanitary protocols and routine health monitoring of slaughterhouse personnel [30].
5. Conclusions
This study confirms the circulation of Brucella abortus biovars 1 and 4—predominantly biovar 4—in cattle from the northern Andean highlands of Ecuador, with a 6.5% seroprevalence detected using RB and SAT-EDTA tests. Seventeen field strains were successfully isolated and molecularly characterized, none corresponding to vaccine strains. Although seropositivity was detected in 11 pigs, no isolates were recovered, marking the first report of seropositive swine in Ecuador, and highlighting the need for improved diagnostic specificity in this species. Given that approximately 80% of the cattle and nearly all the pigs originated from the study area, these results are representative of the local livestock population. The findings underscore the value of slaughterhouse-based active surveillance and traceability as practical tools for monitoring brucellosis, guiding control strategies, and mitigating zoonotic risk.
6. Study Limitations
This study was limited to three provinces in the northern Ecuadorian highlands; thus, the findings may not reflect the situation in other regions. While a relatively high seroprevalence was observed in cattle, the isolation rate of Brucella spp. was low—likely due to the study design targeting slaughterhouses and probably to use of a single culture medium. To increase the likelihood of successful isolation, we focused on seropositive animals, as isolation is more efficient from this group; however, this approach limited our ability to assess culture outcomes among seronegative individuals, which should be included in future studies, and in particular for prevalence estimations and surveillance activities. Also, this would have allowed us to detect possible subclinical carriers or individuals in early stages of infection, thus broadening the epidemiological picture. Future studies should incorporate multiple selective media (for instance CITA medium) and milk samples collected at different time points, and the inclusion of both B. suis biovar 1 (already reported in Colombia and Brazil) and biovar 2 (recognized in European wildlife and swine) in epidemiological surveillance efforts. Additionally, the failure to isolate Brucella from seropositive swine raises questions about potential false positives serological reactions due to cross-reactivity; further research is needed to refine diagnostic tools for pigs.
Likewise, the samples analyzed in this study were collected between 2013 and 2017. While the results provide valuable information on circulating Brucella strains during that period, the current prevalence and epidemiological patterns may differ due to changes in vaccination coverage, management practices, or control measures. This temporal limitation should be considered when extrapolating the findings to present-day conditions, and ongoing surveillance is recommended to monitor shifts in prevalence and strain circulation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hull N.C. Schumaker B.A. Comparisons of Brucellosis Between Human and Veterinary Medicine Infect. Ecol. Epidemiol.20188150084610.1080/20008686.2018.150084630083304 PMC 6063340 · doi ↗ · pubmed ↗
- 2Bonilla-Aldana D.K. Trejos-Mendoza A.E. Pérez-Vargas S. Rivera-Casas E. Muñoz-Lara F. Zambrano L.I. Arteaga-Livias K. Ulloque-Badaracco J.R. Alarcon-Braga E.A. Hernandez-Bustamante E.A. A Systematic Review and Meta-Analysis of Bovine Brucellosis Seroprevalence in Latin America and the Caribbean New Microbes New Infect.20235410116810.1016/j.nmni.2023.10116837692289 PMC 10492193 · doi ↗ · pubmed ↗
- 3Khurana S.K. Sehrawat A. Tiwari R. Prasad M. Gulati B. Shabbir M.Z. Chhabra R. Karthik K. Patel S.K. Pathak M. Bovine Brucellosis–A Comprehensive Review Vet. Q.20214161883335348910.1080/01652176.2020.1868616 PMC 7833053 · doi ↗ · pubmed ↗
- 4Lucero N.E. Ayala S.M. Escobar G.I. Jacob N.R. Brucella Isolated in Humans and Animals in Latin America from 1968 to 2006 Epidemiol. Infect.200813649650310.1017/S 095026880700879517559694 PMC 2870831 · doi ↗ · pubmed ↗
- 5Paucar V. Ron-Román J. Benítez-Ortiz W. Celi M. Berkvens D. Saegerman C. Ron-Garrido L. Bayesian Estimation of the Prevalence and Test Characteristics (Sensitivity and Specificity) of Two Serological Tests (RB and SAT-EDTA) for the Diagnosis of Bovine Brucellosis in Small and Medium Cattle Holders in Ecuador Microorganisms 20219181510.3390/microorganisms 909181534576711 PMC 8465933 · doi ↗ · pubmed ↗
- 6Carbonero A. Guzmán L.T. García-Bocanegra I. Borge C. Adaszek L. Arenas A. Saa L.R. Seroprevalence and Risk Factors Associated with Brucella Seropositivity in Dairy and Mixed Cattle Herds from Ecuador Trop. Anim. Health Prod.20185019720310.1007/s 11250-017-1421-628952067 · doi ↗ · pubmed ↗
- 7Parra K. Minda-Aluisa E. Navarro J.C. Información Filogenética de Los Genes Bcsp 31, Omp 2a y Omp 2b en el Diagnóstico y Epidemiología Molecular de Brucella spp.Universidad Internacional SEK Quito, Ecuador 2021
- 8Ron-Román J. Ron-Garrido L. Abatih E. Celi-Erazo M. Vizcaíno-Ordóñez L. Calva-Pacheco J. González-Andrade P. Berkvens D. Benítez-Ortíz W. Brandt J. Bayesian Evaluation of Three Serological Tests for Detecting Antibodies against Brucella Spp. Among Humans in the Northwestern Part of Ecuador Am. J. Trop. Med. Hyg.20191001312132010.4269/ajtmh.18-062231038099 PMC 6553909 · doi ↗ · pubmed ↗